

Molecular Identification of Protozoan Sarcocystis in Different Types of Water Bodies in Lithuania

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
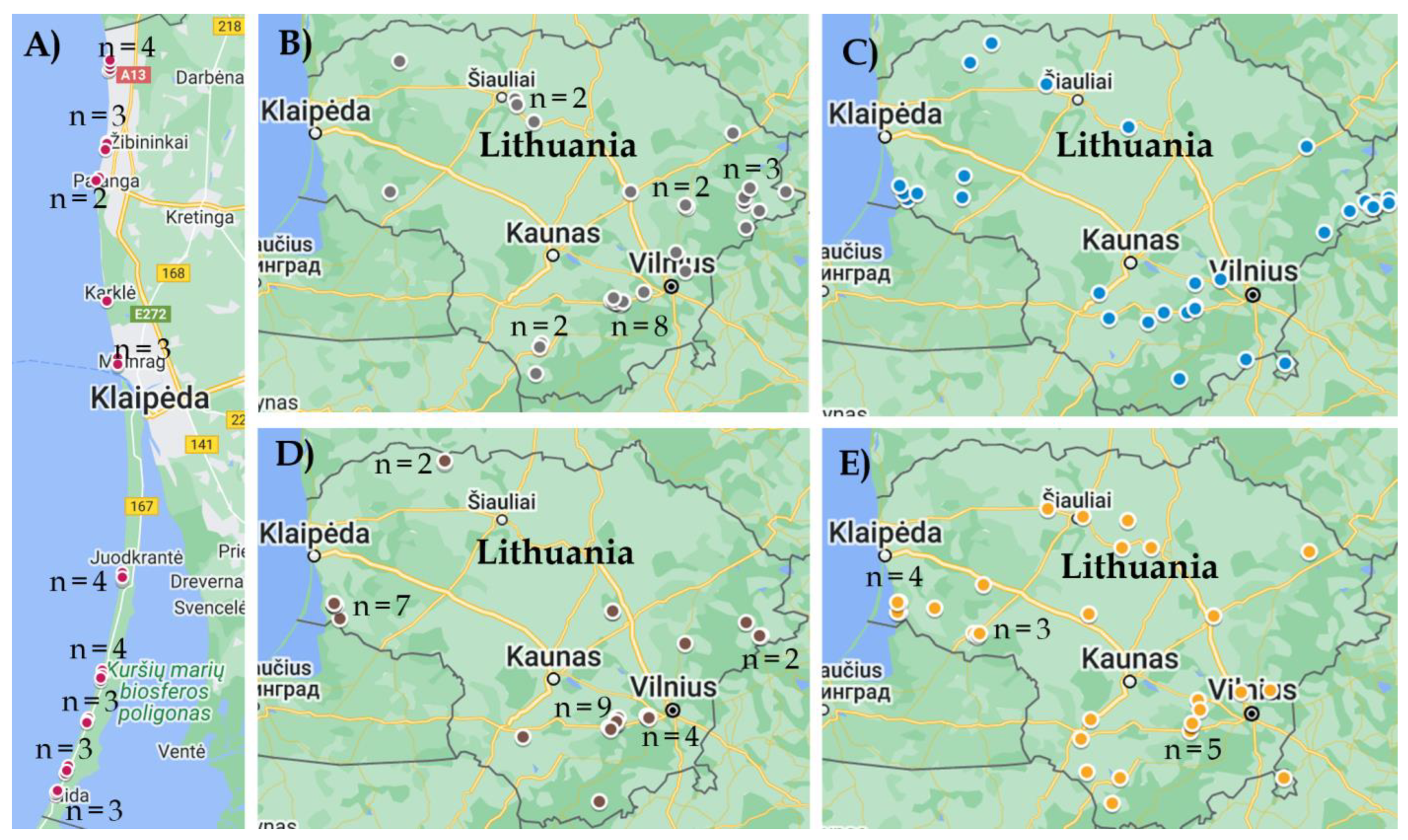
2.1. Sample Collection
2.2. Preparation of Water Samples and Genomic DNA Extraction
2.3. Nested PCR-Based Identification of Sarcocystis Parasites
3. Results
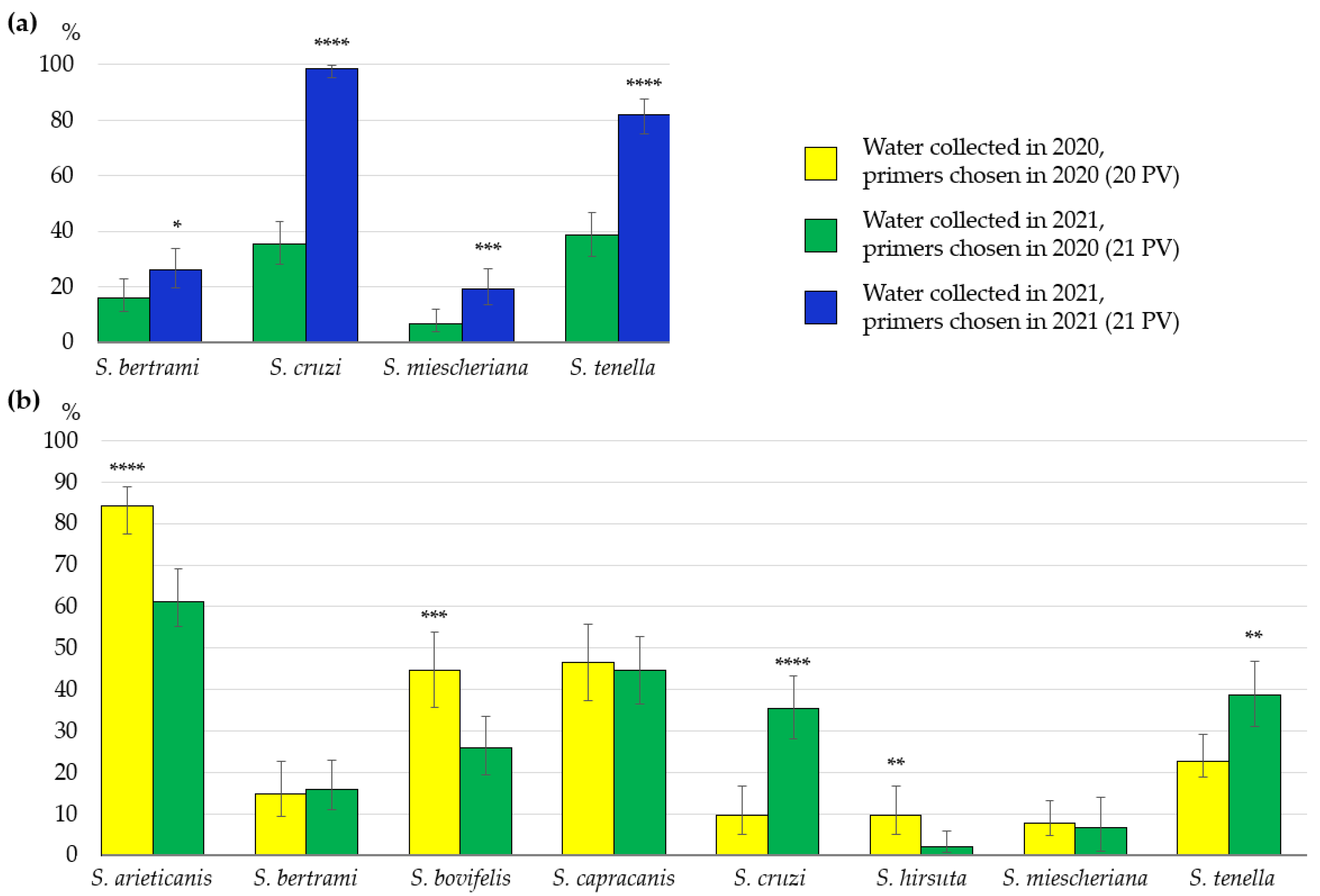
3.1. Identification of Sarcocystis spp. Using Different PCR Primer Sets
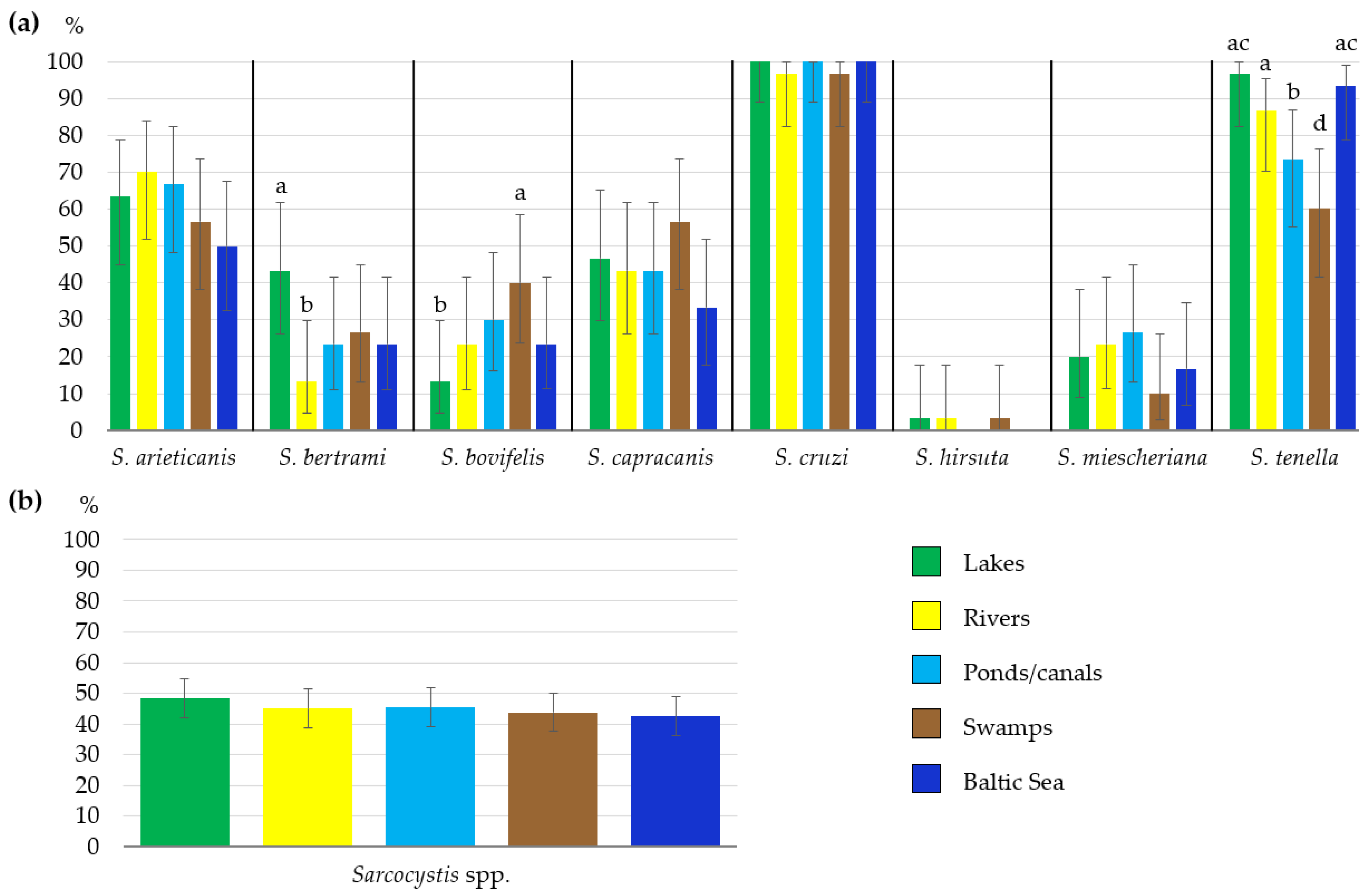
3.2. Sarcocystis spp. Occurrence Rates in Different Types of Water Bodies
3.3. Distribution of Sarcocystis spp. in Water Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nimri, L. Unusual case presentation of intestinal Sarcocystis hominis infection in a healthy adult. JMM Case Rep. 2014, 1, e004069. [Google Scholar] [CrossRef] [Green Version]
- Castro-Forero, S.P.; Bulla-Castañeda, D.M.B.-C.D.M.; Buitrago, H.A.L.; Anaya, A.M.D.; de Carvalho, L.M.M.; Pulido-Medellín, M.O. Sarcocystis spp., a parasite with zoonotic potential. Bulg. J. Veter.-Med. 2022, 25, 175–186. [Google Scholar] [CrossRef]
- Rubiola, S.; Civera, T.; Panebianco, F.; Vercellino, D.; Chiesa, F. Molecular detection of cattle Sarcocystis spp. in North-West Italy highlights their association with bovine eosinophilic myositis. Parasites Vectors 2021, 14, 223. [Google Scholar] [CrossRef]
- Yan, W.-C.; Qian, W.; Li, X.; Wang, T.; Ding, K.; Huang, T. Morphological and molecular characterization of Sarcocystis miescheriana from pigs in the central region of China. Parasitol. Res. 2013, 112, 975–980. [Google Scholar] [CrossRef]
- Dubey, J.P.; Bernal, R.C.; Rosenthal, B.; Speer, C.; Fayer, R. Sarcocystosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Dong, H.; Su, R.; Wang, Y.; Tong, Z.; Zhang, L.; Yang, Y.; Hu, J. Sarcocystis species in wild and domestic sheep (Ovis ammon and Ovis aries) from China. BMC Veter.-Res. 2018, 14, 377. [Google Scholar] [CrossRef]
- Aráoz, V.; Silveira, C.D.S.; Moré, G.; Banchero, G.; Riet-Correa, F.; Giannitti, F. Fatal Sarcocystis cruzi—induced eosinophilic myocarditis in a heifer in Uruguay. J. Veter.-Diagn. Investig. 2019, 31, 656–660. [Google Scholar] [CrossRef]
