
Characteristics of Aflatoxin B1 Degradation by Stenotrophomonas acidaminiphila and It’s Combination with Black Soldier Fly Larvae
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Medium
2.2. Germ-Free Intestinal BSFL Model Construction
2.3. Degradation of AFB1 in Peanut Meal by BSFL
2.4. Screening and Identification of AFB1-Degrading Bacteria from BSFL Gut
2.5. Characterization of the AFB1 Degradation Ability of Stenotrophomonas acidaminiphila Strain A2
2.6. The Influence of Stenotrophomonas acidaminiphila Strain A2 on the Growth Performance and AFB1 Degradation Capacity of BSFL
2.7. Quantification of AFB1 by HPLC
2.8. Statistical Analysis
3. Results
3.1. Comparison of AFB1 Degradation Ability of Germ-Free BSFL and Non-Sterile BSFL
3.2. Screening and Identification of AFB1-Degrading Bacteria from BSFL Gut
3.3. Characterization of the AFB1 Degradation Ability of S. acidaminiphila Strain A2
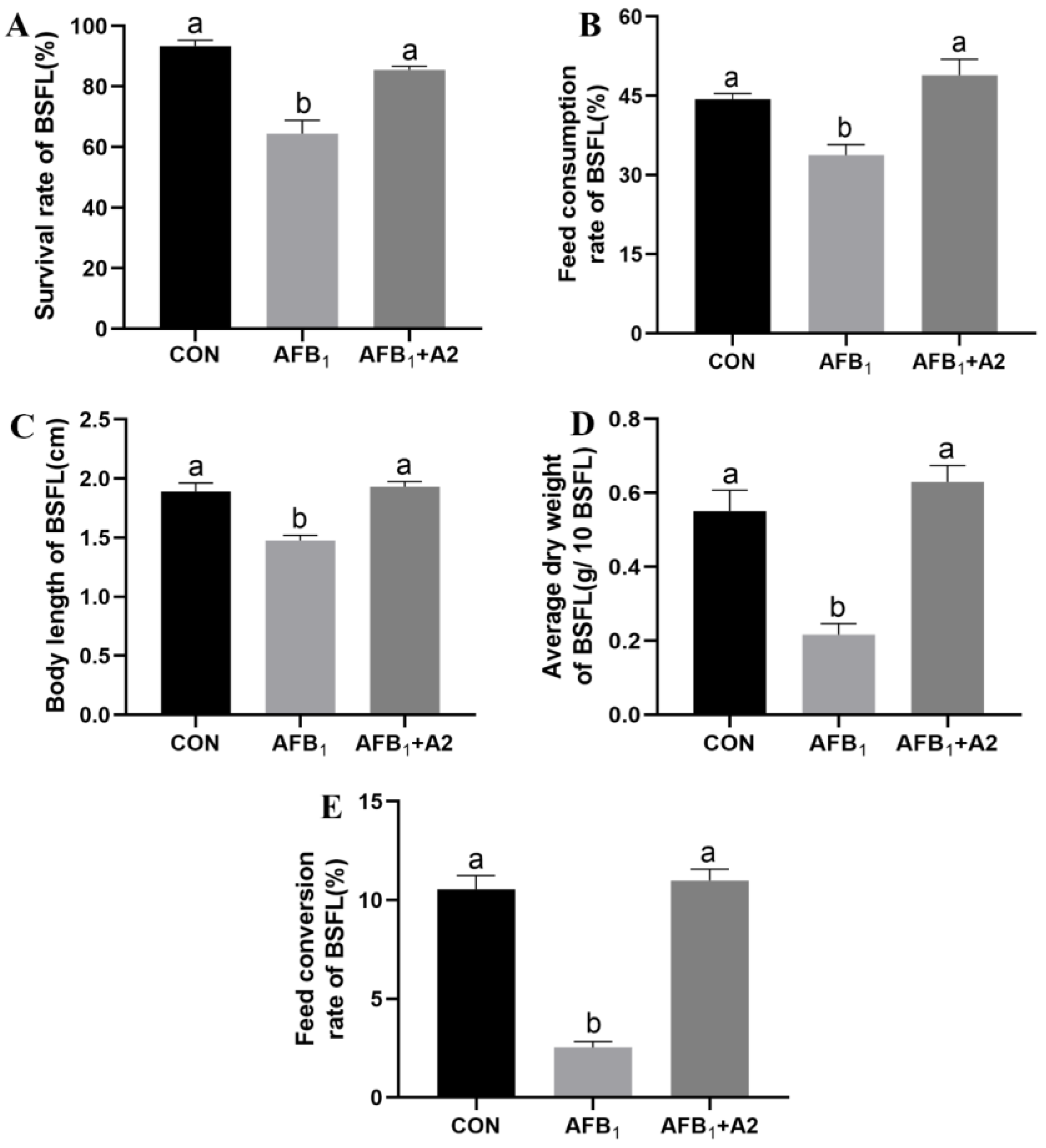
3.4. Effect of Stenotrophomonas acidaminiphila Strain A2 on the Growth Performance of BSFL
3.5. Degradation of AFB1 by BSFL Combined with Stenotrophomonas acidaminiphila Strain A2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reddy, K.R.N.; Salleh, B.; Saad, B.; Abbas, H.K.; Abel, C.A.; Shier, W.T. An overview of mycotoxin contamination in foods and its implications for human health. Toxin Rev. 2010, 29, 3–26. [Google Scholar] [CrossRef]
- My, A.; Ds, S. Dietary factors affecting aflatoxin Bi carcinogenicity. Malays. J. Nutr. 1997, 3, 161–179. [Google Scholar]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global mycotoxin occurrence in feed: A ten-year survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, M.K.; Kumar, R.; Pandey, A.K.; Soni, P.; Gangurde, S.S.; Sudini, H.K.; Fountain, J.C.; Liao, B.; Desmae, H.; Okori, P.; et al. Mitigating aflatoxin contamination in groundnut through a combination of genetic resistance and post-harvest management practices. Toxins 2019, 11, 315. [Google Scholar] [CrossRef] [Green Version]
- Bbosa, G.S.; Kitya, D.; Odda, J.; Ogwal-Okeng, J. Aflatoxins metabolism, effects on epigenetic mechanisms and their role in carcinogenesis. Health 2013, 5, 14–34. [Google Scholar] [CrossRef] [Green Version]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens-the IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, F. Global burden of aflatoxin-induced hepatocellular carcinoma: A risk assessment. Environ. Health Perspect. 2010, 118, 818–824. [Google Scholar] [CrossRef] [Green Version]
