
Evaluation of Carbonic Anhydrase, Acetylcholinesterase, Butyrylcholinesterase, and α-Glycosidase Inhibition Effects and Antioxidant Activity of Baicalin Hydrate


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Fe3+ Reducing Ability Assays
2.3. Cu2+ Reducing Ability Assays
2.4. DPPH• Radical Scavenging Activities
2.5. ABTS•+ Radical Scavenging Activities
2.6. DMPD•+ Radical Scavenging Activities
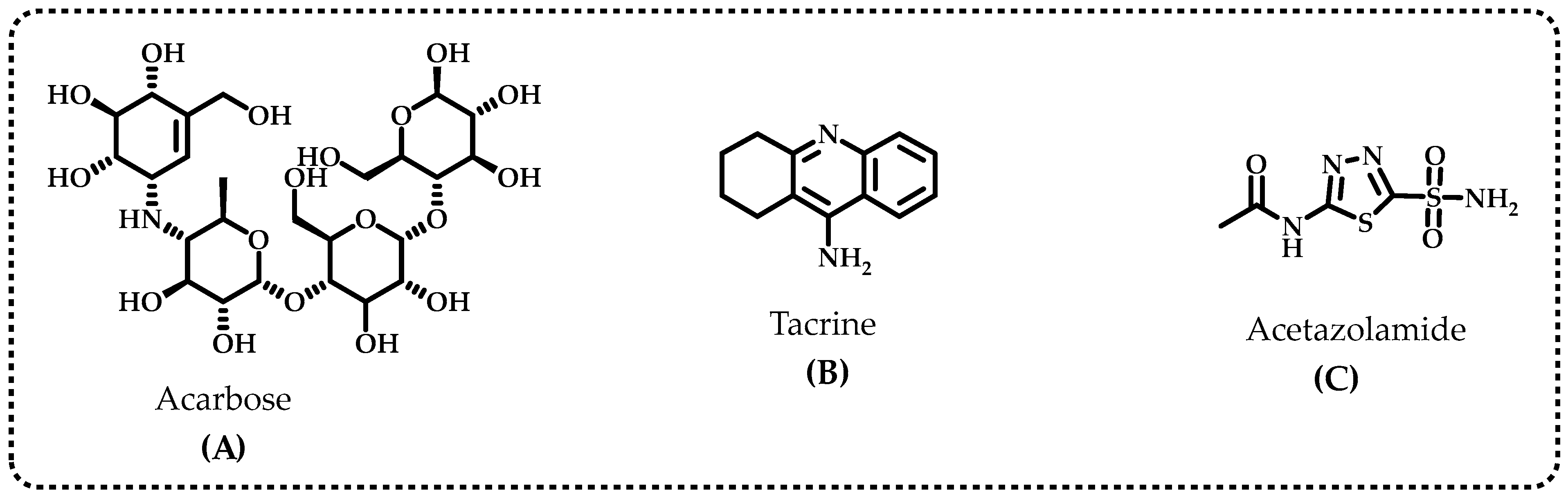
2.7. Acetylcholinesterase/Butyrylcholinesterase Enzymes Inhibition Assay
2.8. α-Glycosidase Enzyme Inhibition Assay
2.9. Human Carbonic Anhydrase II Isoenzyme Inhibition Assay
2.10. IC50 Values Determination
2.11. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, X.; Xie, L.; Long, J.; Liu, K.; Lu, J.; Liang, Y.; Cao, Y.; Dai, X.; Li, X. Therapeutic effect of baicalin on inflammatory bowel disease: A review. J. Ethnopharmacol. 2022, 283, 114749. [Google Scholar] [CrossRef]
- Manconi, M.; Caddeo, C.; Nacher, A.; Sales, O.D.; Peris, J.E.; Ferrer, E.E.; Fadda, A.M.; Manca, M.L. Eco-scalable baicalin loaded vesicles developed by combining phospholipid with ethanol, glycerol, and propylene glycol to enhance skin permeation and protection. Colloids Surf. B 2019, 184, 110504. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.; Nasr, M.; El-Sherbiny, I.M. Baicalin as an emerging magical nutraceutical molecule: Emphasis on pharmacological properties and advances in pharmaceutical delivery. J. Drug Deliv. Sci. Technol. 2022, 70, 103269. [Google Scholar] [CrossRef]
- Lai, W.; Jia, J.; Yan, B.; Jiang, Y.; Shi, Y.; Chen, L.; Mao, C.; Liu, X.; Tang, H.; Gao, M.; et al. Baicalin hydrate inhibits cancer progression in nasopharyngeal carcinoma by affecting genome instability and splicing. Oncotarget 2018, 9, 901–914. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-F.; Gao, F.; Li, X.-W.; Jia, R.-H.; Meng, X.-D.; Zhao, R.; Jing, Y.-Y.; Wang, Y.; Jiang, W. The anticonvulsant and neuroprotective effects of Baicalin on pilocarpine-induced epileptic model in rats. Neurochem. Res. 2012, 37, 1670–1680. [Google Scholar] [CrossRef] [PubMed]
- Martínez Medina, J.J.; Naso, L.G.; Pérez, A.L.; Rizzi, A.; Ferrer, E.G.; Williams, P.A.M. Antioxidant and anticancer effects and bioavailability studies of the flavonoid baicalin and its oxidovanadium(IV) complex. J. Inorg. Biochem. 2017, 166, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Fan, H.; Yin, P.; Yang, L.; Xue, Q.; Li, X.; Sun, L.; Liu, Y. Structure-activity relationship of eight high content flavonoids analyzed with a preliminary assign-score method and their contribution to antioxidant ability of flavonoids-rich extract from Scutellaria baicalensis shoots. Arab. J. Chem. 2018, 11, 159–170. [Google Scholar] [CrossRef]
- Mota, J.C.; Almeida, P.P.; Freitas, M.Q.; Stockler-Pinto, M.B.; Guimaraes, J.T. Far from being a simple question: The complexity between in vitro and in vivo responses from nutrients and bioactive compounds with antioxidant potential. Food Chem. 2023, 402, 134351. [Google Scholar] [CrossRef] [PubMed]
- Hatamnia, A.A.; Abbaspour, N.; Darvishzadeh, R. Antioxidant activity and phenolic profile of different parts of Bene (Pistacia atlantica subsp. kurdica) fruits. Food Chem. 2014, 145, 306–311. [Google Scholar] [CrossRef]