- Shahari, S.; Tengku-Idris, T.I.N.; Fong, M.Y.; Lau, Y.L. Molecular evidence of Sarcocystis nesbitti in water samples of Tioman Island, Malaysia. Parasites Vectors 2016, 9, 598. [Google Scholar] [CrossRef] [Green Version]
- Lee, F.C.H. Finding Sarcocystis spp. on the Tioman Island: 28S rRNA gene next-generation sequencing reveals nine new Sarcocystis species. J. Water Health 2019, 17, 416–427. [Google Scholar] [CrossRef] [Green Version]
- Strazdaitė-Žielienė, Ž.; Baranauskaitė, A.; Butkauskas, D.; Servienė, E.; Prakas, P. Molecular Identification of Parasitic Protozoa Sarcocystis in Water Samples. Veter.-Sci. 2022, 9, 412. [Google Scholar] [CrossRef]
- Gjerde, B. Molecular characterisation of Sarcocystis bovifelis, Sarcocystis bovini n. sp., Sarcocystis hirsuta and Sarcocystis cruzi from cattle (Bos taurus) and Sarcocystis sinensis from water buffaloes (Bubalus bubalis). Parasitol. Res. 2016, 115, 1473–1492. [Google Scholar] [CrossRef]
- Huang, Z.; Ye, Y.; Zhang, H.; Deng, S.; Tao, J.; Hu, J.; Yang, Y. Morphological and molecular characterizations of Sarcocystis miescheriana and Sarcocystis suihominis in domestic pigs (Sus scrofa) in China. Parasitol. Res. 2019, 118, 3491–3496. [Google Scholar] [CrossRef]
- Metwally, D.M.; Al-Damigh, M.A.; Al-Turaiki, I.M.; El-Khadragy, M.F. Molecular Characterization of Sarcocystis Species Isolated from Sheep and Goats in Riyadh, Saudi Arabia. Animals 2019, 9, 256. [Google Scholar] [CrossRef] [Green Version]
- Januskevicius, V.; Januskeviciene, G.; Prakas, P.; Butkauskas, D.; Petkevicius, S. Prevalence and intensity of Sarcocystis spp. infection in animals slaughtered for food in Lithuania. Veter. Med. 2019, 64, 149–157. [Google Scholar] [CrossRef]
- Prakas, P.; Strazdaitė-Žielienė, Ž.; Januškevičius, V.; Chiesa, F.; Baranauskaitė, A.; Rudaitytė-Lukošienė, E.; Servienė, E.; Petkevičius, S.; Butkauskas, D. Molecular identification of four Sarcocystis species in cattle from Lithuania, including S. hominis, and development of a rapid molecular detection method. Parasites Vectors 2020, 13, 610. [Google Scholar] [CrossRef]
