

Analysis of Wine-Producing Vitis vinifera L. Biotypes, Autochthonous to Crete (Greece), Employing Ampelographic and Microsatellite Markers
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Ampelographic Classification
2.3. DNA Isolation
2.4. Determination of SSR Markers
2.5. Statistical Analysis of SSR Markers
3. Results
3.1. Ampelographic Classification
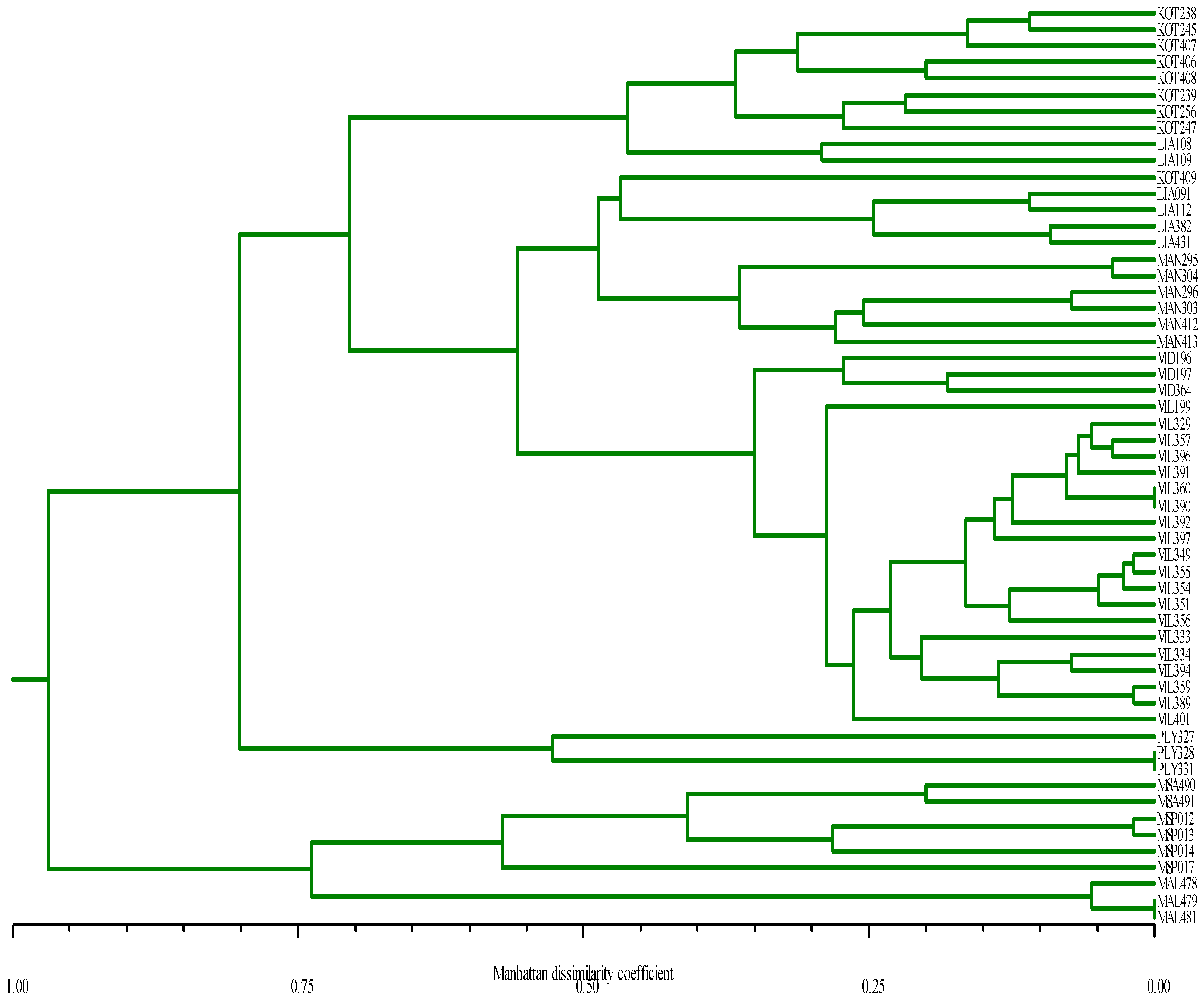
3.2. SSR Analysis
4. Discussion
4.1. In Depth Analysis of Cultivars
4.1.1. ‘Vilana’-’Vidiano’
4.1.2. ‘Plyto’
4.1.3. ‘Liatiko’
4.1.4. ‘Kotsifali’
4.1.5. ‘Mandilari’
4.1.6. ‘Moschato’
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pagnoux, C.; Bouby, L.; Ivorra, S.; Petit, C.; Valamoti, S.-M.; Pastor, T.; Picq, S.; Terral, J.-F. Inferring the agrobiodiversity of Vitis vinifera L.(grapevine) in ancient Greece by comparative shape analysis of archaeological and modern seeds. Veg. Hist. Archaeobotany 2015, 24, 75–84. [Google Scholar] [CrossRef]
- Tsivelikas, A.L.; Avramidou, E.V.; Ralli, P.E.; Ganopoulos, I.V.; Moysiadis, T.; Kapazoglou, A.; Aravanopoulos, F.A.; Doulis, A.G. Genetic diversity of Greek grapevine (Vitis vinifera L.) cultivars using ampelographic and microsatellite markers. Plant Genet. Resour. 2022, 1–13. [Google Scholar] [CrossRef]
- Bibi, A.C.; Gonias, E.D.; Doulis, A.G. Genetic diversity and structure analysis assessed by SSR markers in a large collection of Vitis cultivars from the island of Crete, Greece. Biochem. Genet. 2020, 58, 294–321. [Google Scholar] [CrossRef]
- Krimbas, V. Greek Ampelography, Vol. I. In Ministry of Agriculture, Athens, Greece; Ministry of Agriculture: Athens, Greece, 1943; Volume 83. [Google Scholar]
- Krimbas, V. Greek Ampelography, Vol. II. In Ministry of Agriculture, Athens, Greece; Ministry of Agriculture: Athens, Greece, 1944. [Google Scholar]
- Herrera, R.; Cares, V.; Wilkinson, M.; Caligari, P. Characterisation of genetic variation between Vitis vinifera cultivars from central Chile using RAPD and Inter Simple Sequence Repeat markers. Euphytica 2002, 124, 139–145. [Google Scholar] [CrossRef]
- Sabir, A.; Tangolar, S.; Buyukalaca, S.; Kafkas, S. Ampelographic and molecular diversity among grapevine (Vitis spp.) cultivars. Czech J. Genet. Plant Breed. 2009, 45, 160–168. [Google Scholar] [CrossRef] [Green Version]
- Sefc, K.M.; Regner, F.; Turetschek, E.; Glössl, J.; Steinkellner, H. Identification of microsatellite sequences in Vitis riparia and their applicability for genotyping of different Vitis species. Genome 1999, 42, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Sefc, K.; Lefort, F.; Grando, M.; Scott, K.; Steinkellner, H.; Thomas, M. Microsatellite markers for grapevine: A state of the art. In Molecular Biology & Biotechnology of the Grapevine; Springer: Dordrecht, The Netherlands, 2001; pp. 433–463. [Google Scholar]