- Ozler, E.; Topal, F.; Topal, M.; Sarıkaya, S.B.O. LC-HRMS profiling and phenolic content, cholinesterase, and antioxidant activities of Terminalia citrina. Chem. Biodivers. 2023, 20, e202201250. [Google Scholar] [CrossRef]
- Topal, M. Secondary metabolites of ethanol extracts of Pinus sylvestris cones from eastern Anatolia and their antioxidant, cholinesterase and alpha-glucosidase activities. Rec. Nat. Prod. 2020, 14, 129–138. [Google Scholar] [CrossRef]
- Rana, A.; Samtiya, M.; Dhewa, T.; Mishra, V.; Aluko, R.E. Health benefits of polyphenols: A concise review. J. Food Biochem. 2022, 46, e14264. [Google Scholar] [CrossRef]
- Skrypnik, L.; Grigorev, N.; Michailov, D.; Antipina, M.; Danilova, M.; Pungin, A. Comparative study on radical scavenging activity and phenolic compounds content in water bark extracts of alder (Alnus glutinosa (L.) Gaertn.), oak (Quercus robur L.) and pine (Pinus sylvestris L.). Eur. J. Wood Wood Prod. 2019, 77, 879–890. [Google Scholar] [CrossRef]
- Sever, B.; Turkes, C.; Altıntop, M.D.; Demir, Y.; Ciftci, G.A.; Beydemir, S. Novel metabolic enzyme inhibitors designed through the molecular hybridization of thiazole and pyrazoline scaffolds. Arch. Pharm. 2021, 354, e2100294. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, H.; Gao, Z.; Xu, H. Effects of dietary baicalin supplementation on iron overload-induced mouse liver oxidative injury. Eur. J. Pharmacol. 2005, 509, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Froldi, G.; Djeujo, F.M.; Bulf, N.; Caparelli, E.; Ragazzi, E. Comparative evaluation of the antiglycation and anti-α-glucosidase activities of baicalein, baicalin (baicalein 7-O-glucuronide) and the antidiabetic drug metformin. Pharmaceutics 2022, 14, 2141. [Google Scholar] [CrossRef] [PubMed]
- Gulcin, I.; Taslimi, P.; Aygun, A.; Sadeghian, N.; Bastem, E.; Kufrevioglu, Ö.İ.; Turkan, F.; Sen, F. Antidiabetic and antiparasitic potentials: Inhibition effects of some natural antioxidant compounds on α-glycosidase, α-amylase and human glutathione S-transferase enzymes. Int. J. Biol. Macromol. 2018, 119, 741–746. [Google Scholar] [CrossRef]
- Sarikurkcu, C.; Zengin, G.; Oskay, M.; Uysal, S.; Ceylan, R.; Aktumsek, A. Composition, antioxidant, antimicrobial and enzyme inhibition activities of two Origanum vulgare subspecies (subsp. vulgare and subsp. hirtum) essential oils. Ind. Crops Prod. 2015, 70, 178–184. [Google Scholar] [CrossRef]
- Pedroso, T.F.d.M.; Bonamigo, T.R.; da Silva, J.; Vasconcelos, P.; Félix, J.M.; Cardoso, C.A.L.; Souza, R.I.C.; dos Santos, A.C.; Volobuff, C.R.F.; Formagio, A.S.N.; et al. Chemical constituents of Cochlospermum regium (Schrank) Pilg. root and its antioxidant, antidiabetic, antiglycation, and anticholinesterase effects in Wistar rats. Biomed. Pharmacother. 2019, 111, 1383–1392. [Google Scholar] [CrossRef]
- Durmaz, L.; Erturk, A.; Akyüz, M.; Polat, K.L.; Uc, E.M.; Bingol, Z.; Saglamtas, R.; Alwasel, S.; Gulcin, I. Screening of carbonic anhydrase, acetylcholinesterase, butyrylcholinesterase, and α-glycosidase enzyme inhibition effects and antioxidant activity of coumestrol. Molecules 2022, 27, 3091. [Google Scholar] [CrossRef]
