
ALS-L1023 from Melissa officinalis Alleviates Liver Fibrosis in a Non-Alcoholic Fatty Liver Disease Model
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of ALS-L1023
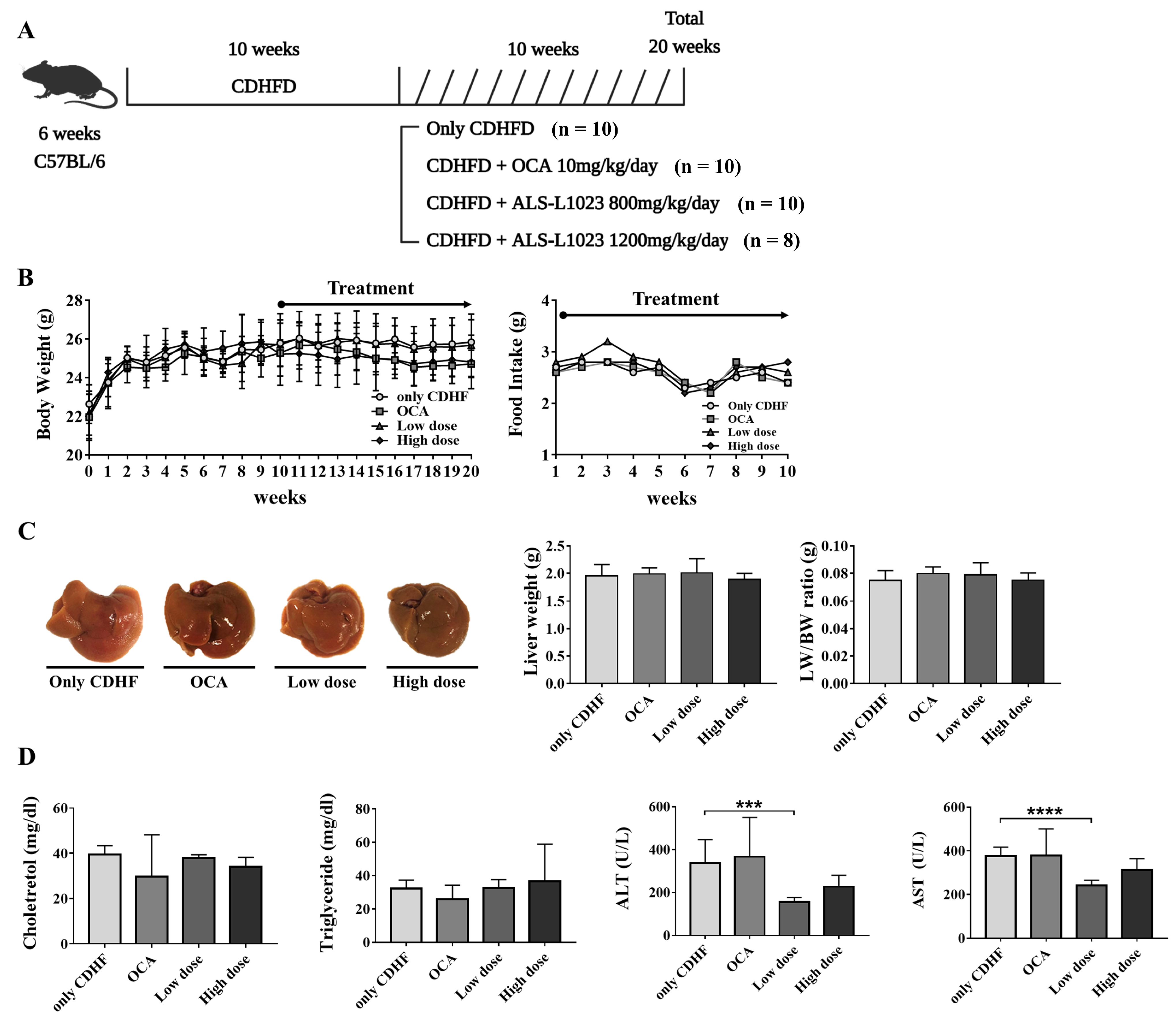
2.2. Animal Studies
2.3. Biochemical Analysis
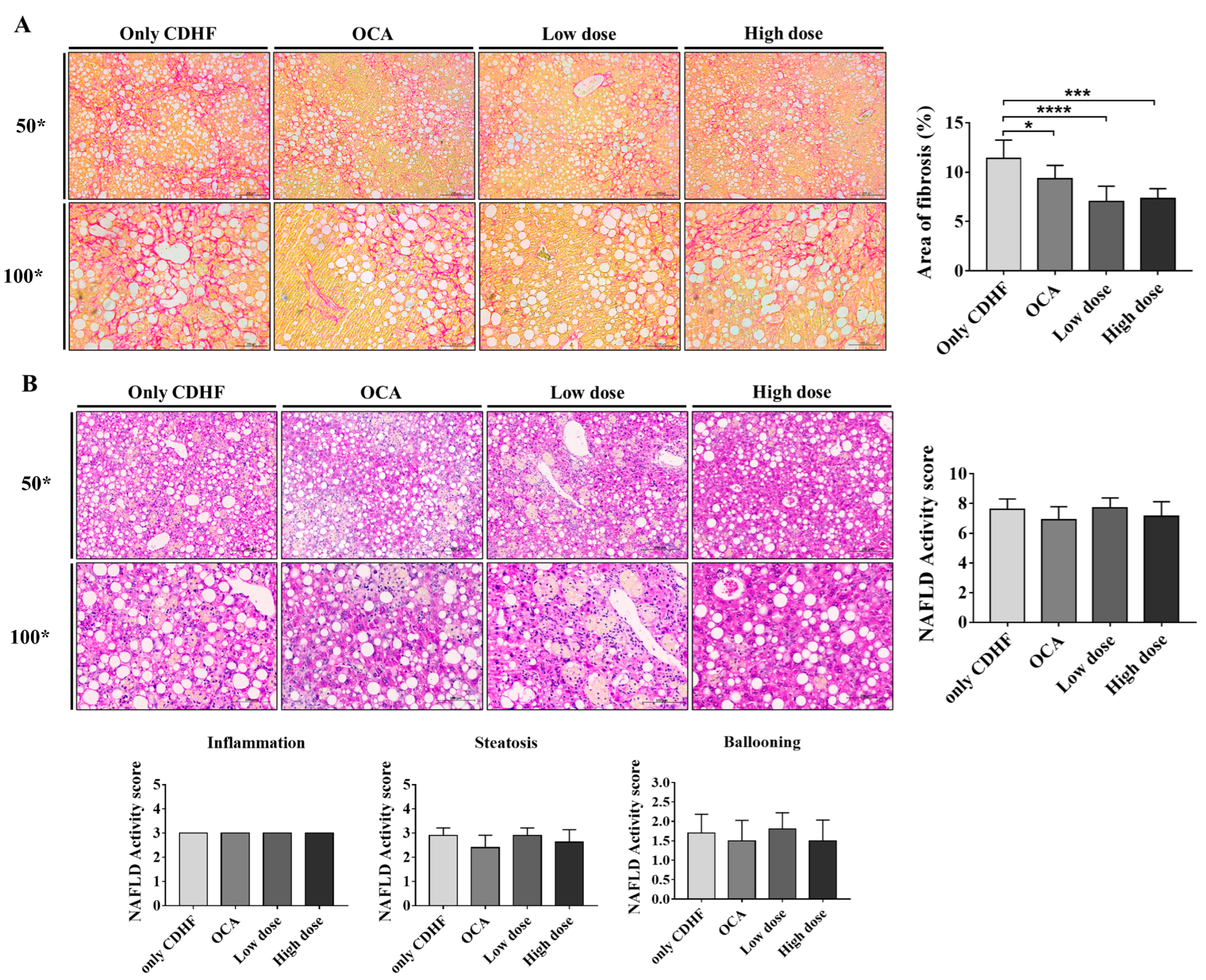
2.4. Histological Analysis
2.5. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.6. RNA Isolation for RNA Sequencing
2.7. Library Preparation and RNA Sequencing
2.8. RNA Sequencing Data Analysis
2.9. Western Blots
2.10. Statistical Analysis
3. Results
3.1. ALS-L1023 Decreased ALT and AST in the CDHFD Model
3.2. ALS-L1023 Decreased the Area of Fibrosis in the CDHFD Model
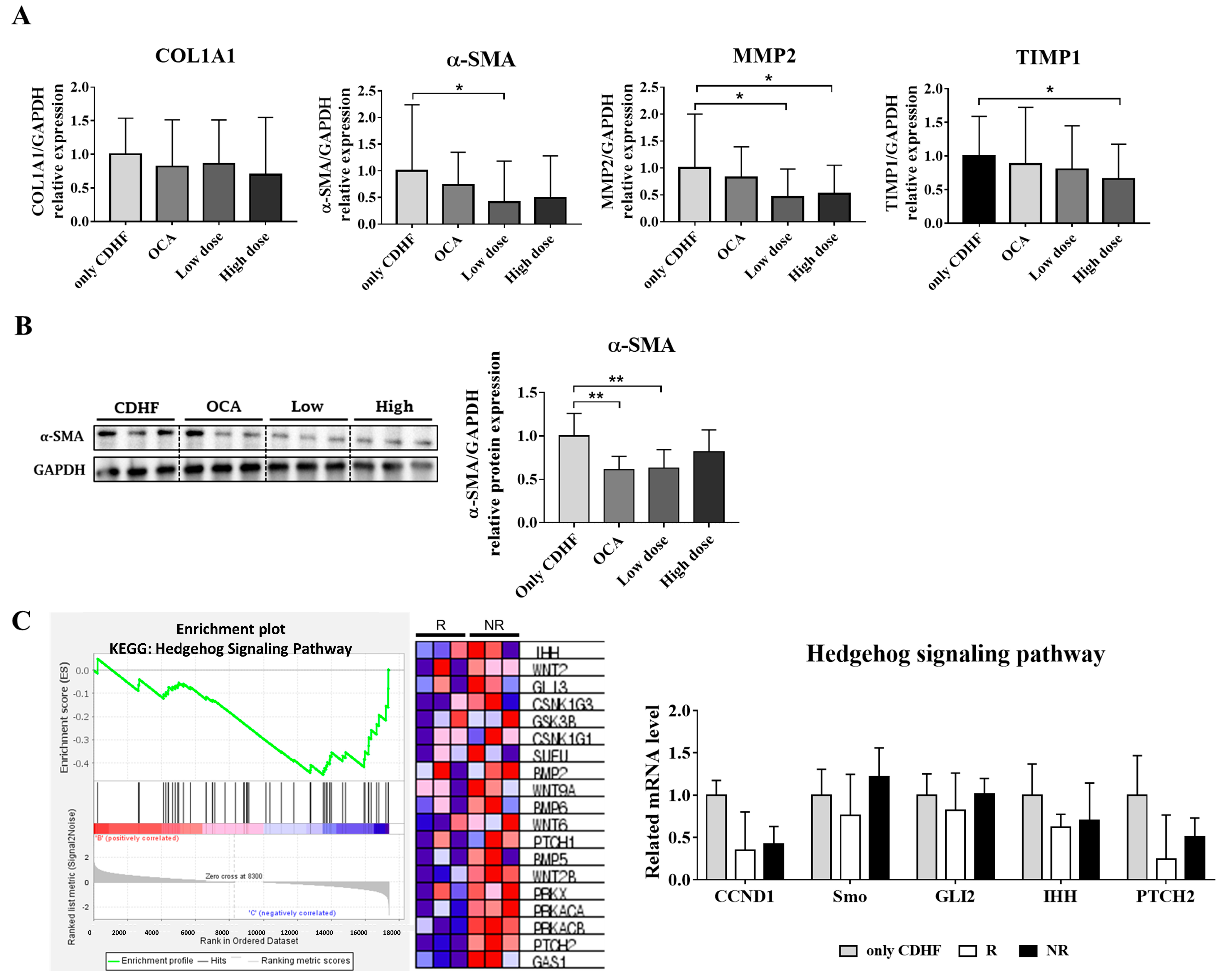
3.3. ALS-L1023 Decreased the Expression of Fibrogenic Markers
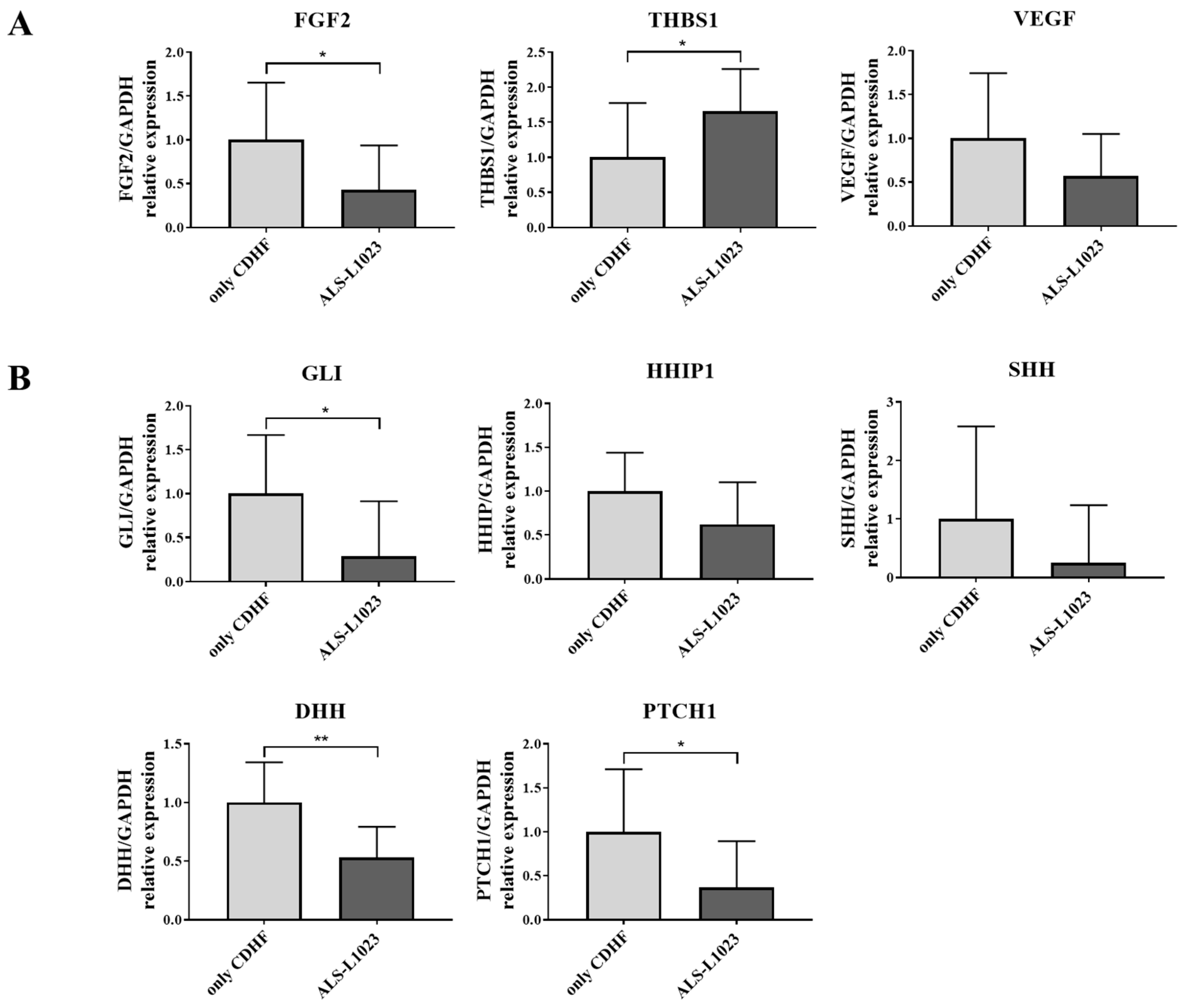
3.4. ALS-L1023 May Exert Anti-Fibrotic Effects via the Hedgehog-Signaling Pathway
3.5. ALS-L1023 Decreased the Expression of Hedgehog-Signaling Markers along with Anti-Angiogenic Effect
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.Q.; El-Serag, H.B.; Loomba, R. Global epidemiology of NAFLD-related HCC: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Kisseleva, T.; Brenner, D. Molecular and cellular mechanisms of liver fibrosis and its regression. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Sanchez, N.; Valencia-Rodriguez, A.; Coronel-Castillo, C.; Vera-Barajas, A.; Contreras-Carmona, J.; Ponciano-Rodriguez, G.; Zamora-Valdes, D. The cellular pathways of liver fibrosis in non-alcoholic steatohepatitis. Ann. Trans. Med. 2020, 8, 400. [Google Scholar] [CrossRef]
- Tacke, F.; Weiskirchen, R. Non-alcoholic fatty liver disease (NAFLD)/non-alcoholic steatohepatitis (NASH)-related liver fibrosis: Mechanisms, treatment and prevention. Ann. Trans. Med. 2021, 9, 729. [Google Scholar] [CrossRef]
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Lei, L.; Ei Mourabit, H.; Housset, C.; Cadoret, A.; Lemoinne, S. Role of Angiogenesis in the Pathogenesis of NAFLD. J. Clin. Med. 2021, 10, 1338. [Google Scholar] [CrossRef]
- Coulon, S.; Heindryckx, F.; Geerts, A.; Van Steenkiste, C.; Colle, I.; Van Vlierberghe, H. Angiogenesis in chronic liver disease and its complications. Liver Int. 2011, 31, 146–162. [Google Scholar] [CrossRef]