- Prakas, P.; Kirillova, V.; Dzerkale, A.; Kirjušina, M.; Butkauskas, D.; Gavarāne, I.; Rudaitytė-Lukošienė, E.; Šulinskas, G. First molecular characterization of Sarcocystis miescheriana in wild boars (Sus scrofa) from Latvia. Parasitol. Res. 2020, 119, 3777–3783. [Google Scholar] [CrossRef]
- Marandykina-Prakienė, A.; Butkauskas, D.; Gudiškis, N.; Juozaitytė-Ngugu, E.; Januškevičius, V.; Rudaitytė-Lukošienė, E.; Prakas, P. Molecular Identification of Sarcocystis Species in Sheep from Lithuania. Animals 2022, 12, 2048. [Google Scholar] [CrossRef]
- Cabaj, W.; Grzelak, S.; Moskwa, B.; Bień-Kalinowska, J. Sarcocystis cruzi infection in free-living European bison (Bison bonasus bonasus L.) from the Białowieża Forest, Poland—A molecular analysis based on the cox1 gene. Int. J. Parasitol. Parasites Wildl. 2021, 16, 59–63. [Google Scholar] [CrossRef]
- Prakas, P.; Rehbein, S.; Rudaitytė-Lukošienė, E.; Butkauskas, D. Molecular identification of Sarcocystis species in diaphragm muscle tissue of European mouflon (Ovis gmelini musimon) from Austria. Parasitol. Res. 2021, 120, 2695–2702. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Trakimas, G.; Juškaitis, R.; Ulevičius, A.; Balčiauskienė, L. Atlas of Lithuanian Mammals, Amphibians and Reptiles, 2nd ed.; Akstis: Vilnius, Lithuania, 1999. [Google Scholar]
- Cacciò, S.M.; De Giacomo, M.; Aulicino, F.A.; Pozio, E. Giardia Cysts in Wastewater Treatment Plants in Italy. Appl. Environ. Microbiol. 2003, 69, 3393–3398. [Google Scholar] [CrossRef] [Green Version]
- Gallas-Lindemann, C.; Sotiriadou, I.; Mahmoudi, M.R.; Karanis, P. Detection of Toxoplasma gondii oocysts in different water resources by Loop Mediated Isothermal Amplification (LAMP). Acta Trop. 2013, 125, 231–236. [Google Scholar] [CrossRef]
- Ramsay, C.N.; Wagner, A.; Robertson, C.; Smith, H.V.; Pollock, K.G. Effects of Drinking-Water Filtration on Cryptosporidium Seroepidemiology, Scotland. Emerg. Infect. Dis. 2014, 20, 70–76. [Google Scholar] [CrossRef]