- Zhao, J.; Lu, S.; Yu, H.; Duan, S.; Zhao, J. Baicalin and ginsenoside Rb1 promote the proliferation and differentiation of neural stem cells in Alzheimer’s disease model rats. Brain Res. 2018, 1678, 187–194. [Google Scholar] [CrossRef]
- Sun, M.; Su, M.; Sun, H. Spectroscopic investigation on the interaction characteristics and inhibitory activities between baicalin and acetylcholinesterase. Med. Chem. Res. 2018, 27, 1589–1598. [Google Scholar] [CrossRef]
- Aktas Anıl, D.; Ozturk Aydın, B.; Demir, Y.; Turkmenoglu, B. Design, synthesis, biological evaluation and molecular docking studies of novel 1H-1,2,3-triazole derivatives as potent inhibitors of carbonic anhydrase, acetylcholinesterase and aldose reductase. J. Mol. Struct. 2022, 1257, 132613. [Google Scholar] [CrossRef]
- Bayrak, C.; Taslimi, P.; Kahraman, H.S.; Gulcin, I.; Menzek, A. The first synthesis, carbonic anhydrase inhibition and anticholinergic activities of some bromophenol derivatives with S including natural products. Bioorg. Chem. 2019, 85, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Kalaycı, M.; Turkes, C.; Arslan, M.; Demir, Y.; Beydemir, S. Novel benzoic acid derivatives: Synthesis and biological evaluation as multitarget acetylcholinesterase and carbonic anhydrase inhibitors. Arch. Pharm. 2021, 354, e2000282. [Google Scholar] [CrossRef] [PubMed]
- Osmaniye, D.; Turkes, C.; Demir, Y.; Okay, Y.; Beydemir, S.; Kaplancıklı, Z.A. Design, synthesis, and biological activity of novel dithiocarbamate-methylsulfonyl hybrids as carbonic anhydrase inhibitors. Arch. Pharm. 2022, 355, e2200132. [Google Scholar] [CrossRef] [PubMed]
- Turkan, F.; Cetin, A.; Taslimi, P.; Karaman, M.; Gulcin, I. Synthesis, biological evaluation and molecular docking of novel pyrazole derivatives as potent carbonic anhydrase and acetylcholinesterase inhibitors. Bioorg. Chem. 2019, 86, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Durmaz, L.; Kiziltas, H.; Guven, L.; Karagecili, H.; Alwasel, S. Antioxidant, antidiabetic, anticholinergic, and antiglaucoma effects of magnofluorine. Molecules 2022, 27, 5902. [Google Scholar] [CrossRef] [PubMed]
- Sever, B.; Turkes, C.; Altintop, M.D.; Demir, Y.; Beydemir, S. Thiazolyl-pyrazoline derivatives: In vitro and in silico evaluation as potential acetylcholinesterase and carbonic anhydrase inhibitors. Int. J. Biol. Macromol. 2020, 163, 1970–1988. [Google Scholar] [CrossRef]
- Hou, Z.; Li, C.; Liu, Y.; Zhang, M.; Wang, Y.; Fan, Z.; Guo, C.; Lin, B.; Liu, Y. Design, synthesis and biological evaluation of carbohydrate-based sulphonamide derivatives as topical antiglaucoma agents through selective inhibition of carbonic anhydrase II. J. Enzym. Inhib. Med. Chem. 2020, 35, 383–390. [Google Scholar] [CrossRef]
- Gong, L.; Zhu, J. Baicalin alleviates oxidative stress damage in trabecular meshwork cells in vitro. Naunyn. Schmiedebergs. Arch. Pharmacol. 2018, 391, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040, A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, Q.; Shi, L.; Zhou, X.; Wu, W.; Wang, X.; Wang, L.; Wu, Z. Baicalin prevents fibrosis of human trabecular meshwork cells via inhibiting the MyD88/NF-κB pathway. Eur. J. Pharmacol. 2023, 938, 175425. [Google Scholar] [CrossRef] [PubMed]
