
Prolactin in Polycystic Ovary Syndrome: Metabolic Effects and Therapeutic Prospects

Abstract
:1. Introduction
1.1. PCOS Overview: Prevalence and Diagnostic Criteria
1.2. PCOS and Metabolic Implications: The Vicious Circle
1.3. PCOS: Diagnosis of Exclusion
1.4. Prolactin
1.5. Aim of the Review
2. The Emerging Role of Prolactin in Metabolic Homeostasis
2.1. Prolactin Synthesis and Regulation
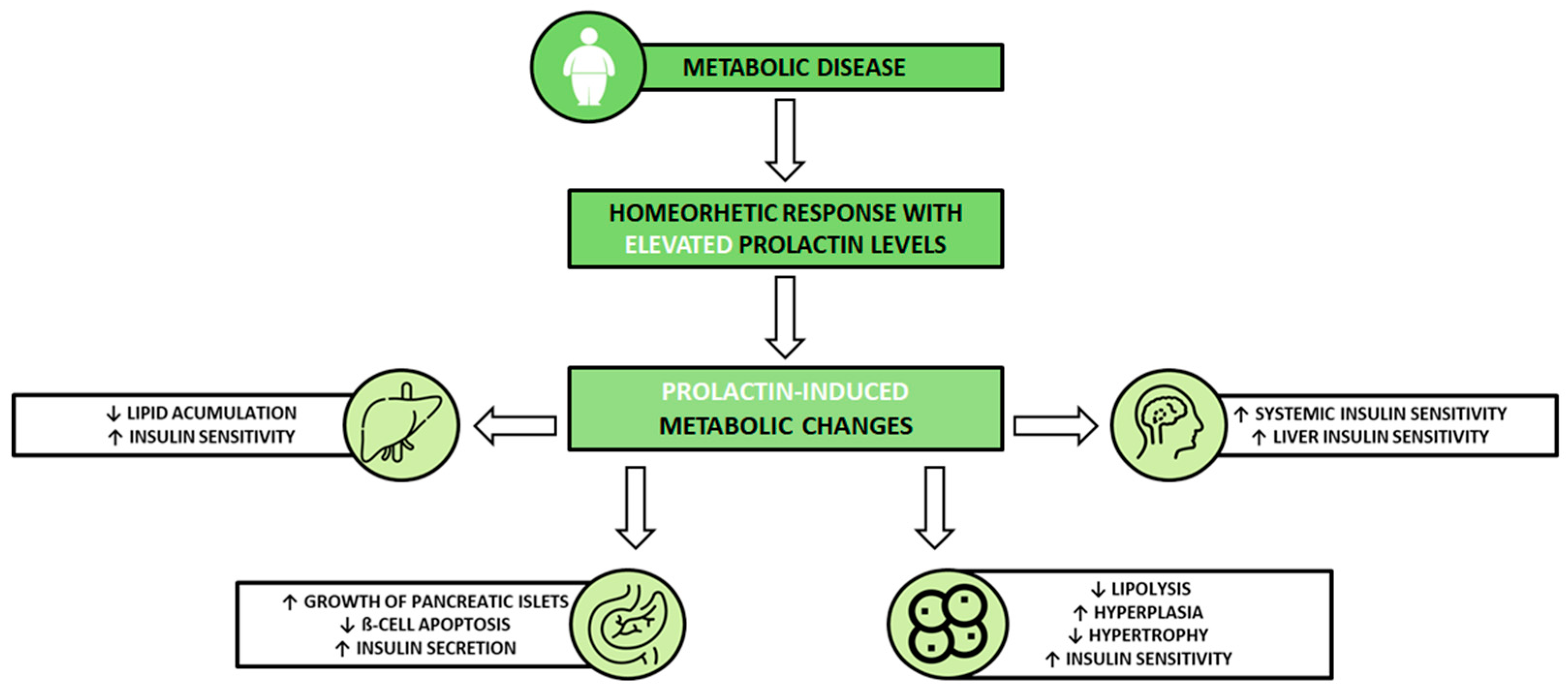
2.2. The Effects of Prolactin on Metabolism
2.3. Mechanisms Mediating the Metabolic Action of Prolactin
3. Prolactin and PCOS
3.1. Hyperprolactinemia and PCOS
3.2. Serum Prolactin Levels in PCOS Women Compared to Healthy Controls
3.3. Association between Serum PRL Levels and Markers of Metabolic Risk in PCOS
4. Role of Dopamine Agonists
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mohammad, M.B.; Seghinsara, A.M. Polycystic Ovary Syndrome (PCOS), Diagnostic Criteria, and AMH. Asian Pac. J. Cancer Prev. APJCP 2017, 18, 17–21. [Google Scholar] [CrossRef]
- Teede, H.; Deeks, A.; Moran, L. Polycystic Ovary Syndrome: A Complex Condition with Psychological, Reproductive and Metabolic Manifestations That Impacts on Health across the Lifespan. BMC Med. 2010, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Azziz, R.; Carmina, E.; Chen, Z.; Dunaif, A.; Laven, J.S.E.; Legro, R.S.; Lizneva, D.; Natterson-Horowtiz, B.; Teede, H.J.; Yildiz, B.O. Polycystic Ovary Syndrome. Nat. Rev. Dis. Primers 2016, 2, 16057. [Google Scholar] [CrossRef]
- Szydlarska, D.; Machaj, M.; Jakimiuk, A. History of Discovery of Polycystic Ovary Syndrome. Adv. Clin. Exp. Med. 2017, 26, 555–558. [Google Scholar] [CrossRef] [PubMed]
- Saei Ghare Naz, M.; Mousavi, M.; Mahboobifard, F.; Niknam, A.; Ramezani Tehrani, F. A Meta-Analysis of Observational Studies on Prolactin Levels in Women with Polycystic Ovary Syndrome. Diagnostics 2022, 12, 2924. [Google Scholar] [CrossRef]
- Teede, H.J.; Tay, C.T.; Laven, J.J.; Dokras, A.; Moran, L.J.; Piltonen, T.T.; Costello, M.F.; Boivin, J.; Redman, L.M.; Boyle, J.A.; et al. Recommendations from the 2023 International Evidence-Based Guideline for the Assessment and Management of Polycystic Ovary Syndrome. Eur. J. Endocrinol. 2023, 189, G43–G64. [Google Scholar] [CrossRef]
- Van der Ham, K.; Louwers, Y.V.; Laven, J.S.E. Cardiometabolic Biomarkers in Women with Polycystic Ovary Syndrome. Fertil. Steril. 2022, 117, 887–896. [Google Scholar] [CrossRef]
- Deswal, R.; Narwal, V.; Dang, A.; Pundir, C.S. The Prevalence of Polycystic Ovary Syndrome: A Brief Systematic Review. J. Hum. Reprod. Sci. 2020, 13, 261–271. [Google Scholar] [CrossRef]
- Singh, S.; Pal, N.; Shubham, S.; Sarma, D.K.; Verma, V.; Marotta, F.; Kumar, M. Polycystic Ovary Syndrome: Etiology, Current Management, and Future Therapeutics. J. Clin. Med. 2023, 12, 1454. [Google Scholar] [CrossRef]
- Mumusoglu, S.; Yildiz, B.O. Polycystic Ovary Syndrome Phenotypes and Prevalence: Differential Impact of Diagnostic Criteria and Clinical versus Unselected Population. Curr. Opin. Endocr. Metab. Res. 2020, 12, 66–71. [Google Scholar] [CrossRef]
- Pigny, P.; Merlen, E.; Robert, Y.; Cortet-Rudelli, C.; Decanter, C.; Jonard, S.; Dewailly, D. Elevated Serum Level of Anti-Mullerian Hormone in Patients with Polycystic Ovary Syndrome: Relationship to the Ovarian Follicle Excess and to the Follicular Arrest. J. Clin. Endocrinol. Metab. 2003, 88, 5957–5962. [Google Scholar] [CrossRef]
- Jeanes, Y.M.; Reeves, S. Metabolic Consequences of Obesity and Insulin Resistance in Polycystic Ovary Syndrome: Diagnostic and Methodological Challenges. Nutr. Res. Rev. 2017, 30, 97–105. [Google Scholar] [CrossRef]