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in good, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, L.; Ma, Q.; Ji, C. Novel strategies for degradation of aflatoxins in food and feed: A review. Food Res. Int. 2021, 140, 109878. [Google Scholar] [CrossRef]
- Samuel, M.S.; Sivaramakrishna, A.; Mehta, A. Degradation and detoxification of aflatoxin B1 by Pseudomonas putida. Int. Biodeterior. Biodegrad. 2014, 86, 202–209. [Google Scholar] [CrossRef]
- Eshelli, M.; Harvey, L.; Edrada-Ebel, R.; McNeil, B. Metabolomics of the bio-degradation process of aflatoxin B1 by Actinomycetes at an initial pH of 6.0. Toxins 2015, 7, 439–456. [Google Scholar] [CrossRef] [PubMed]
- Cserháti, M.; Kriszt, B.; Krifaton, C.; Szoboszlay, S.; Háhn, J.; Tóth, S.; Nagy, I.; Kukolya, J. Mycotoxin-degradation profile of Rhodococcus strains. Int. J. Food Microbiol. 2013, 166, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Ma, Q.; Zhao, L.; Lei, Y.; Shan, Y.; Ji, C. Isolation of Bacillus subtilis: Screening for aflatoxins B1, M1, and G1 detoxification. Eur. Food Res. Technol. 2011, 232, 957–962. [Google Scholar] [CrossRef]
- Allegretti, G.; Talamini, E.; Schmidt, V.; Bogorni, P.C.; Ortega, E. Insect as feed: An emergy assessment of insect meal as a sustainable protein source for the brazilian poultry industry. J. Clean. Prod. 2018, 171, 403–412. [Google Scholar] [CrossRef]
- Salam, M.; Shahzadi, A.; Zheng, H.; Alam, F.; Nabi, G.; Dezhi, S.; Ullah, W.; Ammara, S.; Ali, N.; Bilal, M. Effect of different environmental conditions on the growth and development of black soldier fly larvae and its utilization in solid waste management and pollution mitigation. Environ. Technol. Innov. 2022, 28, 102649. [Google Scholar] [CrossRef]
- Sánchez-Muros María, J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Scala, A.; Cammack, J.A.; Salvia, R.; Scieuzo, C.; Franco, A.; Bufo, S.A.; Tomberlin, J.K.; Falabella, P. Rearing substrate impacts growth and macronutrient composition of Hermetia illucens (L.) (Diptera: Stratiomyidae) larvae produced at an industrial scale. Sci. Rep. 2020, 10, 19448. [Google Scholar] [CrossRef]