- Turkes, C.; Arslan, M.; Demir, Y.; Cocaj, L.; Nixha, A.R.; Beydemir, S. Synthesis, biological evaluation and in silico studies of novel N-substituted phthalazine sulfonamide compounds as potent carbonic anhydrase and acetylcholinesterase inhibitors. Bioorg. Med. 2019, 89, 103004. [Google Scholar]
- Karagecili, H.; Yilmaz, M.A.; Erturk, A.; Kiziltas, H.; Guven, L.; Alwasel, S.H.; Gulcin, I. Comprehensive comprehensive metabolite profiling of berdav propolis using LC-MS/MS: Determination of antioxidant, anticholinergic, antiglaucoma, and antidiabetic effects. Molecules 2023, 28, 1739. [Google Scholar] [CrossRef] [PubMed]
- Oyaizu, M. Studies on product of browning reaction prepared from glucoseamine. Jpn. J. Nut. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Apak, R.; Calokerinos, A.; Gorinstein, S.; Segundo, M.A.; Hibbert, D.B.; Gulcin, I.; Demirci Cekic, S.; Guclu, K.; Ozyürek, M.; Esin Celik, S.; et al. Methods to evaluate the scavenging activity of antioxidants toward reactive oxygen and nitrogen species. Pure Appl. Chem. 2022, 94, 87–144. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 26, 1199–1200. [Google Scholar] [CrossRef]
- Ozcelik, B.; Lee, J.H.; Min, D.B. Effects of light, oxygen and pH on the 2,2-diphenyl-1-picrylhydrazyl (DPPH) method to evaluate antioxidants. J. Food Sci. 2003, 68, 487–490. [Google Scholar] [CrossRef]
- Kiziltas, H. Determination of LC-HRMS profiling, antioxidant activity, cytotoxic effect and enzyme inhibitory properties of Satureja avromanica using in vitro and in silico methods. Proces. Biochem. 2022, 116, 157–172. [Google Scholar] [CrossRef]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritiene, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Zhang, Y.; Cheng, Y.; Wang, Y. Rapid screening and identification of α-glucosidase inhibitors from mulberry leaves using enzyme-immobilized magnetic beads coupled with HPLC/MS and NMR. Biomed. Chromatogr. 2013, 27, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Istrefi, Q.; Türkeş, C.; Arslan, M.; Demir, Y.; Nixha, A.R.; Beydemir, Ş.; Küfrevioğlu, Ö.İ. Sulfonamides incorporating ketene N, S-acetal bioisosteres as potent carbonic anhydrase and acetylcholinesterase inhibitors. Arch. Pharm. 2017, 353, e1900383. [Google Scholar] [CrossRef] [PubMed]
- Verpoorte, J.A.; Mehta, S.; Edsall, J.T. Esterase activities of human carbonic anhydrases B and C. J. Biol. Chem. 1967, 242, 4221–4229. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 67, 248–254. [Google Scholar] [CrossRef]
- Karageçili, H.; İzol, E.; Kireçci, E.; Gülcin, İ. Antioxidant, antidiabetic, antiglaucoma, and anticholinergic effects of Tayfi grape (Vitis vinifera): A phytochemical screening by LC-MS/MS analysis. Open Chem. 2023, 21, 20230120. [Google Scholar] [CrossRef]
- Kuzu, B.; Tan, M.; Gulcin, I.; Menges, N. A novel class for carbonic anhydrases inhibitors and evaluation of their non-zinc binding. Arch. Pharm. 2021, 354, e2100188. [Google Scholar] [CrossRef]
- Nar, M.; Çetinkaya, Y.; Gülçin, I.; Menzek, A. (3,4-Dihydroxyphenyl)(2,3,4-trihydroxyphenyl)methanone and its derivatives as carbonic anhydrase isoenzymes inhibitors. J. Enzyme Inhib. Med. Chem. 2015, 28, 402–406. [Google Scholar] [CrossRef]