- Filho, R.B.; Domingues, L.; Naves, L.; Ferraz, E.; Alves, A.; Casulari, L.A. Polycystic Ovary Syndrome and Hyperprolactinemia Are Distinct Entities. Gynecol. Endocrinol. 2007, 23, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Zhao, Q.; Zhao, X.; Xing, C.; He, B. Analysis of Endocrine and Metabolic Indexes in Non-Obese Patients with Polycystic Ovary Syndrome and Its Compare with Obese Patients. Diabetes Metab. Syndr. Obes. 2021, 14, 4275–4281. [Google Scholar] [CrossRef] [PubMed]
- Glintborg, D.; Altinok, M.; Mumm, H.; Buch, K.; Ravn, P.; Andersen, M. Prolactin Is Associated with Metabolic Risk and Cortisol in 1007 Women with Polycystic Ovary Syndrome. Hum. Reprod. 2014, 29, 1773–1779. [Google Scholar] [CrossRef]
- Herman, R.; Sikonja, J.; Jensterle, M.; Janez, A.; Dolzan, V. Insulin Metabolism in Polycystic Ovary Syndrome: Secretion, Signaling, and Clearance. Int. J. Mol. Sci. 2023, 24, 3140. [Google Scholar] [CrossRef] [PubMed]
- Janez, A.; Herman, R.; Poredos, P.; Mikhailidis, D.P.; Blinc, A.; Sabovic, M.; Studen, K.B.; Jezovnik, M.K.; Schernthaner, G.-H.; Anagnostis, P.; et al. Cardiometabolic Risk, Peripheral Arterial Disease and Cardiovascular Events in Polycystic Ovary Syndrome: Time to Implement Systematic Screening and Update the Management. Curr. Vasc. Pharmacol. 2023. [Google Scholar] [CrossRef]
- Christ, J.P.; Cedars, M.I. Current Guidelines for Diagnosing PCOS. Diagnostics 2023, 13, 1113. [Google Scholar] [CrossRef]
- Kyritsi, E.M.; Dimitriadis, G.K.; Kyrou, I.; Kaltsas, G.; Randeva, H.S. PCOS Remains a Diagnosis of Exclusion: A Concise Review of Key Endocrinopathies to Exclude. Clin. Endocrinol. 2017, 86, 1–6. [Google Scholar] [CrossRef]
- Majumdar, A.; Mangal, N.S. Hyperprolactinemia. J. Hum. Reprod. Sci. 2013, 6, 168–175. [Google Scholar] [CrossRef]
- Serafini, P.; Lobo, R.A. Prolactin Modulates Peripheral Androgen Metabolism. Fertil. Steril. 1986, 45, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Emanuel, R.H.K.; Roberts, J.; Docherty, P.D.; Lunt, H.; Campbell, R.E.; Möller, K. A Review of the Hormones Involved in the Endocrine Dysfunctions of Polycystic Ovary Syndrome and Their Interactions. Front. Endocrinol. 2022, 13, 1017468. [Google Scholar] [CrossRef] [PubMed]
- Bernard, V.; Young, J.; Binart, N. Prolactin—A Pleiotropic Factor in Health and Disease. Nat. Rev. Endocrinol. 2019, 15, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Vicchi, F.; De Winne, C.; Brie, B.; Sorianello, E.; Ladyman, S.R.; Becu-Villalobos, D. Metabolic Functions of Prolactin: Physiological and Pathological Aspects. J. Neuroendocrinol. 2020, 32, e12888. [Google Scholar] [CrossRef] [PubMed]
- Ben-Jonathan, N.; Hugo, E.R.; Brandebourg, T.D.; LaPensee, C.R. Focus on Prolactin as a Metabolic Hormone. Trends Endocrinol. Metab. 2006, 17, 110–116. [Google Scholar] [CrossRef]
- Macotela, Y.; Triebel, J.; Clapp, C. Time for a New Perspective on Prolactin in Metabolism. Trends Endocrinol. Metab. 2020, 31, 276–286. [Google Scholar] [CrossRef]
- Macotela, Y.; Ruiz-Herrera, X.; Vázquez-Carrillo, D.I.; Ramírez-Hernandez, G.; Martínez de la Escalera, G.; Clapp, C. The Beneficial Metabolic Actions of Prolactin. Front. Endocrinol. 2022, 13, 1001703. [Google Scholar] [CrossRef]
- Bernard, V.; Young, J.; Chanson, P.; Binart, N. New Insights in Prolactin: Pathological Implications. Nat. Rev. Endocrinol. 2015, 11, 265–275. [Google Scholar] [CrossRef]
- Kopelman, P. Physiopathology of Prolactin Secretion in Obesity. Int. J. Obes. 2000, 24, S104–S108. [Google Scholar] [CrossRef]
- Capozzi, A.; Scambia, G.; Pontecorvi, A.; Lello, S. Hyperprolactinemia: Pathophysiology and Therapeutic Approach. Gynecol. Endocrinol. 2015, 31, 506–510. [Google Scholar] [CrossRef]
- Harvey, S.; Martínez-Moreno, C.G.; Luna, M.; Arámburo, C. Autocrine/Paracrine Roles of Extrapituitary Growth Hormone and Prolactin in Health and Disease: An Overview. Gen. Comp. Endocrinol. 2015, 220, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Ben-Jonathan, N.; Mershon, J.L.; Allen, D.L.; Steinmetz, R.W. Extrapituitary Prolactin: Distribution, Regulation, Functions, and Clinical Aspects. Endocr. Rev. 1996, 17, 639–669. [Google Scholar] [CrossRef] [PubMed]