- Bowers, J.; Dangl, G.S.; Vignani, R.; Meredith, C. Isolation and characterization of new polymorphic simple sequence repeat loci in grape (Vitis vinifera L.). Genome 1996, 39, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Merkouropoulos, G.; Michailidou, S.; Alifragkis, A.; Zioziou, E.; Koundouras, S.; Argiriou, A.; Nicolaou, N. A combined approach involving ampelographic description, berry oenological traits and molecular analysis to study native grapevine varieties of Greece. VITIS-J. Grapevine Res. 2015, 54, 99–103. [Google Scholar]
- Ekhvaia, J.; Gurushidze, M.; Blattner, F.R.; Akhalkatsi, M. Genetic diversity of Vitis vinifera in Georgia: Relationships between local cultivars and wild grapevine, V. vinifera L. subsp. sylvestris. Genet. Resour. Crop Evol. 2014, 61, 1507–1521. [Google Scholar] [CrossRef]
- Hvarlena, T.D.; Hadjinicoli, A.; Atanassov, I.I.; Ioannou, N. Genotyping Vitis vinifera L. cultivars of Cyprus by microsatellite analysis. VITIS-J. Grapevine Res. 2005, 44, 93–97. [Google Scholar]
- Giannetto, S.; Caruana, R.; La Notte, P.; Costacurta, A.; Crespan, M. A survey of Maltese grapevine germplasm using SSR markers. Am. J. Enol. Vitic. 2010, 61, 419–424. [Google Scholar] [CrossRef]
- El Oualkadi, A.; Ater, M.; Messaoudi, Z.; Laucou, V.; Boursiquot, J.-M.; Lacombe, T.; This, P. Molecular characterization of Moroccan grapevine germplasm using SSR markers for the establishment of a reference collection. OENO One 2009, 43, 135–148. [Google Scholar] [CrossRef]
- de Mattia, F.; Imazio, S.; Grassi, F.; Lovicu, G.; Tardáguila, J.; Failla, O.; Maitt, C.; Scienza, A.; Labra, M. Genetic characterization of Sardinia grapevine cultivars by SSR markers analysis. OENO One 2007, 41, 175–184. [Google Scholar] [CrossRef]
- Ghrissi, H.; De Andrés, M.T.; Andreu, L.J.; Gogorcena, Y. Genetic Diversity and Structure in a Spanish Grape Germplasm Collection Assessed by SSR Markers. Aust. J. Grape Wine Res. 2022, 2022. [Google Scholar] [CrossRef]
- Torres, R.; Aliquó, G.; Toro, A.; Fernandez, F.; Tornello, S.; Palazzo, E.; Sari, S.; Fanzone, M.; De Biazi, F.; Oviedo, H.J. Identification and recovery of local Vitis vinifera L. cultivars collected in ancient vineyards in different locations of Argentina. Aust. J. Grape Wine Res. 2022, 28, 581–589. [Google Scholar] [CrossRef]
- López, M.; Cid, N.; González, M.V.; Cuenca, B.; Prado, M.J.; Rey, M. Microsatellite and AFLP analysis of autochthonous grapevine cultivars from Galicia (Spain). Am. J. Enol. Vitic. 2009, 60, 215–222. [Google Scholar] [CrossRef]
- Stavrakaki, M.; Bouza, D.; Biniari, K. Differentiation of Greek grapevine cultivars (Vitis vinifera L.) based on the combination of ampelographic description and microsatellite markers. Genet. Resour. Crop Evol. 2020, 67, 21–40. [Google Scholar] [CrossRef]
- This, P.; Jung, A.; Boccacci, P.; Borrego, J.; Botta, R.; Costantini, L.; Crespan, M.; Dangl, G.; Eisenheld, C.; Ferreira-Monteiro, F. Development of a standard set of microsatellite reference alleles for identification of grape cultivars. Theor. Appl. Genet. 2004, 109, 1448–1458. [Google Scholar] [CrossRef]