- Lineweaver, H.; Burk, D. The determination of enzyme dissociation constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Bursal, E.; Gulcin, I. Polyphenol contents and in vitro antioxidant activities of lyophilized aqueous extract of kiwifruit (Actinidia deliciosa). Food Res. Int. 2011, 44, 1482–1489. [Google Scholar] [CrossRef]
- Li, H.Y.; Yuan, Q.; Yang, Y.L.; Han, Q.H.; He, J.L.; Zhao, L.; Zhang, Q.; Liu, S.X.; Lin, D.-R.; Wu, D.T.; et al. Phenolic profiles, antioxidant capacities, and inhibitory effects on digestive enzymes of different kiwifruits. Molecules 2018, 23, 2957. [Google Scholar] [CrossRef] [PubMed]
- Cetin Cakmak, K.; Gulcin, I. Anticholinergic and antioxidant activities of usnic acid-An activity-structure insight. Toxicol. Rep. 2019, 6, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, I. Antioxidant activity of caffeic acid (3,4-dihydroxycinnamic acid). Toxicology 2006, 217, 213–220. [Google Scholar] [CrossRef]
- Capuzzo, A.; Occhipinti, A.; Maffei, M.E. Antioxidant and radical scavenging activities of chamazulene. Nat. Prod. Res. 2014, 28, 2321–2323. [Google Scholar] [CrossRef] [PubMed]
- Gulcin, I.; Huyut, Z.; Elmastas, M.; Aboul-Enein, H.Y. Radical scavenging and antioxidant activity of tannic acid. Arab. J. Chem. 2010, 3, 43–53. [Google Scholar] [CrossRef]
- Durmaz, L.; Kiziltas, H.; Karagecili, H.; Alwasel, S.; Gulcin, İ. Potential antioxidant, anticholinergic, antidiabetic and antiglaucoma activities and molecular docking of spiraeoside as a secondary metabolite of onion (Allium cepa). Saudi Pharm. J. 2023, 31, 101760. [Google Scholar] [CrossRef]
- Gocer, H.; Gulcin, I. Caffeic acid phenethyl ester (CAPE): Correlation of structure and antioxidant properties. Int. J. Food Sci. Nutr. 2011, 62, 821–825. [Google Scholar] [CrossRef]
- Wong, C.Y.; Leong, K.H.; He, X.; Zheng, F.; Sun, J.; Wang, Z.; Heh, C.H.; Kong, K.W. Phytochemicals of six selected herbal plants and their inhibitory activities towards free radicals and glycation. Food Biosci. 2022, 46, 101557. [Google Scholar] [CrossRef]
- Karimov, A.; Orujova, A.; Taslimi, P.; Sadeghian, N.; Mammadov, B.; Karaman, H.S.; Farzaliyev, V.; Sujayev, A.; Tas, R.; Alwasel, S.; et al. Novel functionally substituted esters based on sodium diethyldithiocarbamate derivatives: Synthesis, characterization, biological activity and molecular docking studies. Bioorg. Chem. 2020, 99, 103762. [Google Scholar] [CrossRef]
- Hashmi, S.; Khan, S.; Shafiq, Z.; Taslimi, P.; Ishaq, M.; Sadeghian, N.; Karaman, H.S.; Akhtar, N.; Islam, M.; Asari, A.; et al. Probing 4-(diethylamino)-salicylaldehyde-based thiosemicarbazones as multi-target directed ligands against cholinesterases, carbonic anhydrases and α-glycosidase enzymes. Bioorg. Chem. 2021, 107, 104554. [Google Scholar] [CrossRef] [PubMed]
- Kazancı, A.; Gok, Y.; Kaya, R.; Aktas, A.; Taslimi, P.; Gulcin, I. Synthesis, characterization and bioactivities of dative donor ligand N-heterocyclic carbene (NHC) precursors and their Ag(I)NHC coordination compounds. Polyhedron 2021, 193, 114866. [Google Scholar] [CrossRef]
- Karageçili, H.; Yilmaz, M.A.; Alwasel, S.H.; Gülcin, İ. Comprehensively revealing the profile of Pistacia vera L. cv. Siirt turpentine-Antioxidant, antidiabetic, anti-alzheimer, and antiglaucoma effects. Rec. Nat. Prod. 2023, 17, 918–937. [Google Scholar]