- Pirchio, R.; Graziadio, C.; Colao, A.; Pivonello, R.; Auriemma, R.S. Metabolic Effects of Prolactin. Front. Endocrinol. 2022, 13, 1015520. [Google Scholar] [CrossRef] [PubMed]
- Sadaria, R.G.; Ravi, B.V. Assessment of Leptin and Prolactin in Women with Polycystic Ovarian Syndrome. Int. J. Biochem. Res. Rev. 2019, 27, 1–7. [Google Scholar] [CrossRef]
- Albu, A.; Florea, S.; Fica, S. Is Prolactin the Missing Link in Adipose Tissue Dysfunction of Polycystic Ovary Syndrome Patients? Endocrine 2016, 51, 163–173. [Google Scholar] [CrossRef]
- Yang, Z.; She, J.; Zhou, C.; Liangshan, M. Association between Serum Prolactin and Metabolic Disorders: A Systematic Review. medRxiv [Preprint] 2020. [Google Scholar] [CrossRef]
- Kirsch, P.; Kunadia, J.; Shah, S.; Agrawal, N. Metabolic Effects of Prolactin and the Role of Dopamine Agonists: A Review. Front. Endocrinol. 2022, 13, 1002320. [Google Scholar] [CrossRef]
- Gorvin, C.M. The Prolactin Receptor: Diverse and Emerging Roles in Pathophysiology. J. Clin. Transl. Endocrinol. 2015, 2, 85–91. [Google Scholar] [CrossRef]
- Kondegowda, N.G.; Fenutria, R.; Pollack, I.R.; Orthofer, M.; Garcia-Ocaña, A.; Penninger, J.M.; Vasavada, R.C. Osteoprotegerin and Denosumab Stimulate Human Beta Cell Proliferation through Inhibition of the Receptor Activator of NF-ΚB Ligand Pathway. Cell Metab. 2015, 22, 77–85. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, X.; Gao, L.; Zhu, J.; Zhang, H.; Shi, H.; Woo, M.; Wu, X. Prolactin-Stimulated Survivin Induction Is Required for Beta Cell Mass Expansion during Pregnancy in Mice. Diabetologia 2015, 58, 2064–2073. [Google Scholar] [CrossRef]
- Banerjee, R.R.; Cyphert, H.A.; Walker, E.M.; Chakravarthy, H.; Peiris, H.; Gu, X.; Liu, Y.; Conrad, E.; Goodrich, L.; Stein, R.W.; et al. Gestational Diabetes Mellitus From Inactivation of Prolactin Receptor and MafB in Islet β-Cells. Diabetes 2016, 65, 2331–2341. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Toyofuku, Y.; Lynn, F.C.; Chak, E.; Uchida, T.; Mizukami, H.; Fujitani, Y.; Kawamori, R.; Miyatsuka, T.; Kosaka, Y.; et al. Serotonin Regulates Pancreatic Beta Cell Mass during Pregnancy. Nat. Med. 2010, 16, 804–808. [Google Scholar] [CrossRef] [PubMed]
- Terra, L.F.; Garay-Malpartida, M.H.; Wailemann, R.A.M.; Sogayar, M.C.; Labriola, L. Recombinant Human Prolactin Promotes Human Beta Cell Survival via Inhibition of Extrinsic and Intrinsic Apoptosis Pathways. Diabetologia 2011, 54, 1388–1397. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Snider, F.; Cross, J.C. Prolactin Receptor Is Required for Normal Glucose Homeostasis and Modulation of β-Cell Mass during Pregnancy. Endocrinology 2009, 150, 1618–1626. [Google Scholar] [CrossRef]
- Weinhaus, A.J.; Stout, L.E.; Sorenson, R.L. Glucokinase, Hexokinase, Glucose Transporter 2, and Glucose Metabolism in Islets during Pregnancy and Prolactin-Treated Islets in Vitro: Mechanisms for Long Term up-Regulation of Islets. Endocrinology 1996, 137, 1640–1649. [Google Scholar] [CrossRef]
- Weinhaus, A.J.; Stout, L.E.; Bhagroo, N.V.; Brelje, T.C.; Sorenson, R.L. Regulation of Glucokinase in Pancreatic Islets by Prolactin: A Mechanism for Increasing Glucose-Stimulated Insulin Secretion during Pregnancy. J. Endocrinol. 2007, 193, 367–381. [Google Scholar] [CrossRef]
- Zhang, P.; Ge, Z.; Wang, H.; Feng, W.; Sun, X.; Chu, X.; Jiang, C.; Wang, Y.; Zhu, D.; Bi, Y. Prolactin Improves Hepatic Steatosis via CD36 Pathway. J. Hepatol. 2018, 68, 1247–1255. [Google Scholar] [CrossRef]
- Shao, S.; Yao, Z.; Lu, J.; Song, Y.; He, Z.; Yu, C.; Zhou, X.; Zhao, L.; Zhao, J.; Gao, L. Ablation of Prolactin Receptor Increases Hepatic Triglyceride Accumulation. Biochem. Biophys. Res. Commun. 2018, 498, 693–699. [Google Scholar] [CrossRef]
