
Heat Shock Proteins and Antioxidant Genes Involved in Heat Combined with Drought Stress Responses in Perennial Rye Grass

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Cultivation and Treatment
2.2. Determination of Hydrogen Peroxide Accumulation
2.3. Measurement of Malondialdehyde Content
2.4. RNA Extraction and Real-Time PCR Analysis
2.5. Interactome and Gene Expression Partners Analysis
2.6. Statistical Analysis
3. Results
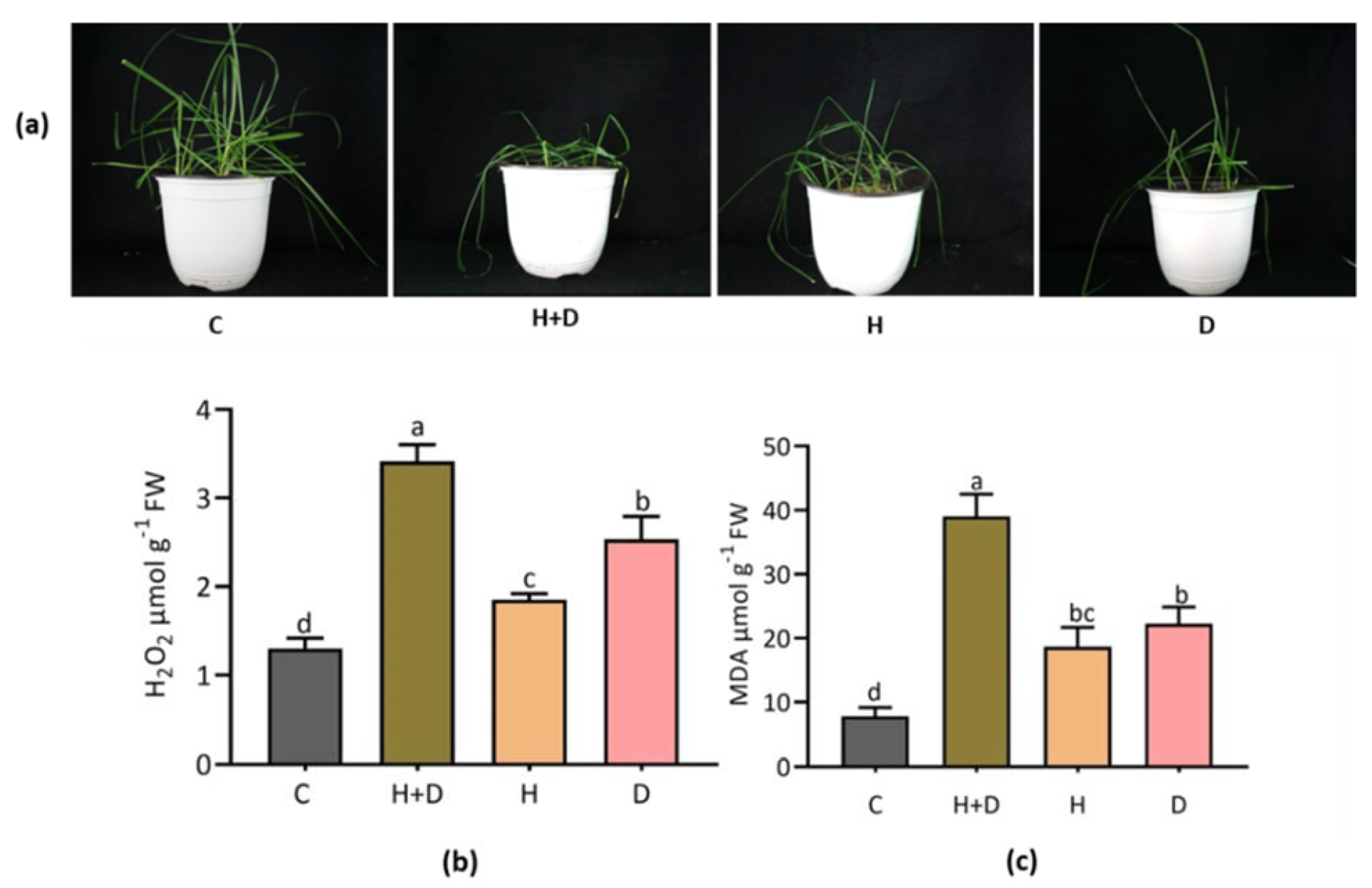
3.1. The Effect of Heat and Drought Stress on Plant Morphology and Membrane Oxidation