- Yeshi, K.; Yangdon, P.; Kashyap, S.; Wangchuk, P. Antioxidant activity and the polyphenolic and flavonoid contents of five high altitude medicinal plants used in Bhutanese Sowa rigpa medicine. J. Biol. Act. Prod. Nat. 2017, 7, 18–26. [Google Scholar]
- Topal, M.; Gocer, H.; Topal, F.; Kalin, P.; Polat Köse, P.; Gulcin, I.; Cetin Cakmak, K.C.; Kücük, M.; Durmaz, L.; Goren, A.C.; et al. Antioxidant, antiradical and anticholinergic properties of cynarin purified from the illyrian thistle (Onopordum illyricum L.). J. Enzyme Inhib. Med. Chem. 2016, 31, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Oktay, M.; Gülçin, I.; Küfrevioǧlu, Ö.I. Determination of in vitro antioxidant activity of fennel (Foeniculum vulgare) seed extracts. Lebensm. Wissen. Technol. 2003, 36, 263–271. [Google Scholar] [CrossRef]
- Cakmakcı, S.; Topdas, E.F.; Kalın, P.; Han, H.; Sekerci, P.; Polat Kose, L.; Gulcin, I. Antioxidant capacity and functionality of oleaster (Elaeagnus angustifolia L.) flour and crust in a new kind of fruity ice cream. Int. J. Food Sci. Technol. 2015, 50, 472–481. [Google Scholar] [CrossRef]
- Wen, Y.F.; Zhao, J.Q.; Bhadauria, M.; Nirala, S.K. Baicalin prevents cadmium induced hepatic cytotoxicity, oxidative stress and histomorphometric alterations. Exp. Toxicol. Pathol. 2013, 65, 189–196. [Google Scholar] [CrossRef]
- Gulcin, I. Antioxidant activity of eugenol-a structure and activity relationship study. J. Med. Food 2011, 14, 975–985. [Google Scholar] [CrossRef]
- Gulcin, I. Antioxidant properties of resveratrol: A structure-activity insight. Innov. Food Sci. Emerg. 2010, 11, 210–218. [Google Scholar] [CrossRef]
- Topal, F.; Topal, M.; Gocer, H.; Kalın, P.; Kocyigit, U.M.; Gulcin, I.; Alwasel, S.H. Antioxidant activity of taxifolin: An activity-structure relationship. J. Enzyme Inhib. Med. Chem. 2016, 31, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Taslimi, P.; Gulcin, I. Antioxidant and anticholinergic properties of olivetol. J. Food Biochem. 2018, 42, e12516. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, Z.Y.; Tang, R.C. Bioactive and UV protective silk materials containing baicalin-The multifunctional plant extract from Scutellaria baicalensis Georgi. Mater. Sci. Eng. C 2016, 67, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Gulcin, I.; Mshvildadze, V.; Gepdiremen, A.; Elias, R. Antioxidant activity of a triterpenoid glycoside isolated from the berries of Hedera colchica: 3-O-(β-D-glucopyranosyl)-hederagenin. Phytother. Res. 2016, 20, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, R.; Kumar, R.P.A. Baicalin provides protection against fluoxetine-induced hepatotoxicity by modulation of oxidative stress and inflammation. World J. Hepatol. 2022, 5182, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Bi, Y.; Zheng, X.; Lu, M.; Diao, Q.; Tu, Y. Effect of baicalin supplementation on the growth, health, antioxidant and anti-inflammatory capacity, and immune function of preweaned calves. Anim. Feed Sci. Technol. 2023, 298, 115598. [Google Scholar] [CrossRef]
- Gocer, H.; Akıncıoglu, A.; Oztaskın, N.; Goksu, S.; Gulcin, I. Synthesis, antioxidant and antiacetylcholinesterase activities of sulfonamide derivatives of dopamine related compounds. Arch. Pharm. 2013, 346, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Taslimi, P.; Koksal, E.; Goren, A.C.; Bursal, E.; Aras, A.; Kılıc, O.; Alwasel, S.; Gulcin, I. Anti-Alzheimer, antidiabetic and antioxidant potential of Satureja cuneifolia and analysis of its phenolic contents by LC-MS/MS. Arab. J. Chem. 2020, 13, 4528–4537. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef]