- Jeyakumar, S.M.; Vajreswari, A. Stearoyl-CoA Desaturase 1: A Potential Target for Non-Alcoholic Fatty Liver Disease?-Perspective on Emerging Experimental Evidence. World J. Hepatol. 2022, 14, 168–179. [Google Scholar] [CrossRef]
- Yu, J.; Xiao, F.; Zhang, Q.; Liu, B.; Guo, Y.; Lv, Z.; Xia, T.; Chen, S.; Li, K.; Du, Y.; et al. PRLR Regulates Hepatic Insulin Sensitivity in Mice via STAT5. Diabetes 2013, 62, 3103–3113. [Google Scholar] [CrossRef]
- Berlanga, J.J.; Gualillo, O.; Buteau, H.; Applanat, M.; Kelly, P.A.; Edery, M. Prolactin Activates Tyrosyl Phosphorylation of Insulin Receptor Substrate 1 and Phosphatidylinositol-3-OH Kinase. J. Biol. Chem. 1997, 272, 2050–2052. [Google Scholar] [CrossRef] [PubMed]
- García-Martínez, J.M.; Calcabrini, A.; González, L.; Martín-Forero, E.; Agulló-Ortuño, M.T.; Simon, V.; Watkin, H.; Anderson, S.M.; Roche, S.; Martín-Pérez, J. A Non-Catalytic Function of the Src Family Tyrosine Kinases Controls Prolactin-Induced Jak2 Signaling. Cell. Signal. 2010, 22, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Xia, T.; Lv, Z.; Zhang, Q.; Xiao, Y.; Yu, J.; Liu, H.; Deng, J.; Guo, Y.; Wang, C.; et al. Central Prolactin Receptors (PRLRs) Regulate Hepatic Insulin Sensitivity in Mice via Signal Transducer and Activator of Transcription 5 (STAT5) and the Vagus Nerve. Diabetologia 2014, 57, 2136–2144. [Google Scholar] [CrossRef] [PubMed]
- Ling, C.; Svensson, L.; Odén, B.; Weijdegård, B.; Edén, B.; Edén, S.; Billig, H. Identification of Functional Prolactin (PRL) Receptor Gene Expression: PRL Inhibits Lipoprotein Lipase Activity in Human White Adipose Tissue. J. Clin. Endocrinol. Metab. 2003, 88, 1804–1808. [Google Scholar] [CrossRef] [PubMed]
- LaPensee, C.R.; Horseman, N.D.; Tso, P.; Brandebourg, T.D.; Hugo, E.R.; Ben-Jonathan, N. The Prolactin-Deficient Mouse Has an Unaltered Metabolic Phenotype. Endocrinology 2006, 147, 4638–4645. [Google Scholar] [CrossRef] [PubMed]
- Nanbu-Wakao, R.; Fujitani, Y.; Masuho, Y.; Muramatu, M.-A.; Wakao, H. Prolactin Enhances CCAAT Enhancer-Binding Protein-β(C/EBPβ) and Peroxisome Proliferator-Activated Receptor γ(PPARγ) Messenger RNA Expression and Stimulates Adipogenic Conversion of NIH-3T3 Cells. Mol. Endocrinol. 2000, 14, 307–316. [Google Scholar] [CrossRef]
- Stewart, W.C.; Baugh, J.E.; Floyd, Z.E.; Stephens, J.M. STAT 5 Activators Can Replace the Requirement of FBS in the Adipogenesis of 3T3-L1 Cells. Biochem. Biophys. Res. Commun. 2004, 324, 355–359. [Google Scholar] [CrossRef]
- Ruiz-Herrera, X.; de los Ríos, E.A.; Díaz, J.M.; Lerma-Alvarado, R.M.; Martínez de la Escalera, L.; López-Barrera, F.; Lemini, M.; Arnold, E.; Martínez de la Escalera, G.; Clapp, C.; et al. Prolactin Promotes Adipose Tissue Fitness and Insulin Sensitivity in Obese Males. Endocrinology 2017, 158, 56–68. [Google Scholar] [CrossRef]
- Ponce, A.J.; Galván-Salas, T.; Lerma-Alvarado, R.M.; Ruiz-Herrera, X.; Hernández-Cortés, T.; Valencia-Jiménez, R.; Cárdenas-Rodríguez, L.E.; Martínez De La Escalera, G.; Clapp, C.; Macotela, Y. Low Prolactin Levels Are Associated with Visceral Adipocyte Hypertrophy and Insulin Resistance in Humans. Endocrine 2020, 67, 331–343. [Google Scholar] [CrossRef]
- Nguyen, T.M.D. Adiponectin: Role in Physiology and Pathophysiology. Int. J. Prev. Med. 2020, 11, 136. [Google Scholar] [CrossRef]
- Berridge, B.R. Animal Study Translation: The Other Reproducibility Challenge. ILAR J. 2021, 62, 1–6. [Google Scholar] [CrossRef]
- Ben-Jonathan, N.; LaPensee, C.R.; LaPensee, E.W. What Can We Learn from Rodents about Prolactin in Humans? Endocr. Rev. 2008, 29, 1–41. [Google Scholar] [CrossRef]