- Zadorozhna, M.; Di Gioia, S.; Conese, M.; Mangieri, D. Neovascularization is a key feature of liver fibrosis progression: Anti-angiogenesis as an innovative way of liver fibrosis treatment. Mol. Biol. Rep. 2020, 47, 2279–2288. [Google Scholar] [CrossRef]
- Lemoinne, S.; Thabut, D.; Housset, C. Portal myofibroblasts connect angiogenesis and fibrosis in liver. Cell Tissue Res. 2016, 365, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Poisson, J.; Lemoinne, S.; Boulanger, C.; Durand, F.; Moreau, R.; Valla, D.; Rautou, P.E. Liver sinusoidal endothelial cells: Physiology and role in liver diseases. J. Hepatol. 2017, 66, 212–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, B.Y.; Lee, H.; Woo, S.; Yoon, M.; Kim, J.; Hong, Y.; Lee, H.S.; Park, E.K.; Hahm, J.C.; Kim, J.W.; et al. Reduction of Adipose Tissue Mass by the Angiogenesis Inhibitor ALS-L1023 from Melissa officinalis. PLoS ONE 2015, 10, e0141612. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Park, B.Y.; Lee, H.S.; Park, E.K.; Hahm, J.C.; Lee, J.; Hong, Y.; Choi, S.; Park, D.; Lee, H.; et al. The anti-angiogenic herbal composition Ob-X inhibits adipose tissue growth in obese mice. Int. J. Obes. 2010, 34, 820–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Lee, H.; Lim, J.; Oh, J.; Shin, S.S.; Yoon, M. The Angiogenesis Inhibitor ALS-L1023 from Lemon-Balm Leaves Attenuates High-Fat Diet-Induced Nonalcoholic Fatty Liver Disease through Regulating the Visceral Adipose-Tissue Function. Int. J. Mol. Sci. 2017, 18, 846. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, H.; Lim, J.; Lee, H.; Yoon, S.; Shin, S.S.; Yoon, M. The lemon balm extract ALS-L1023 inhibits obesity and nonalcoholic fatty liver disease in female ovariectomized mice. Food Chem. Toxicol. 2017, 106, 292–305. [Google Scholar] [CrossRef]
- Bocca, C.; Novo, E.; Miglietta, A.; Parola, M. Angiogenesis and Fibrogenesis in Chronic Liver Diseases. Cell Mol. Gastroenterol. Hepatol. 2015, 1, 477–488. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.; Yoon, M.; Kim, J.; Hong, Y.; Kim, M.Y.; Shin, S.S.; Yoon, M. The anti-angiogenic herbal extract from Melissa officinalis inhibits adipogenesis in 3T3-L1 adipocytes and suppresses adipocyte hypertrophy in high fat diet-induced obese C57BL/6J mice. J. Ethnopharmacol. 2016, 178, 238–250. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Bioinformatics, Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- Lab, H. FASTX Toolkit; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 2014. [Google Scholar]