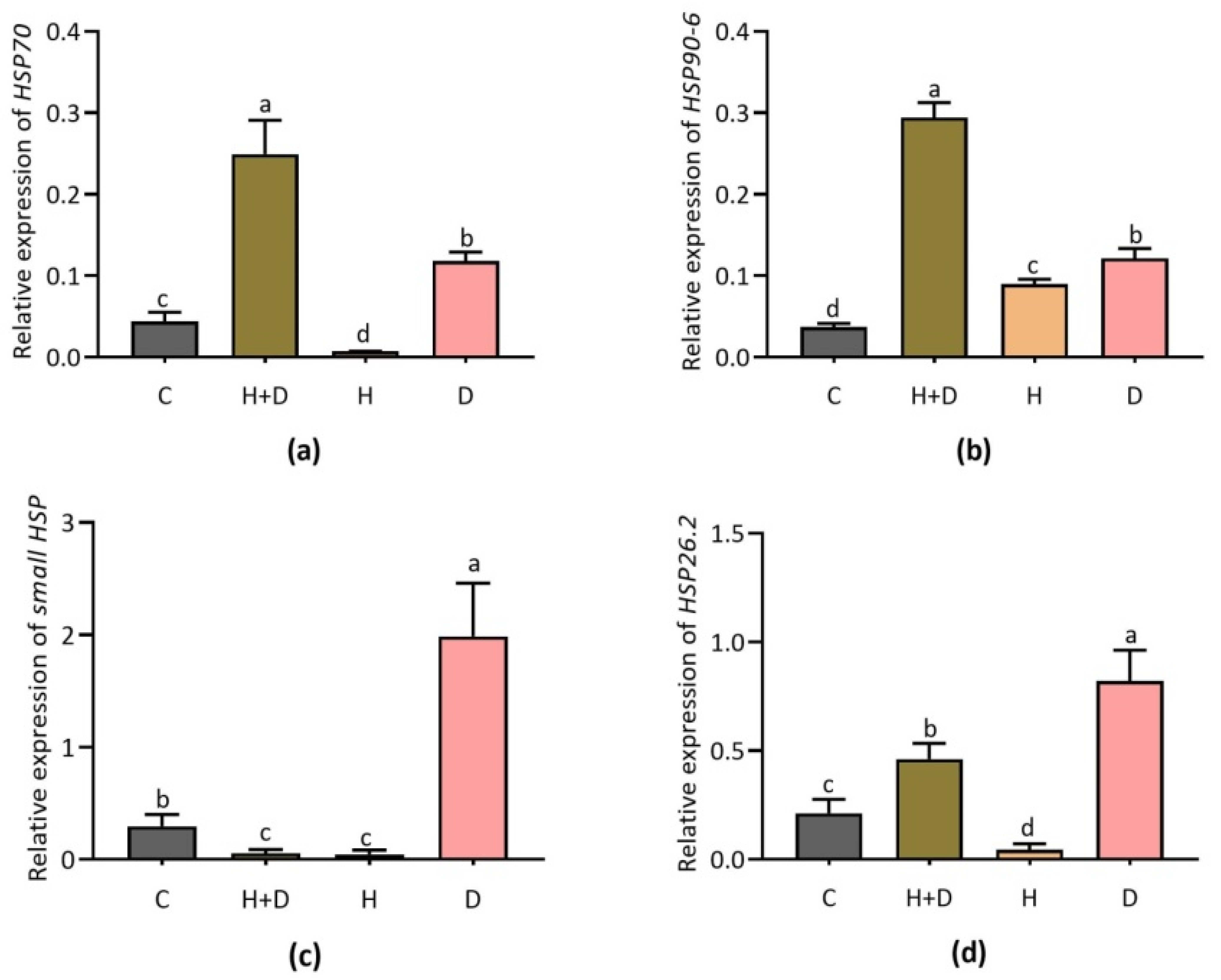
3.2. Expression of Key Heat Shock Protein Genes Involved in Cellular Homeostasis
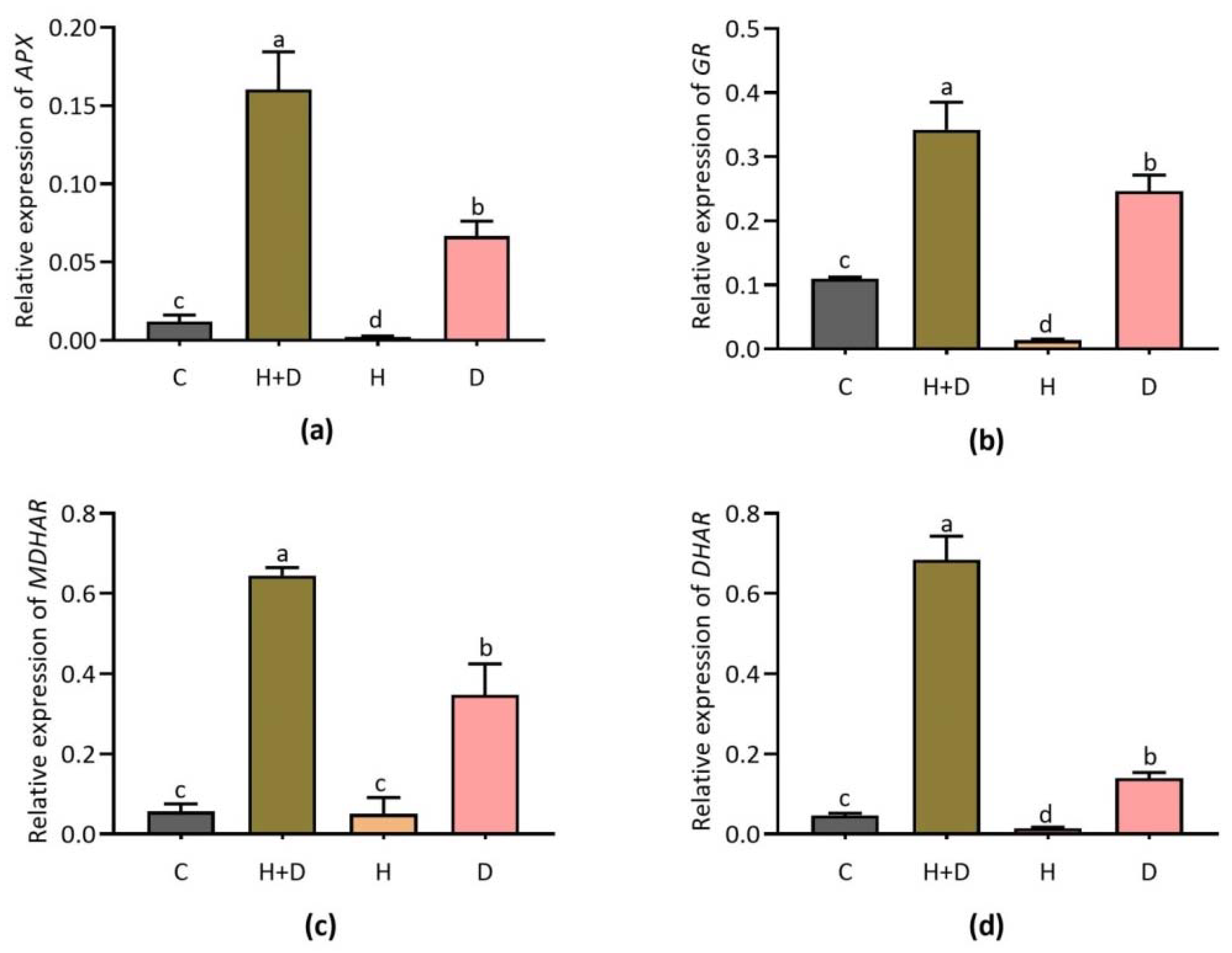
3.3. Expression of Ascorbate-Cycle-Dependent Genes
3.4. Effect of Heat and Drought Stress on Expression Patterns of Glutathione (GSH)-Cycle-Dependent Genes
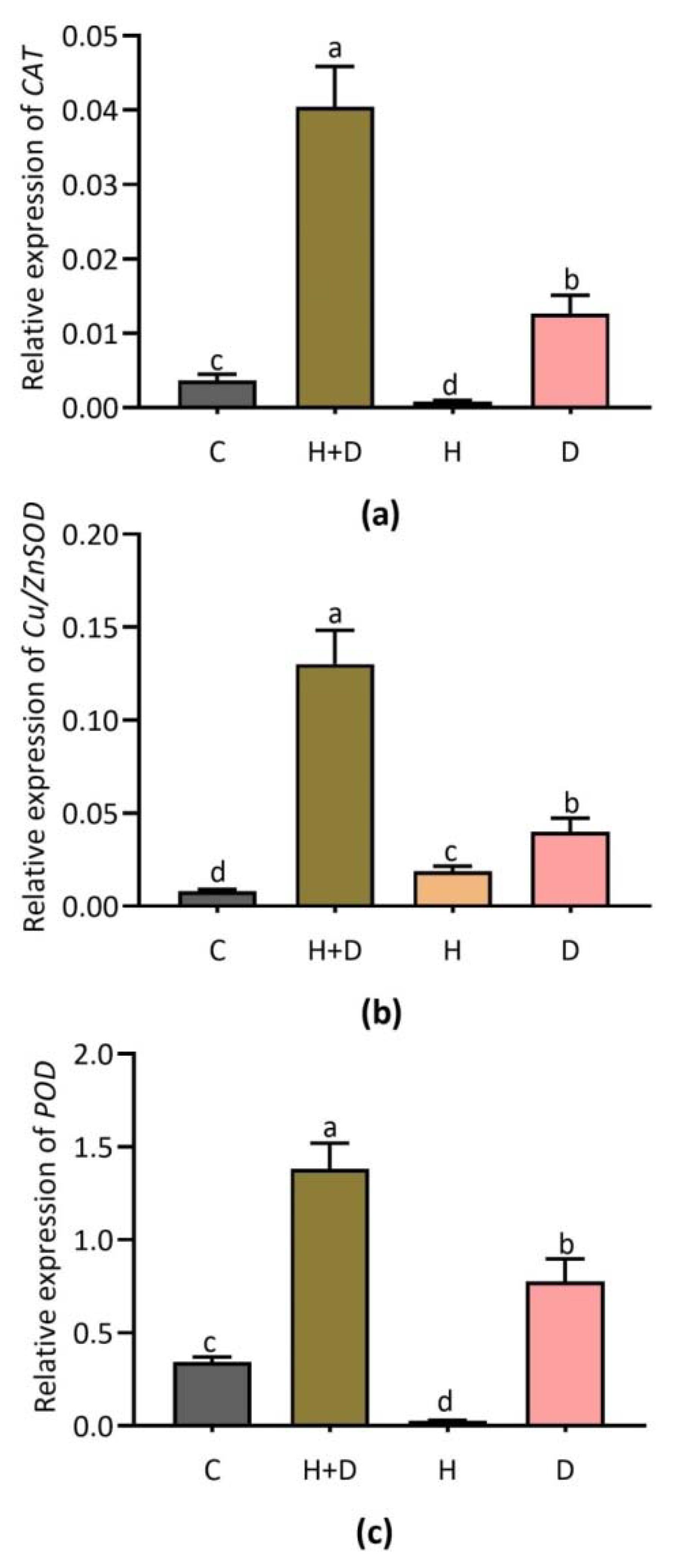
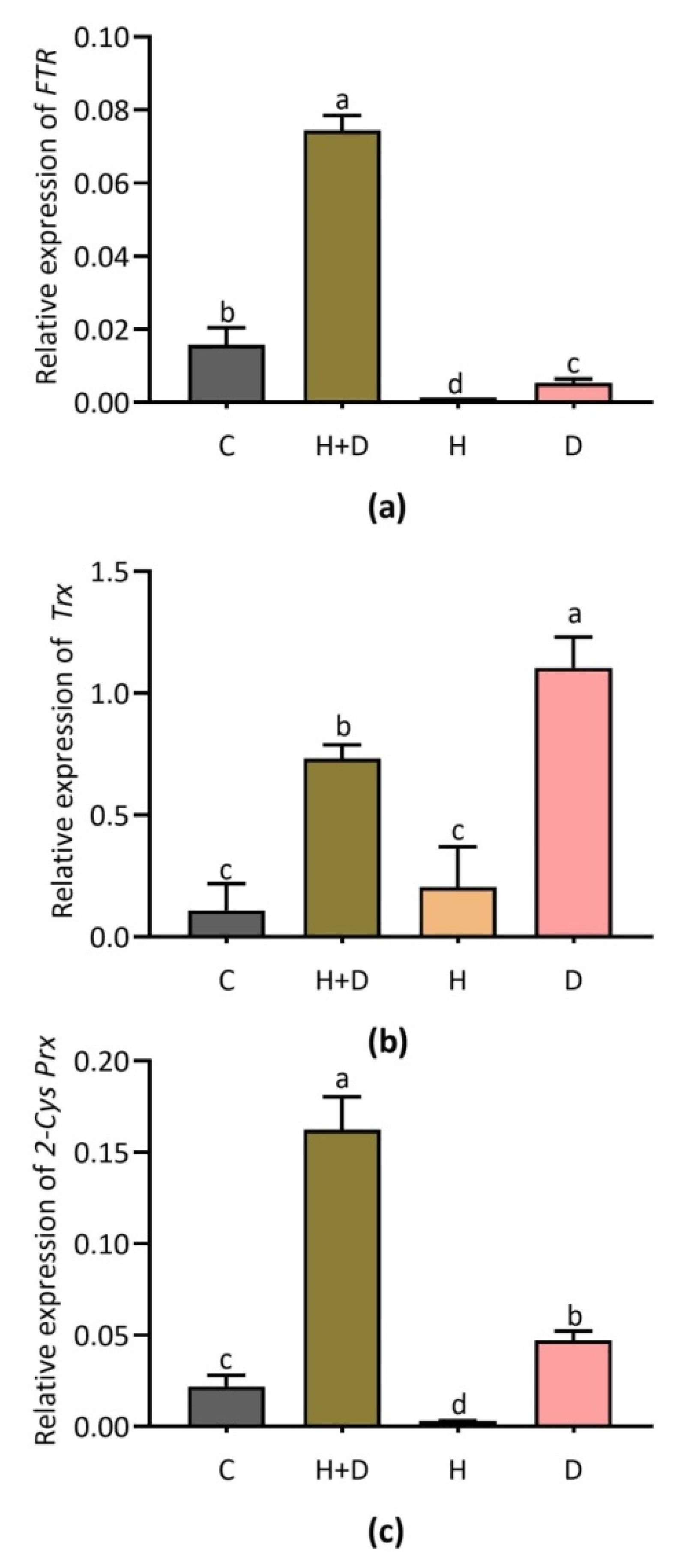
3.5. Effect of Heat and Drought Stress on Key Genes Involved in Antioxidant Defense