- Xiao, D.; Lyu, Z.; Chen, S.; Huo, Y.; Fan, W.; Huo, M. Tracking Cryptosporidium in urban wastewater treatment plants in a cold region: Occurrence, species and infectivity. Front. Environ. Sci. Eng. 2022, 16, 112. [Google Scholar] [CrossRef]
- Castro-Hermida, J.A.; García-Presedo, I.; González-Warleta, M.; Mezo, M. Cryptosporidium and Giardia detection in water bodies of Galicia, Spain. Water Res. 2010, 44, 5887–5896. [Google Scholar] [CrossRef]
- Lass, A.; Kontogeorgos, I.; Ma, L.; Zhang, X.; Li, X.; Karanis, P. Investigation of Toxoplasma gondii in wastewater and surface water in the Qinghai-Tibet Plateau, China using real-time PCR and multilocus genotyping. Sci. Rep. 2022, 12, 5428. [Google Scholar] [CrossRef]
- Lass, A.; Szostakowska, B.; Korzeniewski, K.; Karanis, P. Detection of Giardia intestinalis in water samples collected from natural water reservoirs and wells in northern and north-eastern Poland using LAMP, real-time PCR and nested PCR. J. Water Health 2017, 15, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Zhang, Y.; Zhao, X.; Sun, L.; Hu, S. Presence and molecular characterization of Cryptosporidium and Giardia in recreational lake water in Tianjin, China: A preliminary study. Sci. Rep. 2018, 8, 2353. [Google Scholar] [CrossRef] [Green Version]
- Dreelin, E.A.; Ives, R.L.; Molloy, S.; Rose, J.B. Cryptosporidium and Giardia in Surface Water: A Case Study from Michigan, USA to Inform Management of Rural Water Systems. Int. J. Environ. Res. Public Health 2014, 11, 10480–10503. [Google Scholar] [CrossRef] [Green Version]
- Gjerde, B.; Luzón, M.; Alunda, J.M.; De La Fuente, C. Morphological and molecular characteristics of six Sarcocystis spp. from red deer (Cervus elaphus) in Spain, including Sarcocystis cervicanis and three new species. Parasitol. Res. 2017, 116, 2795–2811. [Google Scholar] [CrossRef]
- Gjerde, B.; Giacomelli, S.; Bianchi, A.; Bertoletti, I.; Mondani, H.; Gibelli, L.R. Morphological and molecular characterization of four Sarcocystis spp., including Sarcocystis linearis n. sp., from roe deer (Capreolus capreolus) in Italy. Parasitol. Res. 2017, 116, 1317–1338. [Google Scholar] [CrossRef]