- Melmed, S.; Casanueva, F.F.; Hoffman, A.R.; Kleinberg, D.L.; Montori, V.M.; Schlechte, J.A.; Wass, J.A.H. Diagnosis and Treatment of Hyperprolactinemia: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2011, 96, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Delcour, C.; Robin, G.; Young, J.; Dewailly, D. PCOS and Hyperprolactinemia: What Do We Know in 2019? Clin. Med. Insights Reprod. Health 2019, 13, 117955811987192. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.I.; Yoon, J.H.; Park, D.C.; Yang, S.H.; Kim, Y.I. What Is the Optimal Prolactin Cutoff for Predicting the Presence of a Pituitary Adenoma in Patients with Polycystic Ovary Syndrome? Int. J. Med. Sci. 2023, 20, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Davoudi, Z.; Araghi, F.; Vahedi, M.; Mokhtari, N.; Gheisari, M. Prolactin Level in Polycystic Ovary Syndrome (PCOS): An Approach to the Diagnosis and Management. Acta Biomed. Atenei Parm. 2021, 92, e2021291. [Google Scholar] [CrossRef]
- Rodier, C.; Courbière, B.; Fernandes, S.; Vermalle, M.; Florence, B.; Resseguier, N.; Brue, T.; Cuny, T. Metoclopramide Test in Hyperprolactinemic Women With Polycystic Ovarian Syndrome: Old Wine Into New Bottles? Front. Endocrinol. 2022, 13, 832361. [Google Scholar] [CrossRef]
- Mahboobifard, F.; Rahmati, M.; Amiri, M.; Azizi, F.; Ramezani Tehrani, F. To What Extent Does Polycystic Ovary Syndrome Influence the Cut-off Value of Prolactin? Findings of a Community-Based Study. Adv. Med. Sci. 2022, 67, 79–86. [Google Scholar] [CrossRef]
- Abdelazim, I.; Amer, O.; Farghali, M.M. Common Endocrine Disorders Associated with the Polycystic Ovary Syndrome. Menopausal Rev. 2020, 19, 179–183. [Google Scholar] [CrossRef]
- Hayashida, S.A.Y.; Marcondes, J.A.M.; Soares, J.M.; Rocha, M.P.; Barcellos, C.R.G.; Kobayashi, N.K.A.; Baracat, E.C.; Maciel, G.A.R. Evaluation of Macroprolactinemia in 259 Women under Investigation for Polycystic Ovary Syndrome. Clin. Endocrinol. 2014, 80, 616–618. [Google Scholar] [CrossRef]
- Kyritsi, E.M.; Dimitriadis, G.K.; Angelousi, A.; Mehta, H.; Shad, A.; Mytilinaiou, M.; Kaltsas, G.; Randeva, H.S. The Value of Prolactin in Predicting Prolactinοma in Hyperprolactinaemic Polycystic Ovarian Syndrome. Eur. J. Clin. Investig. 2018, 48, e12961. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, A.P.; Dunlop, W.; Kendall-Taylor, P. Studies of Prolactin Secretion in Polycystic OVARY syndrome. Clin. Endocrinol. 1986, 24, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Minakami, H.; Abe, N.; Oka, N.; Kimura, K.; Tamura, T.; Tamada, T. Prolactin Release in Polycystic Ovarian Syndrome. Endocrinol. Jpn. 1988, 35, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Szosland, K.; Pawłowicz, P.; Lewiński, A. Prolactin Secretion in Polycystic Ovary Syndrome (PCOS). Neuro Endocrinol Lett. 2015, 36, 53–58. [Google Scholar]
- Ben Salem, A.; Megdich, F.; Kacem, O.; Souayeh, M.; Hachani Ben Ali, F.; Hizem, S.; Janhai, F.; Ajina, M.; Abu-Elmagd, M.; Assidi, M.; et al. Vascular Endothelial Growth Factor (VEGFA) Gene Variation in Polycystic Ovary Syndrome in a Tunisian Women Population. BMC Genom. 2016, 17, 748. [Google Scholar] [CrossRef]
- Yang, H.; Di, J.; Pan, J.; Yu, R.; Teng, Y.; Cai, Z.; Deng, X. The Association Between Prolactin and Metabolic Parameters in PCOS Women: A Retrospective Analysis. Front. Endocrinol. 2020, 11, 263. [Google Scholar] [CrossRef]
- Yang, H.; Lin, J.; Li, H.; Liu, Z.; Chen, X.; Chen, Q. Prolactin Is Associated With Insulin Resistance and Beta-Cell Dysfunction in Infertile Women With Polycystic Ovary Syndrome. Front. Endocrinol. 2021, 12, 571229. [Google Scholar] [CrossRef]
- Das, M.; Gogoi, C. Pool vs Single Sample Determination of Serum Prolactin to Explore Venipuncture Associated Stress Induced Variation. Sci. Rep. 2023, 13, 113. [Google Scholar] [CrossRef]
- Chaudhari, N.; Dawalbhakta, M.; Nampoothiri, L. GnRH Dysregulation in Polycystic Ovarian Syndrome (PCOS) Is a Manifestation of an Altered Neurotransmitter Profile. Reprod. Biol. Endocrinol. 2018, 16, 37. [Google Scholar] [CrossRef]