- Emanuelli, F.; Lorenzi, S.; Grzeskowiak, L.; Catalano, V.; Stefanini, M.; Troggio, M.; Myles, S.; Martinez-Zapater, J.M.; Zyprian, E.; Moreira, F.M. Genetic diversity and population structure assessed by SSR and SNP markers in a large germplasm collection of grape. BMC Plant Biol. 2013, 13, 1–17. [Google Scholar] [CrossRef] [Green Version]
- De Michele, R.; La Bella, F.; Gristina, A.S.; Fontana, I.; Pacifico, D.; Garfi, G.; Motisi, A.; Crucitti, D.; Abbate, L.; Carimi, F. Phylogenetic relationship among wild and cultivated grapevine in Sicily: A hotspot in the middle of the Mediterranean Basin. Front. Plant Sci. 2019, 10, 1506. [Google Scholar] [CrossRef] [Green Version]
- Riaz, S.; De Lorenzis, G.; Velasco, D.; Koehmstedt, A.; Maghradze, D.; Bobokashvili, Z.; Musayev, M.; Zdunic, G.; Laucou, V.; Andrew Walker, M. Genetic diversity analysis of cultivated and wild grapevine (Vitis vinifera L.) accessions around the Mediterranean basin and Central Asia. BMC Plant Biol. 2018, 18, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefort, F.; Kyvelos, C.J.; Zervou, M.; Edwards, K.; Roubelakis-Angelakis, K. Characterization of new microsatellite loci from Vitis vinifera and their conservation in some Vitis species and hybrids. Mol. Ecol. Notes 2002, 2, 20–21. [Google Scholar] [CrossRef]
- Botta, R.; Scott, N.; Eynard, I.; Thomas, M. Evaluation of microsatellite sequence-tagged site markers for characterizing Vitis vinifera cultivars. Vitis 1995, 34, 99–102. [Google Scholar]
- Cipriani, G.; Spadotto, A.; Jurman, I.; Di Gaspero, G.; Crespan, M.; Meneghetti, S.; Frare, E.; Vignani, R.; Cresti, M.; Morgante, M. The SSR-based molecular profile of 1005 grapevine (Vitis vinifera L.) accessions uncovers new synonymy and parentages, and reveals a large admixture amongst varieties of different geographic origin. Theor. Appl. Genet. 2010, 121, 1569–1585. [Google Scholar] [CrossRef] [PubMed]
- Alba, V.; Bergamini, C.; Cardone, M.F.; Gasparro, M.; Perniola, R.; Genghi, R.; Antonacci, D. Morphological variability in leaves and molecular characterization of novel table grape candidate cultivars (Vitis vinifera L.). Mol. Biotechnol. 2014, 56, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Silvestroni, O.; Di Pietro, D.; Intrieri, C.; Vignani, R.; Filippetti, I.; Del Casino, C.; Scali, M.; Cresti, M. Detection of genetic diversity among clones of cv. Fortana (Vitis vinifera L.) by microsatellite DNA polymorphism analysis. VITIS-GEILWEILERHOF- 1997, 36, 147–150. [Google Scholar]
- Aradhya, M.K.; Dangl, G.S.; Prins, B.H.; Boursiquot, J.-M.; Walker, M.A.; Meredith, C.P.; Simon, C.J. Genetic structure and differentiation in cultivated grape, Vitis vinifera L. Genet. Res. 2003, 81, 179–192. [Google Scholar] [CrossRef] [Green Version]