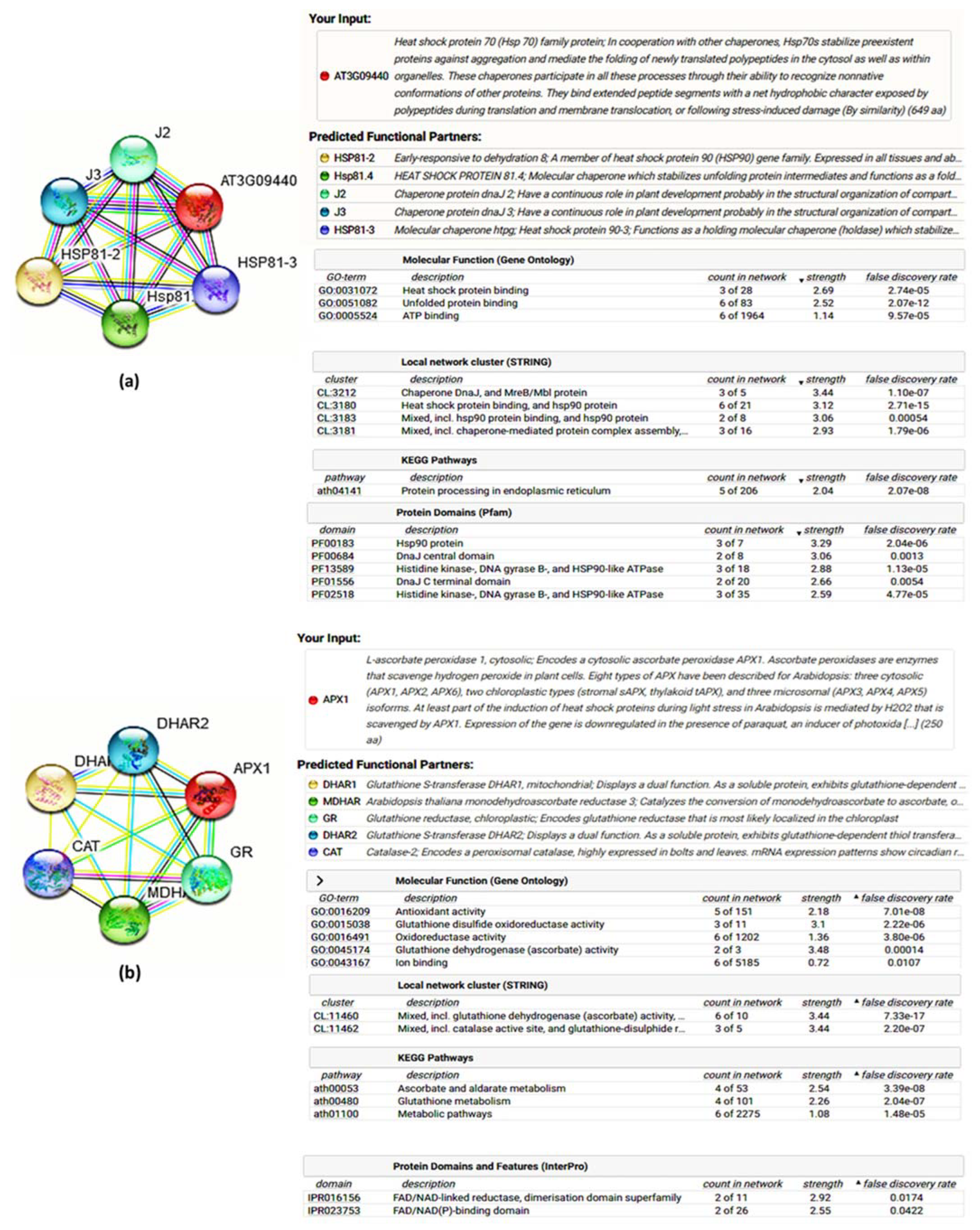
3.6. Analysis of Interactome Map of Heat Shock Protein and Antioxidant Candidates
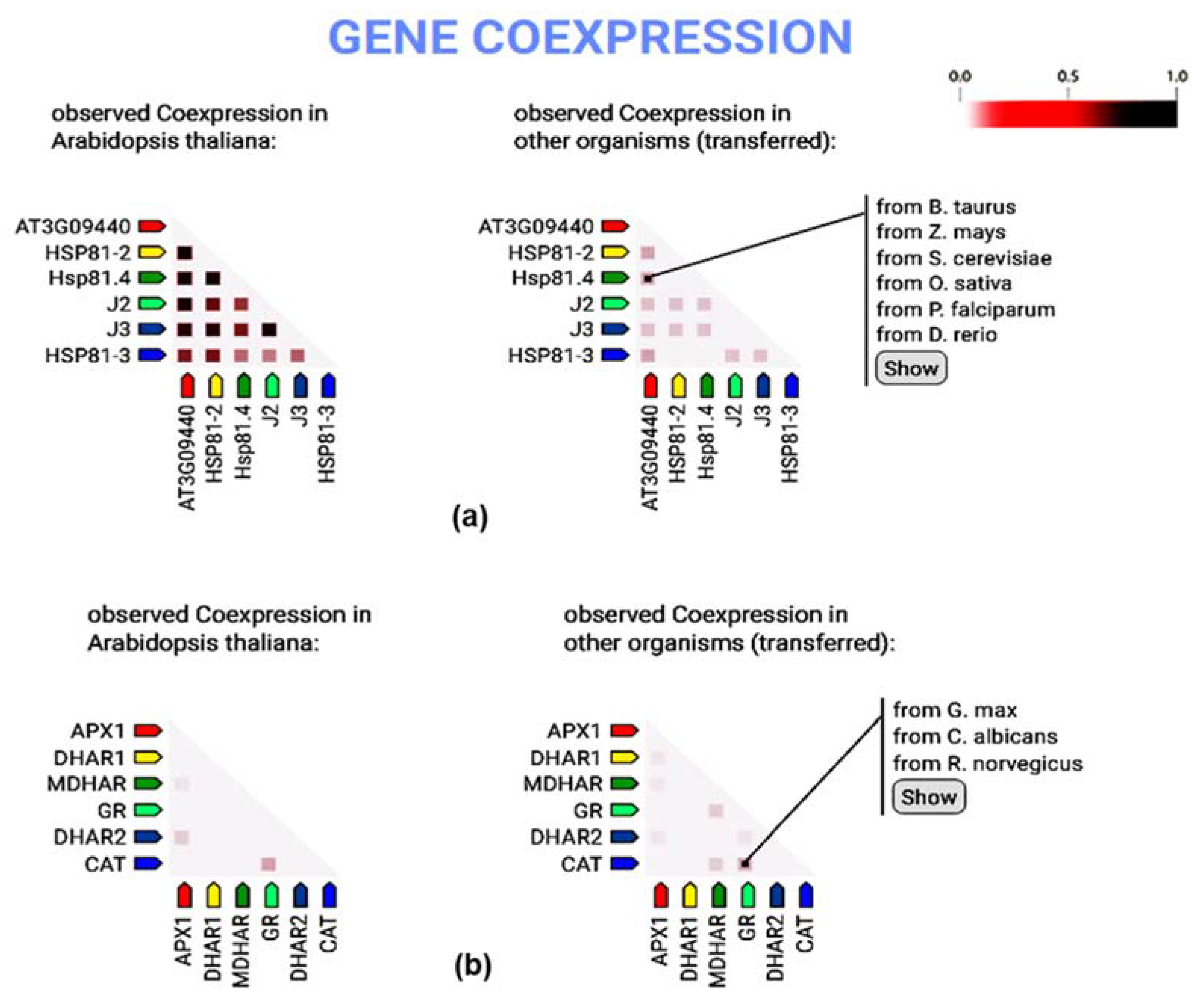
3.7. Gene Co-Expression Analysis of Heat Shock Protein and Antioxidant Candidates
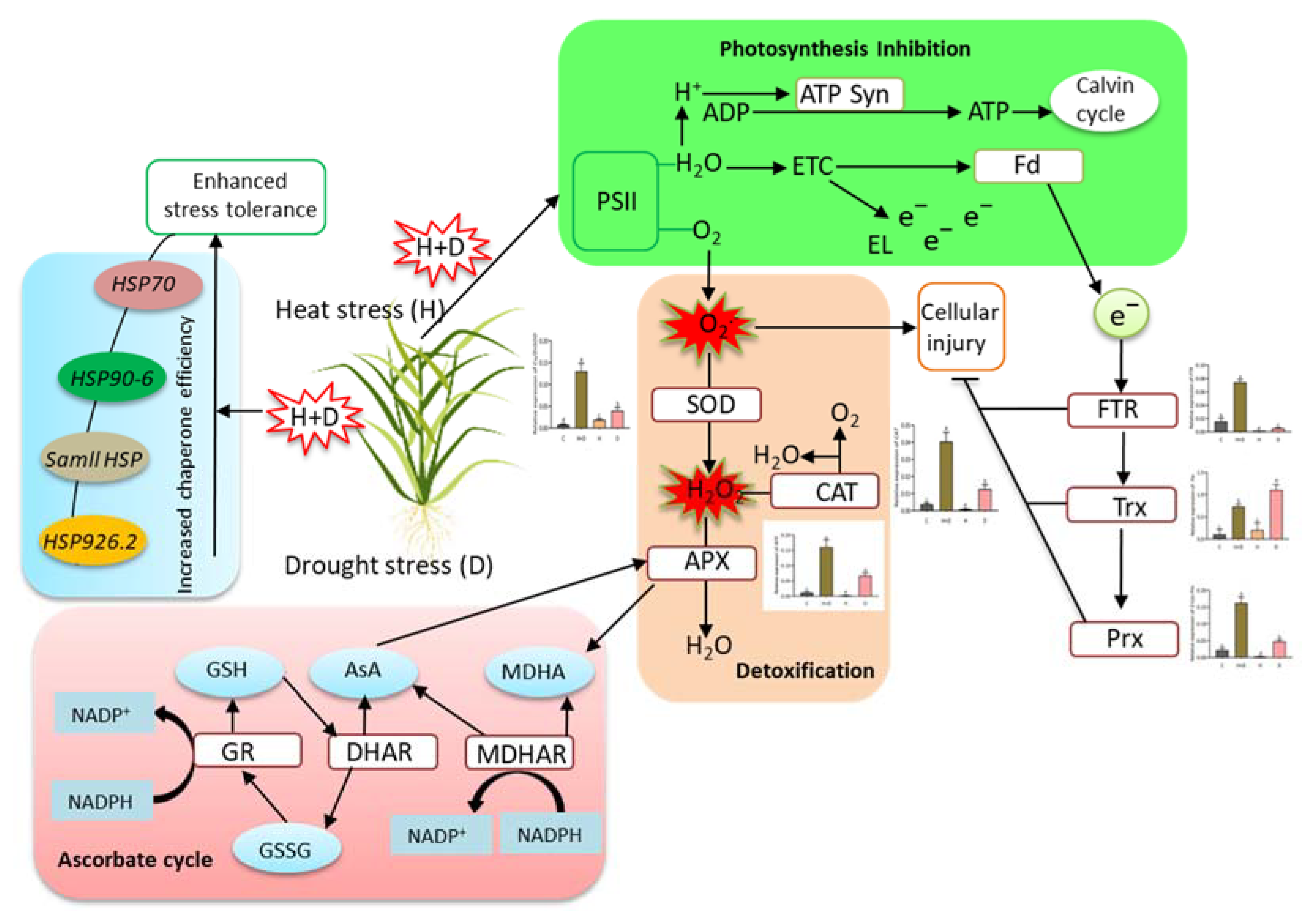
4. Discussion
4.1. Heat Combined with Drought Stress Severely Induced Wilting and Accumulation of Oxidative Stress Indices in Perennial Rye Grass
4.2. Heat Shock Protein Genes Responded to Heat and Drought Stress in Perennial Rye Grass