- Gazzonis, A.L.; Gjerde, B.; Villa, L.; Minazzi, S.; Zanzani, S.A.; Riccaboni, P.; Sironi, G.; Manfredi, M.T. Prevalence and molecular characterisation of Sarcocystis miescheriana and Sarcocystis suihominis in wild boars (Sus scrofa) in Italy. Parasitol. Res. 2019, 118, 1271–1287. [Google Scholar] [CrossRef]
- Rudaitytė-Lukošienė, E.; Cuevas, G.E.D.D.L.; Prakas, P.; Calero-Bernal, R.; Martínez-González, M.; Strazdaitė-Žielienė, Ž.; Servienė, E.; Habela, M.A.; Butkauskas, D. Sarcocystis spp. diversity in the roe deer (Capreolus capreolus) from Lithuania and Spain. Parasitol. Res. 2020, 119, 1363–1370. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, B.M.; Dunams, D.B.; Pritt, B. Restricted genetic diversity in the ubiquitous cattle parasite, Sarcocystis cruzi. Infect. Genet. Evol. 2008, 8, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, R.; Ugorski, M.; Bednarski, M. Molecular characterization of Sarcocystis species from Polish roe deer based on ssu rRNA and cox1 sequence analysis. Parasitol. Res. 2014, 113, 3029–3039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudaitytė-Lukošienė, E.; Prakas, P.; Butkauskas, D. Molecular identification of seven Sarcocystis species in red deer (Cervus elaphus) from Lithuania. Parasitol. Int. 2021, 85, 102419. [Google Scholar] [CrossRef] [PubMed]
- Fradette, M.-S.; Culley, A.I.; Charette, S.J. Detection of Cryptosporidium spp. and Giardia spp. in Environmental Water Samples: A Journey into the Past and New Perspectives. Microorganisms 2022, 10, 1175. [Google Scholar] [CrossRef]
- Karanis, P.; Kourenti, C.; Smith, H. Waterborne transmission of protozoan parasites: A worldwide review of outbreaks and lessons learnt. J. Water Health 2007, 5, 1–38. [Google Scholar] [CrossRef]
- Galvani, A.T.; Christ, A.P.G.; Padula, J.A.; Barbosa, M.R.F.; de Araújo, R.S.; Sato, M.I.Z.; Razzolini, M.T.P. Real-time PCR detection of Toxoplasma gondii in surface water samples in São Paulo, Brazil. Parasitol. Res. 2019, 118, 631–640. [Google Scholar] [CrossRef]