- Dzhambazova, T.; Tsvetkov, I.; Atanassov, I.; Rusanov, K.; Martínez-Zapater, J.M.; Atanassov, A.; Hvarleva, T. Genetic diversity in native Bulgarian grapevine germplasm (Vitis vinifera L.) based on nuclear and chloroplast microsatellite polymorphisms. Vitis 2009, 48, 115–121. [Google Scholar]
- Grigoriou, A.; Tsaniklidis, G.; Hagidimitriou, M.; Nikoloudakis, N. The Cypriot indigenous grapevine germplasm is a multi-clonal varietal mixture. Plants 2020, 9, 1034. [Google Scholar] [CrossRef]
- Laucou, V.; Lacombe, T.; Dechesne, F.; Siret, R.; Bruno, J.-P.; Dessup, M.; Dessup, T.; Ortigosa, P.; Parra, P.; Roux, C. High throughput analysis of grape genetic diversity as a tool for germplasm collection management. Theor. Appl. Genet. 2011, 122, 1233–1245. [Google Scholar] [CrossRef]
- Martín, J.P.; Pinto-Carnide, O.; Leal, F.; del Carmen Martínez, M.; Ortiz, J.M. Determination of relationships among autochthonous grapevine varieties (Vitis vinifera L.) in the Northwest of the Iberian Peninsula by using microsatellite markers. Genet. Resour. Crop Evol. 2006, 53, 1255–1261. [Google Scholar] [CrossRef]
- Loukas, M.; Stavrakakis, M.; Krimbas, C. Inheritance of polymorphic isoenzymes in grape cultivars. J. Hered. 1983, 74, 181–183. [Google Scholar] [CrossRef]
- Stavrakaki, M.; Biniari, K. Ampelographic and genetic characterization of grapevine varieties (Vitis vinifera L.) of the ‘Mavroudia’group cultivated in Greece. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 525–531. [Google Scholar] [CrossRef] [Green Version]
- Stavrakakis, M.; Biniari, K. Genetic study of grape cultivars belonging to the muscat family by random amplified polymorphic DNA markers. Vitis 2012, 37, 119–122. [Google Scholar]
- Lefort, F.; Roubelakis-Angelakis, K.K. Genetic comparison of Greek cultivars of Vitis vinifera L. by nuclear microsatellite profiling. Am. J. Enol. Vitic. 2001, 52, 101–108. [Google Scholar] [CrossRef]
- Biniari, K.; Stavrakaki, M. Genetic study of native grapevine varieties of northern, western and central Greece with the use of ampelographic and molecular methods. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 46–53. [Google Scholar] [CrossRef]
- Mercenaro, L.; Nieddu, G.; Porceddu, A.; Pezzotti, M.; Camiolo, S. Sequence polymorphisms and structural variations among four grapevine (Vitis vinifera L.) cultivars representing Sardinian agriculture. Front. Plant Sci. 2017, 8, 1279. [Google Scholar] [CrossRef] [Green Version]
- Ghaffari, S.; Hasnaoui, N.; Zinelabidine, L.H.; Ferchichi, A.; Martínez-Zapater, J.M.; Ibáñez, J. Genetic diversity and parentage of Tunisian wild and cultivated grapevines (Vitis vinifera L.) as revealed by single nucleotide polymorphism (SNP) markers. Tree Genet. Genomes 2014, 10, 1103–1112. [Google Scholar] [CrossRef]
- González, R.; Vallès, J.; Garnatje, T. Genome Size Variation Assessment in Vitis vinifera L. Landraces in Ibiza and Formentera (Balearic Islands). Plants 2022, 11, 1892. [Google Scholar] [CrossRef]