- Bushnell, B. BBMap: A Fast, Accurate, Splice-Aware Aligner; Lawrence Berkeley National Lab. (LBNL): Berkeley, CA, USA, 2014. [Google Scholar]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq expression estimates by correcting for fragment bias. Genome Biol. 2011, 12, R22. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, J.; Zhang, D.; Zhang, J.; Ma, J.; Jiang, H. New insights into the antifibrotic effects of sorafenib on hepatic stellate cells and liver fibrosis. J. Hepatol. 2010, 53, 132–144. [Google Scholar] [CrossRef]
- Hong, F.; Chou, H.; Fiel, M.I.; Friedman, S.L. Antifibrotic activity of sorafenib in experimental hepatic fibrosis: Refinement of inhibitory targets, dosing, and window of efficacy in vivo. Dig. Dis. Sci. 2013, 58, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Stefano, J.T.; Pereira, I.V.; Torres, M.M.; Bida, P.M.; Coelho, A.M.; Xerfan, M.P.; Cogliati, B.; Barbeiro, D.F.; Mazo, D.F.; Kubrusly, M.S.; et al. Sorafenib prevents liver fibrosis in a non-alcoholic steatohepatitis (NASH) rodent model. Braz. J. Med. Biol. Res. 2015, 48, 408–414. [Google Scholar] [CrossRef] [Green Version]
- Qu, K.; Huang, Z.; Lin, T.; Liu, S.; Chang, H.; Yan, Z.; Zhang, H.; Liu, C. New Insight into the Anti-liver Fibrosis Effect of Multitargeted Tyrosine Kinase Inhibitors: From Molecular Target to Clinical Trials. Front. Pharmacol. 2015, 6, 300. [Google Scholar] [CrossRef] [Green Version]
- Raoul, J.L.; Gilabert, M.; Adhoute, X.; Edeline, J. An in-depth review of chemical angiogenesis inhibitors for treating hepatocellular carcinoma. Expert Opin. Pharm. 2017, 18, 1467–1476. [Google Scholar] [CrossRef]
- Nakamura, I.; Zakharia, K.; Banini, B.A.; Mikhail, D.S.; Kim, T.H.; Yang, J.D.; Moser, C.D.; Shaleh, H.M.; Thornburgh, S.R.; Walters, I.; et al. Brivanib attenuates hepatic fibrosis in vivo and stellate cell activation in vitro by inhibition of FGF, VEGF and PDGF signaling. PLoS ONE 2014, 9, e92273. [Google Scholar] [CrossRef] [Green Version]