- Laaroussi, H.; Ferreira-Santos, P.; Genisheva, Z.; Bakour, M.; Ousaaid, D.; Teixeira, J.A.; Lyoussi, B. Unraveling the chemical composition, antioxidant, α-amylase and α-glucosidase inhibition of Moroccan propolis. Food Biosci. 2021, 42, 101160. [Google Scholar] [CrossRef]
- Cheng, F.; Lu, Y.; Zhong, X.; Song, W.; Wang, X.; Sun, X.; Qin, J.; Guo, S.; Wang, Q. Baicalin’s therapeutic time window of neuroprotection during transient focal cerebral ischemia and its antioxidative effects in vitro and in vivo. Evid. Based Complement Altern Med. 2013, 2013, 120261. [Google Scholar] [CrossRef] [PubMed]
- Peng-Fei, L.; Fu-Gen, H.; Bin-Bin, D.; Tian-Sheng, D.; Xiang-Lin, H.; Ming-Qin, Z. Purification and antioxidant activities of baicalin isolated from the root of huangqin (Scutellaria baicalensis gcorsi). J. Food Sci. Technol. 2013, 50, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Köksal, E.; Gülçin, I.; Beyza, S.; Sarikaya, Ö.; Bursal, E. In vitro antioxidant activity of silymarin. J. Enzym. Inhib. Med. Chem. 2009, 24, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, I. Antioxidant activity of l-adrenaline: A structure-activity insight. Chem. Biol. Interact. 2009, 179, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Ak, T.; Gülçin, I. Antioxidant and radical scavenging properties of curcumin. Chem. Biol. Interact. 2008, 174, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radical Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Roginsky, V.; Lissi, E.A. Review of methods to determine chainbreaking antioxidant activity in food. Food Chem. 2005, 92, 235–254. [Google Scholar] [CrossRef]
- Gulcin, I.; Sat, I.G.; Beydemir, S.; Küfrevioğlu, O.I. Evaluation of the in vitro antioxidant properties of extracts of broccoli (Brassica oleracea L.). Ital. J. Food Sci. 2004, 16, 17–30. [Google Scholar]
- Özenver, N.; Güvenalp, Z.; Kuruüzüm-Uz, A.; Demirezer, L.O. Inhibitory potential on key enzymes relevant to type II diabetes mellitus and antioxidant properties of the various extracts and phytochemical constituents from Rumex acetosella L. J. Food Biochem. 2020, 44, e13415. [Google Scholar] [CrossRef]
- Yu, M.; Han, S.; Wang, M.; Han, L.; Huang, Y.; Bo, P.; Fang, P.; Zhang, Z. Baicalin protects against insulin resistance and metabolic dysfunction through activation of GALR2/GLUT4 signaling. Phytomedicine 2022, 95, 153869. [Google Scholar] [CrossRef]
- Tanveer, A.; Akram, K.; Farooq, U.; Hayat, Z.; Shafi, A. Management of diabetic complications through fruit flavonoids as a natural remedy. Crit. Rev. Food Sci. Nutr. 2017, 57, 1411–1422. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Liu, J.G.; Li, H.; Yang, H.M. Pharmacological Effects of active components of chinese herbal medicine in the treatment of Alzheimer’s disease: A review. Am. J. Chin. Med. 2016, 44, 1525–1541. [Google Scholar] [CrossRef]