- Hameed, U.K.A.; Abdelaziz, K.; El Sherif, N. Genetic diversity of grapevine (Vitis vinifera L.) Cultivars in al-madinah al-munawara based on molecular markers and morphological traits. Bangladesh J. Plant Taxon. 2020, 27, 113–127. [Google Scholar] [CrossRef]
- Santiago, J.; Boso, S.; Gago, P.; Alonso-Villaverde, V.; Martínez, M. Molecular and ampelographic characterisation of Vitis vinifera L. “Albariño”, “Savagnin Blanc” and “Caíño Blanco” shows that they are different cultivars. Span. J. Agric. Res. 2007, 5, 333–340. [Google Scholar] [CrossRef]
- Shecori, S.; Kher, M.M.; Tyagi, K.; Lerno, L.; Netzer, Y.; Lichter, A.; Ebeler, S.E.; Drori, E. A Field Collection of Indigenous Grapevines as a Valuable Repository for Applied Research. Plants 2022, 11, 2563. [Google Scholar] [CrossRef]
- Rohlf, F. NTSYS pc: Numerical Taxonomy System, Version 2.1; Exeter Publishing: Setauket, NY, USA, 2002. [Google Scholar]
- Bowers, J.E.; Meredith, C.P. The parentage of a classic wine grape, Cabernet Sauvignon. Nat. Genet. 1997, 16, 84–87. [Google Scholar] [CrossRef]
- Thomas, M.; Scott, N. Microsatellite repeats in grapevine reveal DNA polymorphisms when analysed as sequence-tagged sites (STSs). Theor. Appl. Genet. 1993, 86, 985–990. [Google Scholar] [CrossRef]
- Sefc, K.; Steinkellner, H.; Glössl, J.; Kampfer, S.; Regner, F. Reconstruction of a grapevine pedigree by microsatellite analysis. Theor. Appl. Genet. 1998, 97, 227–231. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising How the Computer Program Cervus Accommodates Genotyping Error Increases Success in Paternity Assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Lynch, M. The rate of morphological evolution in mammals from the standpoint of the neutral expectation. Am. Nat. 1990, 136, 727–741. [Google Scholar] [CrossRef]
- Forneck, A. Plant breeding: Clonality—A concept for stability and variability during vegetative propagation. In Progress in Botany; Springer: Berlin/Heidelberg, Germany, 2005; pp. 164–183. [Google Scholar]
- Ergül, A.; Perez-Rivera, G.; Söylemezoğlu, G.; Kazan, K.; Arroyo-Garcia, R. Genetic diversity in Anatolian wild grapes (Vitis vinifera subsp. sylvestris) estimated by SSR markers. Plant Genet. Resour. 2011, 9, 375–383. [Google Scholar] [CrossRef]
- Štajner, N.; Tomić, L.; Ivanišević, D.; Korać, N.; Cvetković-Jovanović, T.; Beleski, K.; Angelova, E.; Maraš, V.; Javornik, B. Microsatellite inferred genetic diversity and structure of Western Balkan grapevines (Vitis vinifera L.). Tree Genet. Genomes 2014, 10, 127–140. [Google Scholar] [CrossRef]
- Sefc, K.; Lopes, M.; Lefort, F.; Botta, R.; Roubelakis-Angelakis, K.; Ibanez, J.; Pejić, I.; Wagner, H.; Glössl, J.; Steinkellner, H. Microsatellite variability in grapevine cultivars from different European regions and evaluation of assignment testing to assess the geographic origin of cultivars. Theor. Appl. Genet. 2000, 100, 498–505. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cultivar | Number of Independent Genotypes Analyzed | Genotype Code |