- Lefere, S.; Van de Velde, F.; Hoorens, A.; Raevens, S.; Van Campenhout, S.; Vandierendonck, A.; Neyt, S.; Vandeghinste, B.; Vanhove, C.; Debbaut, C.; et al. Angiopoietin-2 Promotes Pathological Angiogenesis and Is a Therapeutic Target in Murine Nonalcoholic Fatty Liver Disease. Hepatology 2019, 69, 1087–1104. [Google Scholar] [CrossRef]
- Oliner, J.; Min, H.; Leal, J.; Yu, D.; Rao, S.; You, E.; Tang, X.; Kim, H.; Meyer, S.; Han, S.J.; et al. Suppression of angiogenesis and tumor growth by selective inhibition of angiopoietin-2. Cancer Cell 2004, 6, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, R.; Yamamoto, H.; Ngan, C.Y.; Ohtsuka, M.; Kitani, K.; Uemura, M.; Nishimura, J.; Takemasa, I.; Mizushima, T.; Sekimoto, M.; et al. Inhibition of angiopoietin 2 attenuates lumen formation of tumour-associated vessels in vivo. Int. J. Oncol. 2013, 43, 1447–1455. [Google Scholar] [CrossRef] [Green Version]
- Orlandi, P.; Solini, A.; Banchi, M.; Brunetto, M.R.; Cioni, D.; Ghiadoni, L.; Bocci, G. Antiangiogenic Drugs in NASH: Evidence of a Possible New Therapeutic Approach. Pharmaceuticals 2021, 14, 995. [Google Scholar] [CrossRef]
- Dhar, D.; Baglieri, J.; Kisseleva, T.; Brenner, D.A. Mechanisms of liver fibrosis and its role in liver cancer. Exp. Biol. Med. 2020, 245, 96–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Shin, Y.; Jang, J.; Park, Y.; Ahn, J.; Jeong, S.; Shin, S.S.; Yoon, M. The herbal extract ALS-L1023 from Melissa officinalis alleviates visceral obesity and insulin resistance in obese female C57BL/6J mice. J. Ethnopharmacol. 2020, 253, 112646. [Google Scholar] [CrossRef] [PubMed]
- Teglund, S.; Toftgard, R. Hedgehog beyond medulloblastoma and basal cell carcinoma. Biochim. Biophys. Acta 2010, 1805, 181–208. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.S.; Omenetti, A.; Syn, W.K.; Diehl, A.M. The role of Hedgehog signaling in fibrogenic liver repair. Int. J. Biochem. Cell Biol. 2011, 43, 238–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fattahi, S.; Nikbakhsh, N.; Ranaei, M.; Sabour, D.; Akhavan-Niaki, H. Association of sonic hedgehog signaling pathway genes IHH, BOC, RAB23a and MIR195-5p, MIR509-3-5p, MIR6738-3p with gastric cancer stage. Sci. Rep. 2021, 11, 7471. [Google Scholar] [CrossRef]
- Corbit, K.C.; Aanstad, P.; Singla, V.; Norman, A.R.; Stainier, D.Y.; Reiter, J.F. Vertebrate Smoothened functions at the primary cilium. Nature 2005, 437, 1018–1021. [Google Scholar] [CrossRef]