- Gulcin, I.; Scozzafava, A.; Supuran, C.T.; Akıncıoglu, H.; Koksal, Z.; Turkan, F.; Alwasel, S. The effect of caffeic acid phenethyl ester (CAPE) metabolic enzymes including acetylcholinesterase, butyrylcholinesterase, glutathione s-transferase, lactoperoxidase and carbonic anhydrase isoenzymes I, II, IX and XII. J. Enzyme Inhib. Med. Chem. 2016, 31, 1095–1101. [Google Scholar] [CrossRef]
- Aksu, K.; Ozgeriş, B.; Taslimi, P.; Naderi, A.; Gulcin, I.; Goksu, S. Antioxidant activity, acetylcholinesterase and carbonic anhydrase inhibitory properties of novel ureas derived from phenethylamines. Arch. Pharm. 2016, 349, 944–954. [Google Scholar] [CrossRef]
- Erdoğan, M.; Polat Kose, L.; Essiz, S.; Gulcin, I. Synthesis and biological evaluation of some 1-naphthol derivatives as antioxidants, acetylcholinesterase, carbonic anhydrase inhibitors. Arch. Pharm. 2021, 354, e2100113. [Google Scholar] [CrossRef]
- Zhao, N.; Shi, J.; Xu, H.; Luo, Q.; Li, Q.; Liu, M. Baicalin suppresses glaucoma pathogenesis by regulating the PI3K/AKT signaling in vitro and in vivo. Bioengineered 2021, 12, 10187–10198. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antioxidants | Fe3+ Reducing | Cu2+ Reducing | ||
|---|---|---|---|---|
| λ (700 nm) | r2 | λ (450 nm) | r2 | |
| BHA | 2.347 | 0.9086 | 2.216 | 0.9928 |
| BHT | 0.952 | 0.9154 | 2.044 | 0.9937 |
| Trolox | 2.119 | 0.9586 | 1.548 | 0.9305 |
| α-Tocopherol | 0.957 | 0.9863 | 0.816 | 0.9897 |
| Baicalin hydrate | 1.249 | 0.9848 | 0.344 | 0.9517 |
| Antioxidants | DPPH• Scavenging | ABTS•+ Scavenging | DMPD•+ Scavenging | |||
|---|---|---|---|---|---|---|
| IC50 | r2 | IC50 | r2 | IC50 | r2 | |
| BHA | 10.10 | 0.9015 | 5.07 | 0.9356 | 11.99 | 0.9580 |
| BHT | 25.95 | 0.9221 | 6.99 | 0.9350 | 8.72 | 0.9375 |
| Trolox | 7.05 | 0.9614 | 6.16 | 0.9692 | 4.33 | 0.9447 |
| α-Tocopherol | 11.31 | 0.9642 | 8.37 | 0.9015 | 7.11 | 0.9509 |
| Baicalin hydrate | 13.40 | 0.9940 | 38.37 | 0.9888 | 90.91 | 0.7410 |
| Compounds | IC50 (nM) | Ki (nM, n: 3) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CA II | r2 | AChE | r2 | BChE | r2 | α-Gly | r2 | CA II | AChE | BChE | α-Gly | |
| Baicalin hydrate | 31.13 | 0.9562 | 14.65 | 0.9758 | 12.31 | 0.9524 | 24.62 | 0.9524 | 19.25 ± 1.79 | 10.01 ± 2.86 | 3.50 ± 0.68 | 26.98 ± 9.91 |
| Acetazolamide * | 8.37 | 0.9825 | - | - | - | - | - | - | 4.41 ± 0.35 | - | - | - |
| Tacrine ** | - | - | 5.97 | 0.9706 | 8.37 | 0.9846 | - | - | - | 2.43 ± 0.92 | 5.99 ± 1.79 | - |
| Acarbose *** | - | - | - | - | - | - | 22,800 | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durmaz, L.; Karagecili, H.; Gulcin, İ. Evaluation of Carbonic Anhydrase, Acetylcholinesterase, Butyrylcholinesterase, and α-Glycosidase Inhibition Effects and Antioxidant Activity of Baicalin Hydrate. Life 2023, 13, 2136. https://doi.org/10.3390/life13112136
Durmaz L, Karagecili H, Gulcin İ. Evaluation of Carbonic Anhydrase, Acetylcholinesterase, Butyrylcholinesterase, and α-Glycosidase Inhibition Effects and Antioxidant Activity of Baicalin Hydrate. Life. 2023; 13(11):2136. https://doi.org/10.3390/life13112136
Chicago/Turabian StyleDurmaz, Lokman, Hasan Karagecili, and İlhami Gulcin. 2023. "Evaluation of Carbonic Anhydrase, Acetylcholinesterase, Butyrylcholinesterase, and α-Glycosidase Inhibition Effects and Antioxidant Activity of Baicalin Hydrate" Life 13, no. 11: 2136. https://doi.org/10.3390/life13112136