- Reiling, S.J.; Merks, H.; Zhu, S.; Boone, R.; Corneau, N.; Dixon, B.R. A cloth-based hybridization array system for rapid detection of the food- and waterborne protozoan parasites Giardia duodenalis, Cryptosporidium spp. and Toxoplasma gondii. Food Waterborne Parasitol. 2021, 24, e00130. [Google Scholar] [CrossRef]
- Verma, S.K.; Lindsay, D.S.; Grigg, M.E.; Dubey, J.P. Isolation, Culture and Cryopreservation of Sarcocystiss pecies. Curr. Protoc. Microbiol. 2017, 45, 20D.1.1–20D.1.27. [Google Scholar] [CrossRef] [Green Version]
- Alonso, J.L.; Amorós, I.; Guy, R.A. Quantification of viable Giardia cysts and Cryptosporidium oocysts in wastewater using propidium monoazide quantitative real-time PCR. Parasitol. Res. 2014, 113, 2671–2678. [Google Scholar] [CrossRef]
- Rousseau, A.; La Carbona, S.; Dumètre, A.; Robertson, L.J.; Gargala, G.; Escotte-Binet, S.; Favennec, L.; Villena, I.; Gérard, C.; Aubert, D. Assessing viability and infectivity of foodborne and waterborne stages (cysts/oocysts) of Giardia duodenalis, Cryptosporidium spp., and Toxoplasma gondii: A review of methods. Parasite 2018, 25, 14. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Primers | Ta, °C | ExT, s | ProS, bp | ||||
|---|---|---|---|---|---|---|---|---|
| PCR Round | Primer Origin | Name | Orientation | Sequence (5′–3′) | ||||
| S. bovifelis | 1st | PV | V2bo1 | Forward | AACTTCCTAGGTACAGCGGTATTCG | 60 | 40 | 556 |
| V2bo2 | Reverse | TGAACAGCAGTACGAAGGCAAC | ||||||
| 2nd | V2bo3 | Forward | ATATTTACCGGTGCCGTACTTATGTT | 60 | 30 | 410 | ||
| V2bo4 | Reverse | GCCACATCATTGGTGCTTAGTCT | ||||||
| S. cruzi | 1st | PV | V2cr1 | Forward | TACAATGTGCTGTTTACGCTCCA | 57 | 50 | 776 |
| V2cr2 | Reverse | GCAATCATGATAGTTACGGCAGA | ||||||
| 2nd | V2cr3 | Forward | ACCATCCTGTTCTGTGGTGCTATG | 65 | 30 | 298 | ||
| V2cr4 | Reverse | AAACTACTTTACTGCCTACGGTACTC | ||||||
| 1st | PS | V2cr1 | Forward | TACAATGTGCTGTTTACGCTCCA | 63 | 55 | 777 | |
| V2cr2a | Reverse | CAATCATGATAGTTACGGCAGAGA | ||||||
| 2nd | V2cr3c | Forward | TCCAAGTACACGGCATTATTTACC | 59 | 30 | 268 | ||
| V2cr4 | Reverse | AAACTACTTTACTGCCTACGGTACTC | ||||||
| S. hirsuta | 1st | PV | V2hi5 | Forward | TATGTTGGTTCTGCCGAAGTCAT | 60 | 45 | 686 |