- Rohatgi, R.; Milenkovic, L.; Scott, M.P. Patched1 regulates hedgehog signaling at the primary cilium. Science 2007, 317, 372–376. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kato, M.; Beachy, P.A. Gli2 trafficking links Hedgehog-dependent activation of Smoothened in the primary cilium to transcriptional activation in the nucleus. Proc. Natl. Acad. Sci. USA 2009, 106, 21666–21671. [Google Scholar] [CrossRef] [Green Version]
- Chong, Y.C.; Lim, T.E.; Fu, Y.; Shin, E.M.; Tergaonkar, V.; Han, W. Indian Hedgehog links obesity to development of hepatocellular carcinoma. Oncogene 2019, 38, 2206–2222. [Google Scholar] [CrossRef]
- Swiderska-Syn, M.; Syn, W.K.; Xie, G.; Kruger, L.; Machado, M.V.; Karaca, G.; Michelotti, G.A.; Choi, S.S.; Premont, R.T.; Diehl, A.M. Myofibroblastic cells function as progenitors to regenerate murine livers after partial hepatectomy. Gut 2014, 63, 1333–1344. [Google Scholar] [CrossRef]
- Choi, S.S.; Omenetti, A.; Witek, R.P.; Moylan, C.A.; Syn, W.K.; Jung, Y.; Yang, L.; Sudan, D.L.; Sicklick, J.K.; Michelotti, G.A.; et al. Hedgehog pathway activation and epithelial-to-mesenchymal transitions during myofibroblastic transformation of rat hepatic cells in culture and cirrhosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G1093–G1106. [Google Scholar] [CrossRef] [Green Version]
- Sicklick, J.K.; Li, Y.X.; Choi, S.S.; Qi, Y.; Chen, W.; Bustamante, M.; Huang, J.; Zdanowicz, M.; Camp, T.; Torbenson, M.S.; et al. Role for hedgehog signaling in hepatic stellate cell activation and viability. Lab. Invest. 2005, 85, 1368–1380. [Google Scholar] [CrossRef] [Green Version]
- Pellicciari, R.; Fiorucci, S.; Camaioni, E.; Clerici, C.; Costantino, G.; Maloney, P.R.; Morelli, A.; Parks, D.J.; Willson, T.M. 6alpha-ethyl-chenodeoxycholic acid (6-ECDCA), a potent and selective FXR agonist endowed with anticholestatic activity. J. Med. Chem. 2002, 45, 3569–3572. [Google Scholar] [CrossRef]
- Jhaveri, M.A.; Kowdley, K.V. New developments in the treatment of primary biliary cholangitis—Role of obeticholic acid. Ther. Clin. Risk Manag. 2017, 13, 1053–1060. [Google Scholar] [CrossRef] [Green Version]