- Amin, M.; Horst, N.; Gragnoli, C. Linkage and Association of Variants in the Dopamine Receptor 2 Gene (DRD2) with Polycystic Ovary Syndrome. J. Ovarian Res. 2023, 16, 158. [Google Scholar] [CrossRef]
- McCartney, C.R.; Campbell, R.E. Abnormal GnRH Pulsatility in Polycystic Ovary Syndrome: Recent Insights. Curr. Opin. Endocr. Metab. Res. 2020, 12, 78–84. [Google Scholar] [CrossRef]
- Xing, L.; Xu, J.; Wei, Y.; Chen, Y.; Zhuang, H.; Tang, W.; Yu, S.; Zhang, J.; Yin, G.; Wang, R.; et al. Depression in Polycystic Ovary Syndrome: Focusing on Pathogenesis and Treatment. Front. Psychiatry 2022, 13, 1001484. [Google Scholar] [CrossRef]
- Abbara, A.; Dhillo, W.S. Targeting Elevated GnRH Pulsatility to Treat Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2021, 106, e4275–e4277. [Google Scholar] [CrossRef] [PubMed]
- Laatikainen, T.; Tulenheimo, A. Prolactin Pulsatility in Polycystic Ovarian Disease. J. Endocrinol. Investig. 1985, 8, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Venturoli, S.; Porcu, E.; Fabbri, R.; Magrini, O.; Gammi, L.; Paradisi, R.; Forcacci, M.; Bolzani, R.; Flamigni, C. Episodic Pulsatile Secretion of Fsh, Lh, Prolactin, Oestradiol, Oestrone, and Lh Circadian Variations in Polycystic Ovary Syndrome. Clin. Endocrinol. 1988, 28, 93–107. [Google Scholar] [CrossRef]
- Kalro, B.N.; Loucks, T.L.; Berga, S.L. Neuromodulation in Polycystic Ovary Syndrome. Obstet. Gynecol. Clin. North Am. 2001, 28, 35–62. [Google Scholar] [CrossRef] [PubMed]
- Gómez, R.; Ferrero, H.; Delgado-Rosas, F.; Gaytan, M.; Morales, C.; Zimmermann, R.C.; Simón, C.; Gaytan, F.; Pellicer, A. Evidences for the Existence of a Low Dopaminergic Tone in Polycystic Ovarian Syndrome: Implications for OHSS Development and Treatment. J. Clin. Endocrinol. Metab. 2011, 96, 2484–2492. [Google Scholar] [CrossRef]
- Hernández, I.; Parra, A.; Méndez, I.; Cabrera, V.; del Carmen Cravioto, M.; Mercado, M.; Díaz-Sánchez, V.; Larrea, F. Hypothalamic Dopaminergic Tone and Prolactin Bioactivity in Women with Polycystic Ovary Syndrome. Arch. Med. Res. 2000, 31, 216–222. [Google Scholar] [CrossRef]
- Edinoff, A.N.; Silverblatt, N.S.; Vervaeke, H.E.; Horton, C.C.; Girma, E.; Kaye, A.D.; Kaye, A.; Kaye, J.S.; Garcia, A.J.; Neuchat, E.E.; et al. Hyperprolactinemia, Clinical Considerations, and Infertility in Women on Antipsychotic Medications. Psychopharmacol. Bull. 2021, 51, 131–148. [Google Scholar]
- Wang, G.-J.; Volkow, N.D.; Logan, J.; Pappas, N.R.; Wong, C.T.; Zhu, W.; Netusll, N.; Fowler, J.S. Brain Dopamine and Obesity. Lancet 2001, 357, 354–357. [Google Scholar] [CrossRef]
- Ruegsegger, G.N.; Booth, F.W. Running from Disease: Molecular Mechanisms Associating Dopamine and Leptin Signaling in the Brain with Physical Inactivity, Obesity, and Type 2 Diabetes. Front. Endocrinol. 2017, 8, 109. [Google Scholar] [CrossRef] [PubMed]
- Benrick, A.; Chanclón, B.; Micallef, P.; Wu, Y.; Hadi, L.; Shelton, J.M.; Stener-Victorin, E.; Wernstedt Asterholm, I. Adiponectin Protects against Development of Metabolic Disturbances in a PCOS Mouse Model. Proc. Natl. Acad. Sci. USA 2017, 114, E7187–E7196. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.-Y.; Sung, Y.-A.; Lee, H.J. The Visceral Adiposity Index as a Predictor of Insulin Resistance in Young Women with Polycystic Ovary Syndrome. Obesity 2013, 21, 1690–1694. [Google Scholar] [CrossRef] [PubMed]
- Kok, P.; Roelfsema, F.; Frölich, M.; Meinders, A.E.; Pijl, H. Prolactin Release Is Enhanced in Proportion to Excess Visceral Fat in Obese Women. J. Clin. Endocrinol. Metab. 2004, 89, 4445–4449. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, Y.; Ozaksit, G.; Serdar Unlu, B.; Ozgu, E.; Energin, H.; Kaba, M.; Ugur, M. Serum Adiponectin Level and Clinical, Metabolic, and Hormonal Markers in Patients with Polycystic Ovary Syndrome. Int. J. Fertil. Steril. 2014, 7, 331–336. [Google Scholar] [PubMed]