| V2hi6 | Reverse | GGTATGGCAATCATTATGGTTACAG | ||||||
| 2nd | V2hi7 | Forward | GCACCGTAATATTTCAGGGATGT | 60 | 30 | 299 | ||
| V2hi8 | Reverse | AACCTGCTTGCCGGAGTAAGTA | ||||||
| S. arieticanis | 1st | PV | V2arie1 | Forward | CTCTTTGCCGTAGATTCGCTAGTTA | 63 | 55 | 884 |
| V2arie2 | Reverse | CAAAGATCGGTAGATATCCAATGC | ||||||
| 2nd | V2arie3 | Forward | TAGTTCTTGGCCTGGCTATTCTT | 59 | 30 | 371 | ||
| V2arie4 | Reverse | CTGACCTCCAAAAACTGGCTTAC | ||||||
| S. tenella | 1st | PV | V2te1 | Forward | GAGCGGTGAACTTCTTAGGAACC | 60 | 40 | 537 |
| V2te2 | Reverse | CCCAATAATCCGCTGTTAACGTA | ||||||
| 2nd | V2te3b | Forward | ATTGTAATGCTCCTCGACGATATG | 57 | 30 | 314 | ||
| V2te4 | Reverse | ATAGTCACGGCAGAGAAGTAGGAC | ||||||
| 1st | PS | V2te1 | Forward | GAGCGGTGAACTTCTTAGGAACC | 60 | 40 | 537 | |
| V2te2 | Reverse | CCCAATAATCCGCTGTTAACGTA | ||||||
| 2nd | V2te3c | Forward | ATGTTGATCATAACCATACCGATATTC | 61 | 30 | 348 | ||
| V2te4 | Reverse | ATAGTCACGGCAGAGAAGTAGGAC | ||||||
| S. capracanis | 1st | PV | VocaF | Forward | GTAAACTTCCTGGGTACTGTGCTGT | 60 | 40 | 531 |
| VocaR1 | Reverse | CCAGTAATCCGCTGTCAAGATAC | ||||||
| 2nd | V2cap3 | Forward | ATACCGATCTTTACGGGAGCAGTA | 63 | 30 | 330 | ||
| V2cap4 | Reverse | GGTCACCGCAGAGAAGTACGAT | ||||||
| S. bertrami | 1st | PV | V2ber1 | Forward | GTATGAACTGTCAACGGATGGAGTA | 58 | 60 | 883 |
| V2ber2 | Reverse | AGAAGCCATGTTCGTGACTACC | ||||||
| 2nd | V2ber3 | Forward | GTACTACCTCCTTCCAGTCGGTTC | 57 | 40 | 600 | ||
| V2ber4 | Reverse | CGGGTATCCACTTCAAGTCCAG | ||||||
| 1st | PS | V2ber3 | Forward | GTACTACCTCCTTCCAGTCGGTTC | 58 | 45 | 605 | |
| V2ber6 | Reverse | ACGACCGGGTATCCACTTCA | ||||||
| 2nd | V2ber7 | Forward | CCCCACTCAGTACGAACTCC | 59 | 30 | 381 | ||
| V2ber8 | Reverse | ACTGCGATATAACTCCAAAACCA | ||||||
| S. miescheriana | 1st | PV | V2mie1 | Forward | TGCTGCGGTATGAACTATCTACCT | 61 | 60 | 922 |
| V2mie2 | Reverse | GCCCAGAGATCCAAATCCAG | ||||||
| 2nd | V2mie3 | Forward | CTTGGTTCAACGTTACTCCTCCA | 61 | 30 | 474 | ||
| V2mie4 | Reverse | CTTCGATCCAGCTGAACTAAAGC | ||||||
| 1st | PS | V2mie3 | Forward | CTTGGTTCAACGTTACTCCTCCA | 58 | 50 | 701 | |