- Abenavoli, L.; Falalyeyeva, T.; Boccuto, L.; Tsyryuk, O.; Kobyliak, N. Obeticholic Acid: A New Era in the Treatment of Nonalcoholic Fatty Liver Disease. Pharmaceuticals 2018, 11, 104. [Google Scholar] [CrossRef] [Green Version]
- Shah, R.A.; Kowdley, K.V. Obeticholic acid for the treatment of nonalcoholic steatohepatitis. Exp. Rev. Gastroenterol. Hepatol. 2020, 14, 311–321. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Ratziu, V.; Loomba, R.; Rinella, M.; Anstee, Q.M.; Goodman, Z.; Bedossa, P.; Geier, A.; Beckebaum, S.; Newsome, P.N.; et al. Obeticholic acid for the treatment of non-alcoholic steatohepatitis: Interim analysis from a multicentre, randomised, placebo-controlled phase 3 trial. Lancet 2019, 394, 2184–2196. [Google Scholar] [CrossRef] [Green Version]
- Feigh, M.; Tølbøl, K.S.; Kristiansen, M.B.; Veidal, S.S.; Fensholdt, L.; Vrang, N. Comparative effects of liraglutide, elafibranor, and obeticholic acid on NAFLD activity score and fibrosis stage in a diet-induced obese mouse model of biopsy-confirmed NASH. Hepatology 2017, 66, S599. [Google Scholar] [CrossRef]
- Haczeyni, F.; Poekes, L.; Wang, H.; Mridha, A.R.; Barn, V.; Geoffrey Haigh, W.; Ioannou, G.N.; Yeh, M.M.; Leclercq, I.A.; Teoh, N.C. Obeticholic acid improves adipose morphometry and inflammation and reduces steatosis in dietary but not metabolic obesity in mice. Obesity 2017, 25, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Kong, B.; Luyendyk, J.P.; Tawfik, O.; Guo, G.L. Farnesoid X receptor deficiency induces nonalcoholic steatohepatitis in low-density lipoprotein receptor-knockout mice fed a high-fat diet. J. Pharmacol. Exp. Ther. 2009, 328, 116–122. [Google Scholar] [CrossRef]
- Morrison, M.C.; Verschuren, L.; Salic, K.; Verheij, J.; Menke, A.; Wielinga, P.Y.; Iruarrizaga-Lejarreta, M.; Gole, L.; Yu, W.M.; Turner, S.; et al. Obeticholic Acid Modulates Serum Metabolites and Gene Signatures Characteristic of Human NASH and Attenuates Inflammation and Fibrosis Progression in Ldlr-/-.Leiden Mice. Hepatol. Commun. 2018, 2, 1513–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolbol, K.S.; Kristiansen, M.N.; Hansen, H.H.; Veidal, S.S.; Rigbolt, K.T.; Gillum, M.P.; Jelsing, J.; Vrang, N.; Feigh, M. Metabolic and hepatic effects of liraglutide, obeticholic acid and elafibranor in diet-induced obese mouse models of biopsy-confirmed nonalcoholic steatohepatitis. World J. Gastroenterol. 