- Hajer, G.R.; van Haeften, T.W.; Visseren, F.L.J. Adipose Tissue Dysfunction in Obesity, Diabetes, and Vascular Diseases. Eur. Heart J. 2008, 29, 2959–2971. [Google Scholar] [CrossRef]
- Amato, M.C.; Giordano, C. Visceral Adiposity Index: An Indicator of Adipose Tissue Dysfunction. Int. J. Endocrinol. 2014, 2014, 730827. [Google Scholar] [CrossRef]
- Sanchez-Garrido, M.A.; Tena-Sempere, M. Metabolic Dysfunction in Polycystic Ovary Syndrome: Pathogenic Role of Androgen Excess and Potential Therapeutic Strategies. Mol. Metab. 2020, 35, 100937. [Google Scholar] [CrossRef]
- Choi, J.; Horner, K.A. Dopamine Agonists. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Nunes-Nogueira, V.S.; Gomes, P.L.; da Silva, E.M.; Atallah, Á.N. Dopamine Agonists for Idiopathic Hyperprolactinaemia and Prolactinoma in Adults. Cochrane Database Syst. Rev. 2018, 2018, CD009718. [Google Scholar] [CrossRef]
- Ignacak, A.; Kasztelnik, M.; Sliwa, T.; Korbut, R.A.; Rajda, K.; Guzik, T.J. Prolactin--Not Only Lactotrophin. A “New” View of the “Old” Hormone. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2012, 63, 435–443. [Google Scholar]
- Al-Husaynei, A.J.; Mahmood, I.H.; Al-Jubori, Z.S. Comparison of the Effects of Cabergoline and Bromocriptine in Women with Hyperprolactinemic Amenorrhea. Middle East Fertil. Soc. J. 2008, 13, 33–38. [Google Scholar]
- Liu, X.; Tang, C.; Wen, G.; Zhong, C.; Yang, J.; Zhu, J.; Ma, C. The Mechanism and Pathways of Dopamine and Dopamine Agonists in Prolactinomas. Front. Endocrinol. 2019, 9, 768. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, C.M.; Milsom, S.R. Dopamine Agonists in the Treatment of Prolactinoma: Are They Still First Choice? Intern. Med. J. 2011, 41, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Ke, X.; Wang, L.; Chen, M.; Liu, S.; Yu, N.; Duan, L.; Gong, F.; Zhu, H. The Side Effects of Dopamine Receptor Agonist Drugs in Chinese Prolactinoma Patients: A Cross Sectional Study. BMC Endocr. Disord. 2022, 22, 97. [Google Scholar] [CrossRef]
- Ioachimescu, A.G.; Fleseriu, M.; Hoffman, A.R.; Vaughan, T.B., III; Katznelson, L. Psychological Effects of Dopamine Agonist Treatment in Patients with Hyperprolactinemia and Prolactin-Secreting Adenomas. Eur. J. Endocrinol. 2019, 180, 31–40. [Google Scholar] [CrossRef]
- Tran, T.; Brophy, J.M.; Suissa, S.; Renoux, C. Risks of Cardiac Valve Regurgitation and Heart Failure Associated with Ergot- and Non-Ergot-Derived Dopamine Agonist Use in Patients with Parkinson’s Disease: A Systematic Review of Observational Studies. CNS Drugs 2015, 29, 985–998. [Google Scholar] [CrossRef]
- Bancos, I.; Nannenga, M.R.; Bostwick, J.M.; Silber, M.H.; Erickson, D.; Nippoldt, T.B. Impulse Control Disorders in Patients with Dopamine Agonist-Treated Prolactinomas and Nonfunctioning Pituitary Adenomas: A Case–Control Study. Clin. Endocrinol. 2014, 80, 863–868. [Google Scholar] [CrossRef]
- Yavasoglu, I.; Kucuk, M.; Coskun, A.; Guney, E.; Kadikoylu, G.; Bolaman, Z. Polycystic Ovary Syndrome and Prolactinoma Association. Intern. Med. 2009, 48, 611–613. [Google Scholar] [CrossRef]
- Falaschi, P.; Rocco, A.; del Pozo, E. Inhibitory Effect of Bromocriptine Treatment on Luteinizing Hormone Secretion in Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 1986, 62, 348–351. [Google Scholar] [CrossRef]
- Papaleo, E.; Doldi, N.; De Santis, L.; Marelli, G.; Marsiglio, E.; Rofena, S.; Ferrari, A. Cabergoline Influences Ovarian Stimulation in Hyperprolactinaemic Patients with Polycystic Ovary Syndrome. Hum. Reprod. 2001, 16, 2263–2266. [Google Scholar] [CrossRef]
- Ghaneei, A.; Jowkar, A.; Hasani Ghavam, M.R.; Ghaneei, E. Cabergoline plus Metformin Therapy Effects on Menstrual Irregularity and Androgen System in Polycystic Ovary Syndrome Women with Hyperprolactinemia. Iran J. Reprod. Med. 2015, 13, 93–100. [Google Scholar] [PubMed]