| V2mie2 | Reverse | GCCCAGAGATCCAAATCCAG | ||||||
| 2nd | V2mie5 | Forward | TCCTCGGTATTAGCAGCGTACTG | 55 | 30 | 358 | ||
| V2mie6 | Reverse | ATTGAAGGGCCACCAAACAC | ||||||
| Species | Assigned No. in GenBank (Length, bp) | Position of cox1 Fragment Corresponding to S. gracilis MN339303 | Sequence Similarity, % | |
|---|---|---|---|---|
| Comparison of Acquired Sequences vs. the Same Species Accessible in GenBank | Comparison of Acquired Sequences vs. Greatly Related Species | |||
| S. bovifelis | OP681482–OP681486 (361) | 594–954 | 99.5–100 | S. bovini 93.1–94.5 |
| S. cruzi | OP681492–OP681501 (248, 218) | 493–741 a, 523–741 b | 95.4–100 | S. levinei 89.9–90.8 |
| S. hirsuta | OP681502–OP681504 (254) | 490–743 | 97.6–100 | S. buffalonis 92.4–93.2 |
| S. arieticanis | OP681467–OP681471 (325) | 430–754 | 92.6 *–99.4 | S. hircicanis 86.5–87.4 |
| S. tenella | OP681515–OP681524 (263, 296) | 607–869 c, 574–869 d | 96.3–100 | S. capracanis 91.3–93.2 |
| S. capracanis | OP681487–OP681491 (284) | 586–869 | 96.8–99.7 | S. tenella 90.4–92.9 |
| S. bertrami | OP681472–OP681481 (554, 336) | 294–847 e, 376–711 f | 96.4–99.8 | S. matsuoae 77.9–79.7 |
| S. miescheriana | OP681505–OP681514 (428, 315) | 308–739 g, 448–765 h | 92.4 **–99.4 | S. rangiferi 76.8–80.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baranauskaitė, A.; Strazdaitė-Žielienė, Ž.; Servienė, E.; Butkauskas, D.; Prakas, P. Molecular Identification of Protozoan Sarcocystis in Different Types of Water Bodies in Lithuania. Life 2023, 13, 51. https://doi.org/10.3390/life13010051
Baranauskaitė A, Strazdaitė-Žielienė Ž, Servienė E, Butkauskas D, Prakas P. Molecular Identification of Protozoan Sarcocystis in Different Types of Water Bodies in Lithuania. Life. 2023; 13(1):51. https://doi.org/10.3390/life13010051
Chicago/Turabian StyleBaranauskaitė, Agnė, Živilė Strazdaitė-Žielienė, Elena Servienė, Dalius Butkauskas, and Petras Prakas. 2023. "Molecular Identification of Protozoan Sarcocystis in Different Types of Water Bodies in Lithuania" Life 13, no. 1: 51. https://doi.org/10.3390/life13010051