2018, 24, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Jackson, C.V.; Zimmerman, K.; Zhang, L.K.; Finnearty, C.M.; Sandusky, G.E.; Zhang, G.; Peterson, R.G.; Wang, Y.J. The FATZO mouse, a next generation model of type 2 diabetes, develops NAFLD and NASH when fed a Western diet supplemented with fructose. BMC Gastroenterol. 2019, 19, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbeke, L.; Mannaerts, I.; Schierwagen, R.; Govaere, O.; Klein, S.; Vander Elst, I.; Windmolders, P.; Farre, R.; Wenes, M.; Mazzone, M.; et al. FXR agonist obeticholic acid reduces hepatic inflammation and fibrosis in a rat model of toxic cirrhosis. Sci. Rep. 2016, 6, 33453. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Huang, N.; Guo, Y.; Cui, S.; Ge, C.; He, Q.; Pan, X.; Wang, G.; Wang, H.; Hao, H. Combined obeticholic acid and apoptosis inhibitor treatment alleviates liver fibrosis. Acta Pharm. Sin. B 2019, 9, 526–536. [Google Scholar] [CrossRef]
- Schwabl, P.; Hambruch, E.; Seeland, B.A.; Hayden, H.; Wagner, M.; Garnys, L.; Strobel, B.; Schubert, T.L.; Riedl, F.; Mitteregger, D.; et al. The FXR agonist PX20606 ameliorates portal hypertension by targeting vascular remodelling and sinusoidal dysfunction. J. Hepatol. 2017, 66, 724–733. [Google Scholar] [CrossRef] [Green Version]
- Ferrigno, A.; Di Pasqua, L.G.; Berardo, C.; Siciliano, V.; Rizzo, V.; Adorini, L.; Richelmi, P.; Vairetti, M. The farnesoid X receptor agonist obeticholic acid upregulates biliary excretion of asymmetric dimethylarginine via MATE-1 during hepatic ischemia/reperfusion injury. PLoS ONE 2018, 13, e0191430. [Google Scholar] [CrossRef]
- Garbuzenko, D.V.; Arefyev, N.O.; Belov, D.V. Mechanisms of adaptation of the hepatic vasculature to the deteriorating conditions of blood circulation in liver cirrhosis. World J. Hepatol. 2016, 8, 665–672. [Google Scholar] [CrossRef]
- Garbuzenko, D.V.; Arefyev, N.O.; Kazachkov, E.L. Antiangiogenic therapy for portal hypertension in liver cirrhosis: Current progress and perspectives. World J. Gastroenterol. 2018, 24, 3738–3748. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| Mouse GAPDH | F: 5′-GTT GTC TCC TGC GAC TTC-3′ |
| R: 5′-GGT GGTCCA GGG TTT CTT-3′ | |
| Mouse COL1A1 | F: 5′-CTG GAC AAC GTG GTG TGG-3′ |
| R: 5′-TGT TTG CCA GGT TCA CCA-3′ | |