- Elsersy, M.A.M. Efficacy of Combined Cabergoline and Metformin Compared to Metformin Alone on Cycle Regularity in Patients with Polycystic Ovarian Disease with Hyperprolactinemia: A Randomized Clinical Trial. J. Obstet. Gynecol. India 2017, 67, 363–369. [Google Scholar] [CrossRef]
- Spruce, B.A.; Kendall-Taylor, P.; Dunlop, W.; Anderson, A.J.; Watson, M.J.; Cook, D.B.; Gray, C. The Effect of Bromocriptine in the Polycystic Ovary Syndrome. Clin. Endocrinol. 1984, 20, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, A.M.; Cagnacci, A.; Depau, G.F.; Orrù, M.; Ajossa, S.; Melis, G.B. The Chronic Administration of Cabergoline Normalizes Androgen Secretion and Improves Menstrual Cyclicity in Women with Polycystic Ovary Syndrome. Fertil. Steril. 1996, 66, 527–532. [Google Scholar] [CrossRef]
- Steingold, K.A.; Lobo, R.A.; Judd, H.L.; Lu, J.K.H.; Chang, R.J. The Effect of Bromocriptine on Gonadotropin and Steroid Secretion in Polycystic Ovarian Disease*. J. Clin. Endocrinol. Metab. 1986, 62, 1048–1051. [Google Scholar] [CrossRef]
- Parsanezhad, M.E.; Alborzi, S.; Jahromi, B.N. A Prospective, Double-Blind, Randomized, Placebo-Controlled Clinical Trial of Bromocriptine in Clomiphene-Resistant Patients with Polycystic Ovary Syndrome and Normal Prolactin Level. Int. J. Fertil. Womens Med. 2002, 47, 272–277. [Google Scholar] [PubMed]
- Prelević, G.M.; Würzburger, M.I.; Perić, L.A. Acute Effects of L-Dopa and Bromocriptine on Serum PRL, LH and FSH Levels in Patients with Hyperprolactinemic and Normoprolactinemic Polycystic Ovary Syndrome. J. Endocrinol. Investig. 1987, 10, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Chien, H.-Y.; Chen, S.-M.; Li, W.-C. Dopamine Receptor Agonists Mechanism of Actions on Glucose Lowering and Their Connections with Prolactin Actions. Front. Clin. Diabetes Healthc. 2023, 4, 935872. [Google Scholar] [CrossRef]


{kind=link}
| Study | Sample Size, Participants | Age (Years) | BMI of Participants (kg/m2) | PRL (μg/L) | Main Outcomes |
|---|---|---|---|---|---|
| Glintborg, 2014 [15] | 1007, patients with PCOS | 30 (23–36) | 27.4 (23.2–33) | 7 (5–10) | PRL was inversely associated with WC (r = −0.13, p < 0.05), TC (−0.13, p < 0.05), LDL-C (r = −0.15, p < 0.05), TG (r = −0.14, p < 0.05), and positively with HDL-C (r = 0.11, p < 0.05) |
| Albu, 2016 [35] | 322, patients with PCOS | 24 (IQR 7) | 27 (IQR 11.8) | 12.9 (IQR 8.25) | PRL was inversely associated with HOMA-IR (r = −0.185, p = 0.002), BMI (r = −0.194, p < 0.0001), WC (r = −0.217, p < 0.0001), and fasting glucose (r = −0.123, p = 0.037) |
| Yang, 2020 [76] | 2052, infertile patients with PCOS | 29.12 ± 0.63 | 22.8 ± 1.2 | 11.71 ± 1.92 | PRL was inversely associated with BMI (r = −0.015, p < 0.01), TG (r = −0.067, p < 0.01), TC (r = −0.089, p < 0.01), LDL-C (r = −0.074, p < 0.01) |
| Yang, 2021 [77] | 792, infertile patients with PCOS | 29 (27–32.5) | 23.73 (21.48–26.85) | 11.08 (8.78–14.95) | PRL was inversely associated with BMI (r = −0.086, p < 0.05), WC (r = −0.302, p < 0.001), TG (r = −0.107, p < 0.05), FINS (r = −0.152, p < 0.001), HOMA-IR (r = −0.144, p < 0.001), HOMA-ß (r = −0.165, p < 0.001), and positively with HDL-C (r = 0.084, p < 0.05) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastnak, L.; Herman, R.; Ferjan, S.; Janež, A.; Jensterle, M. Prolactin in Polycystic Ovary Syndrome: Metabolic Effects and Therapeutic Prospects. Life 2023, 13, 2124. https://doi.org/10.3390/life13112124
Mastnak L, Herman R, Ferjan S, Janež A, Jensterle M. Prolactin in Polycystic Ovary Syndrome: Metabolic Effects and Therapeutic Prospects. Life. 2023; 13(11):2124. https://doi.org/10.3390/life13112124
Chicago/Turabian StyleMastnak, Lara, Rok Herman, Simona Ferjan, Andrej Janež, and Mojca Jensterle. 2023. "Prolactin in Polycystic Ovary Syndrome: Metabolic Effects and Therapeutic Prospects" Life 13, no. 11: 2124. https://doi.org/10.3390/life13112124