|---|---|---|
| Noir (red berry) | ||
| ‘Kotsifali’ | 9 | 238, 239, 245, 247, 256, 406, 407, 408, 409 |
| ‘Liatiko’ | 6 | 091, 108, 109, 112, 382, 431 |
| ‘Mandilari’ | 6 | 295, 296, 303, 304, 412, 413 |
| sub-total Noir | 21 | |
| Blanc (white berry) | ||
| ‘Vidiano’ | 3 | 196, 197, 364 |
| ‘Vilana’ | 20 | 199, 329, 333, 334, 349, 351, 354, 355, 356, 357, 359, 360, 389, 390, 391, 392, 394, 396, 397, 401 |
| ‘Plyto’ | 3 | 327, 328, 331 |
| ‘Moschato Spinas’ | 4 | 012, 013, 014, 017 |
| sub-total Blanc | 30 | |
| Total autochthonous | 51 | |
| Out-group control | 5 | |
| ‘Moschato Samou’ | 2 | 490, 491 |
| ‘Moschato Alexandrias’ | 3 | 478, 479, 481 |
| Total analyzed | 56 |
| Locus | Number of Alleles | Ne | He | Ho | PI | Probability of Null Alleles |
|---|---|---|---|---|---|---|
| VVS2 | 12 | 3.175 | 0.794 | 0.981 | 0.072 | −0.127 |
| VVMD5 | 9 | 3.310 | 0.850 | 0.893 | 0.044 | −0.030 |
| VVMD7 | 10 | 2.793 | 0.763 | 0.679 | 0.082 | 0.058 |
| VVMD27 | 12 | 3.017 | 0.858 | 0.839 | 0.040 | 0.007 |
| VRZAG62 | 8 | 2.385 | 0.655 | 0.537 | 0.145 | 0.146 |
| VRZAG79 | 10 | 3.283 | 0.844 | 0.907 | 0.046 | −0.047 |
| VVZAG21 | 15 | 3.060 | 0.869 | 0.704 | 0.034 | 0.099 |
| VVZAG47 | 9 | 2.435 | 0.749 | 0.732 | 0.104 | −0.000 |
| VRZAG64 | 6 | 2.085 | 0.758 | 1.000 | 0.100 | −0.154 |
| VRZAG83 | 9 | 3.507 | 0.802 | 1.000 | 0.069 | −0.125 |
| VVUCH11 | 7 | 2.857 | 0.781 | 0.870 | 0.087 | −0.059 |
| VVUCH12 | 10 | 2.516 | 0.795 | 0.764 | 0.072 | 0.022 |
| VVUCH29 | 16 | 3.502 | 0.871 | 0.981 | 0.032 | −0.067 |
| Total | 133 | 3.389 × 10−16 | ||||
| Mean | 10.23 (allele/locus) | 2.085 | 0.799 | 0.837 |
| Source of Variation | Degrees of Freedom | Sum of Squares |
|---|---|---|
| among cultivars | 8 | 236,907 |
| within cultivars | 47 | 333,861 |
| total | 55 | 570,768 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avramidou, E.V.; Masaoutis, I.; Pitsoli, T.D.; Kapazoglou, A.; Pikraki, M.; Trantas, E.A.; Nikolantonakis, M.; Doulis, A.G. Analysis of Wine-Producing Vitis vinifera L. Biotypes, Autochthonous to Crete (Greece), Employing Ampelographic and Microsatellite Markers. Life 2023, 13, 220. https://doi.org/10.3390/life13010220
Avramidou EV, Masaoutis I, Pitsoli TD, Kapazoglou A, Pikraki M, Trantas EA, Nikolantonakis M, Doulis AG. Analysis of Wine-Producing Vitis vinifera L. Biotypes, Autochthonous to Crete (Greece), Employing Ampelographic and Microsatellite Markers. Life. 2023; 13(1):220. https://doi.org/10.3390/life13010220
Chicago/Turabian StyleAvramidou, Evangelia V., Ioannis Masaoutis, Theodora D. Pitsoli, Aliki Kapazoglou, Maria Pikraki, Emmanouil A. Trantas, Michael Nikolantonakis, and Andreas G. Doulis. 2023. "Analysis of Wine-Producing Vitis vinifera L. Biotypes, Autochthonous to Crete (Greece), Employing Ampelographic and Microsatellite Markers" Life 13, no. 1: 220. https://doi.org/10.3390/life13010220