| Mouse α-SMA | F: 5′-TCG GAT ACT TCA GCG TCA GGA-3′ |
| R: 5′-GTC CCA GAC ATC AGG GAG TAA-3′ | |
| Mouse MMP2 | F: 5′-GCT GTA TTC CCG ACC GTT GA-3′ |
| R: 5′-TGG TCC GCG TAA AGT ATG GG-3′ | |
| Mouse TIMP1 | F: 5′-ATC TCT GGC ATC TGG CAT CC-3′ |
| R: 5′-TTG CAG AAG GCT GTC TGT GG-3′ | |
| Mouse CCND1 | F: 5′-CGC CCT CCG TAT CTT ACT TCA-3′ |
| R: 5′-TCT TCG CAC TTC TGC TCC TC-3′ | |
| Mouse Smo | F: 5′-GCG GGT TTG GCT TTT GAC-3′ |
| R: 5′-GGA CAC ATC CTG GGG CAA TA-3′ | |
| Mouse GLI1 | F: 5′-GCC TTG AAA ACC TCA AGA CG-3′ |
| R: 5′-ATG GCT TCT CAT TGG AGT GG-3′ | |
| Mouse GLI2 | F: 5′-AGT GGA ATG AGG TGA GTT CTG G-3′ |
| R: 5′-AGG CTG GCT TCT GTT GGA C-3′ | |
| Mouse HHIP1 | F: 5′-GAA GGA GAT GCG AAG TTT GG-3′ |
| R: 5′-CCC TTC TCT TTA GGC GCT TT-3′ | |
| Mouse IHH | F: 5′-TCA AGG ACG AGG AGA ACA CG-3′ |
| R: 5′-ACC CGC AGT TTC ACA CCA-3′ | |
| Mouse SHH | F: 5′-CTG GTG ATC CTT GCT TCC TC-3′ |
| R: 5′-GGC TAA AGG GGT CAG CTT TT-3′ | |
| Mouse DHH | F: 5′-CAA GCA GTT TGT GCC CAG TA-3′ |
| R: 5′-GTC GGG GTT GTA GTT GGG TA-3′ | |
| Mouse PTCH1 | F: 5′-GGA GCT CAG GCA ATA CGA AG-3′ |
| R: 5′-GGA GGC TGA TGT CTG GAG TC-3′ | |
| Mouse PTCH2 | F: 5′-GCC TGC GTA ACA ATG CTG-3′ |
| R: 5′- GCA CAA AGC CCA AGA CCT -3′ | |
| Mouse VEGF | F: 5′-GTA CCT CCA CCA TGC CAA GT-3′ |
| R: 5′-TCA CAT CTG CAA GTA CGT TCG-3′ | |
| Mouse FGF2 | F: 5′-AGC GGC TCT ACT GCA AGA AC-3′ |
| R: 5′-GCC GTC CAT CTT CCT TCA TA-3′ | |
| Mouse THBS1 | F: 5′-CCA AAG CCT GCA AGA AAG AC-3′ |
| R: 5′-CCT GCT TGT TGC AAA CTT GA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, E.J.; Kim, Y.; Kim, J.E.; Yoon, E.L.; Lee, S.R.; Jun, D.W. ALS-L1023 from Melissa officinalis Alleviates Liver Fibrosis in a Non-Alcoholic Fatty Liver Disease Model. Life 2023, 13, 100. https://doi.org/10.3390/life13010100
Lee EJ, Kim Y, Kim JE, Yoon EL, Lee SR, Jun DW. ALS-L1023 from Melissa officinalis Alleviates Liver Fibrosis in a Non-Alcoholic Fatty Liver Disease Model. Life. 2023; 13(1):100. https://doi.org/10.3390/life13010100
Chicago/Turabian StyleLee, Eun Jeoung, Yun Kim, Ji Eun Kim, Eileen Laurel Yoon, Sung Ryol Lee, and Dae Won Jun. 2023. "ALS-L1023 from Melissa officinalis Alleviates Liver Fibrosis in a Non-Alcoholic Fatty Liver Disease Model" Life 13, no. 1: 100. https://doi.org/10.3390/life13010100