- Schiavone, A.; Dabbou, S.; De Marco, M.; Cullere, M.; Biasato, I.; Biasibetti, E.; Capucchio, M.T.; Bergagna, S.; Dezzutto, D.; Meneguz, M.; et al. Black soldier fly larva fat inclusion in finisher broiler chicken diet as an alternative fat source. Animal 2018, 12, 2032–2039. [Google Scholar] [CrossRef]
- Belghit, I.; Liland, N.S.; Waagbø, R.; Biancarosa, I.; Pelusio, N.; Li, Y.; Krogdahl, Å.; Lock, E.-J. Potential of insect-based diets for Atlantic salmon (Salmo salar). Aquaculture 2018, 491, 72–81. [Google Scholar] [CrossRef]
- Bosch, G.; Van Der Fels-Klerx, H.J.; De Rijk, T.C.; Oonincx, D.G.A.B. Aflatoxin B1 tolerance and accumulation in black soldier fly larvae (Hermetia illucens) and yellow mealworms (Tenebrio molitor). Toxins 2017, 9, 185. [Google Scholar] [CrossRef]
- Camenzuli, L.; Van Dam, R.; de Rijk, T.; Andriessen, R.; Van Schelt, J.; Van der Fels-Klerx, H.J.I. Tolerance and excretion of the mycotoxins aflatoxin B1, zearalenone, deoxynivalenol, and ochratoxin a by Alphitobius diaperinus and Hermetia illucens from contaminated substrates. Toxins 2018, 10, 91. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Zhao, S.; Chen, X.; Zhang, J.; Ni, H.; Sun, M.; Lin, H.; Liu, X.; Chen, H.; Yang, S. Bacillus Velezensis EEAM 10b strengthens nutrient metabolic process in black soldier fly larvae (Hermetia illucens) via changing gut microbiome and metabolic pathways. Front. Nutr. 2022, 9, 880488. [Google Scholar] [CrossRef] [PubMed]
- GB 5009.5-2016; The Ministry of Health of the People’s Republic of China. National Food Safety Standard: Determination of Protein in Foods. China Standards Press: Beijing, China, 2016.
- GB 5009.6-2016; The Ministry of Health of the People’s Republic of China. National Food Safety Standard: Determination of Fat in Foods. China Standards Press: Beijing, China, 2016.
- GB 5009.4-2016; The Ministry of Health of the People’s Republic of China. National Food Safety Standard: Determination of Ash in Foods. China Standards Press: Beijing, China, 2016.
- GB 5009.124-2016; The Ministry of Health of the People’s Republic of China. National Food Safety Standard: Determination of Amino Acids in Foods. China Standards Press: Beijing, China, 2016.
- Purschke, B.; Scheibelberger, R.; Axmann, S.; Adler, A.; Jäger, H. Impact of substrate contamination with mycotoxins, heavy metals and pesticides on the growth performance and composition of black soldier fly larvae (Hermetia illucens) for use in the feed and food value chain. Food Addit. Contam. A 2017, 34, 1410–1420. [Google Scholar] [CrossRef]
- Meijer, N.; Stoopen, G.; Van der Fels-Klerx, H.J.; Van Loon, J.J.A.; Carney, J.; Bosch, G. Aflatoxin B1 conversion by black soldier fly (Hermetia illucens) larval enzyme extracts. Toxins 2019, 11, 532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.; Wang, X.; Xu, X.; Cai, R.; Xie, S. Effects of heavy metals on the bioaccumulation, excretion and gut microbiome of black soldier fly larvae (Hermetia illucens). Ecotoxicol. Environ. Saf. 2020, 192, 110323. [Google Scholar] [CrossRef]
- Luo, X.; Yang, Q.; Lin, Y.; Tang, Z.; Tomberlin, J.K.; Liu, W.; Huang, Y. Black soldier fly larvae effectively degrade lincomycin from pharmaceutical industry wastes. J. Environ. Manag. 2022, 307, 114539. [Google Scholar] [CrossRef]
- Yang, C.; Ma, S.; Li, F.; Zheng, L.; Tomberlin, J.K.; Yu, Z.; Zhang, J.; Yu, C.; Fan, M.; Cai, M. Characteristics and mechanisms of ciprofloxacin degradation by black soldier fly larvae combined with associated intestinal microorganisms. Sci. Total Environ. 2022, 811, 151371. [Google Scholar] [CrossRef]
- Guan, S.; Ji, C.; Zhou, T.; Li, J.; Ma, Q.; Niu, T. Aflatoxin B1 degradation by Stenotrophomonas maltophilia and other microbes selected using coumarin medium. Int. J. Mol. Sci. 2008, 9, 1489–1503. [Google Scholar] [CrossRef] [Green Version]
- Cai, M.; Qian, Y.; Chen, N.; Ling, T.; Wang, J.; Jiang, H.; Wang, X.; Qi, K.; Zhou, Y. Detoxification of aflatoxin B1 by Stenotrophomonas sp. CW117 and characterization the thermophilic degradation process. Environ. Pollut. 2020, 261, 114178. [Google Scholar] [CrossRef]
- Zhao, L.; Guan, S.; Gao, X.; Ma, Q.; Lei, Y.; Bai, X.; Ji, C. Preparation, purification and characteristics of an aflatoxin degradation enzyme from Myxococcus fulvus ANSM068. J. Appl. Microbiol. 2011, 110, 147–155. [Google Scholar] [CrossRef]
- Guo, Y.; Qin, X.; Tang, Y.; Ma, Q.; Zhang, J.; Zhao, L. CotA laccase, a novel aflatoxin oxidase from Bacillus licheniformis, transforms aflatoxin B1 to aflatoxin Q1 and epi-aflatoxin Q1. Food Chem. 2020, 325, 126877. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zhang, X.; Chen, H.; Huang, W.; Jiang, H.; Wang, C.; Xiao, Z.; Zhang, Y.; Xu, J. Isolation and aflatoxin B1-degradation characteristics of a Microbacterium proteolyticum B204 Strain from bovine faeces. Toxins 2022, 14, 525. [Google Scholar] [CrossRef] [PubMed]
- Sangare, L.; Zhao, Y.; Folly, Y.M.E.; Chang, J.; Li, J.; Selvaraj, J.N.; Xing, F.; Zhou, L.; Wang, Y.; Liu, Y. Aflatoxin B1 degradation by a Pseudomonas strain. Toxins 2014, 6, 3028–3040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, X.; Ren, X.; Fu, J.; Wei, N.; Altomare, C.; Haidukowski, M.; Logrieco, A.F.; Zhang, Q.; Li, P. Characterization and mechanism of aflatoxin degradation by a novel strain of Trichoderma reesei CGMCC3.5218. Front. Microbiol. 2022, 13, 1003039. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Xie, H.; Gao, L.; Zhang, J.; Li, Y.; Mao, G.; Zhang, H.; Wang, F.; Lam, S.S.; Song, A. Detoxication and bioconversion of aflatoxin B1 by yellow mealworms (Tenebrio molitor): A sustainable approach for valuable larval protein production from contaminated grain. Ecotoxicol. Environ. Saf. 2022, 242, 113935. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Group 1 | SEM 2 | p-Valve | |
|---|---|---|---|---|
| GF-BSFL | NS-BSFL | |||
| Dried frass weight (g) | 8.29 a | 5.96 b | 0.09 | <0.001 |
| AFB1 concentration in frass (ng g−1) | 74.13 a | 17.24 b | 2.09 | <0.001 |
| Total residual AFB1 content in frass (ng) | 614.63 a | 101.53 b | 14.71 | <0.001 |
| AFB1 degradation rate (%) | 31.71 a | 88.72 b | 1.64 | <0.001 |
| Items | Group 1 | SEM 2 | p-Valve | ||
|---|---|---|---|---|---|
| CON | AFB1 | AFB1 + A2 | |||
| Crude fat (%) | 23.49 b | 25.47 a | 25.53 a | 0.035 | <0.001 |
| Crude fiber (%) | 8.90 | 8.90 | 9.00 | 0.019 | 0.125 |
| Crude protein (%) | 51.86 a | 51.46 ab | 51.03 b | 0.071 | 0.009 |
| Aspartic acid (%) | 3.90 | 3.94 | 3.81 | 0.029 | 0.253 |
| Glutamate (%) | 4.17 | 4.10 | 4.04 | 0.030 | 0.252 |
| Serine (%) | 1.25 a | 1.26 a | 1.20 b | 0.007 | 0.028 |
| Arginine (%) | 1.56 | 1.58 | 1.58 | 0.014 | 0.720 |
| Glycine (%) | 3.09 | 3.00 | 2.94 | 0.020 | 0.060 |
| Threonine (%) | 1.53 | 1.50 | 1.47 | 0.011 | 0.196 |
| Prolin (%) | 2.77 a | 2.65 b | 2.60 b | 0.018 | 0.025 |
| Alanine (%) | 4.25 a | 4.04 b | 3.94 b | 0.027 | 0.008 |
| Valine (%) | 2.68 | 2.61 | 2.56 | 0.018 | 0.087 |
| Methionine (%) | 0.65 | 0.62 | 0.61 | 0.007 | 0.124 |
| Cystine (%) | 0.31 b | 0.34 a | 0.30 b | 0.001 | <0.001 |
| Isoleucine (%) | 1.98 | 1.96 | 1.90 | 0.013 | 0.086 |
| Leucine (%) | 2.90 | 2.88 | 2.78 | 0.020 | 0.093 |
| Phenylalanine (%) | 1.93 | 1.93 | 1.91 | 0.016 | 0.762 |
| Histidine (%) | 1.11 | 1.12 | 1.06 | 0.012 | 0.173 |
| Lysine (%) | 2.62 | 2.70 | 2.59 | 0.015 | 0.065 |
| Tyrosine (%) | 2.50 | 2.41 | 2.41 | 0.016 | 0.116 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suo, J.; Liang, T.; Zhang, H.; Liu, K.; Li, X.; Xu, K.; Guo, J.; Luo, Q.; Yang, S. Characteristics of Aflatoxin B1 Degradation by Stenotrophomonas acidaminiphila and It’s Combination with Black Soldier Fly Larvae. Life 2023, 13, 234. https://doi.org/10.3390/life13010234
Suo J, Liang T, Zhang H, Liu K, Li X, Xu K, Guo J, Luo Q, Yang S. Characteristics of Aflatoxin B1 Degradation by Stenotrophomonas acidaminiphila and It’s Combination with Black Soldier Fly Larvae. Life. 2023; 13(1):234. https://doi.org/10.3390/life13010234
Chicago/Turabian StyleSuo, Jianghua, Tingting Liang, Haixu Zhang, Kun Liu, Xinfeng Li, Kun Xu, Jianlai Guo, Qiujiang Luo, and Sen Yang. 2023. "Characteristics of Aflatoxin B1 Degradation by Stenotrophomonas acidaminiphila and It’s Combination with Black Soldier Fly Larvae" Life 13, no. 1: 234. https://doi.org/10.3390/life13010234