4.3. ROS-Scavenging Related Antioxidant Genes Involved in Heat and Drought Stress Responses in Perennial Rye Grass
4.4. Interactome Discoveries and Background of Functional Genomics Studies in Grass, and Related Plant Species
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Basavaraj, P.S.; Rane, J. Avenues to realize potential of phenomics to accelerate crop breeding for heat tolerance. Plant Physiol. Rep. 2020, 25, 594–610. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; García-Reyes, M.; Jacox, M.G.; Rykaczewski, R.R.; Black, B.A.; Bograd, S.J.; Sydeman, W.J. Co-occurrence of california drought and northeast pacific marine heatwaves under climate change. Geophys. Res. Lett. 2021, 48, e2021GL092765. [Google Scholar] [CrossRef]
- Ahmed, M.; Hayat, R.; Ahmad, M.; ul-Hassan, M.; Kheir, A.M.S.; ul-Hassan, F.; ur-Rehman, M.H.; Shaheen, F.A.; Raza, M.A.; Ahmad, S. Impact of climate change on dryland agricultural systems: A review of current status, potentials, and further work need. Int. J. Plant Prod. 2022, 16, 341–363. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Liu, H.; Huang, J.; Gao, Z.; Wang, G.; Li, D.; Yu, H.; Chen, X. Accelerated dryland expansion regulates future variability in dryland gross primary production. Nat. Commun. 2020, 11, 1665. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef]
- Al Khateeb, W.; Muhaidat, R.; Alahmed, S.; Al Zoubi, M.S.; Al-Batayneh, K.M.; El-Oqlah, A.; Abo Gamar, M.; Hussein, E.; Aljabali, A.A.; Alkaraki, A.K. Heat shock proteins gene expression and physiological responses in durum wheat (Triticum durum) under salt stress. Physiol. Mol. Biol. Plants 2020, 26, 1599–1608. [Google Scholar] [CrossRef]
- Bourgine, B.; Guihur, A. Heat shock signaling in land plants: From plasma membrane sensing to the transcription of small heat shock proteins. Front. Plant Sci. 2021, 12, 1583. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Rahman, M.A.; Kim, Y.-G.; Lee, B.-H. Proteomic response of alfalfa subjected to aluminum (Al) stress at low pH soil. Je Korean Soc. Grassl. Forage Sci. 2014, 34, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Kabir, A.H.; Song, Y.; Lee, S.-H.; Hasanuzzaman, M.; Lee, K.-W. Nitric oxide prevents Fe deficiency-induced photosynthetic disturbance and oxidative stress in alfalfa by regulating Fe acquisition and antioxidant defense. Antioxidants 2021, 10, 1556. [Google Scholar] [CrossRef] [PubMed]
- He, M.; He, C.Q.; Ding, N.Z. Abiotic stresses: General defenses of land plants and chances for engineering multistress tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar] [CrossRef] [PubMed]
- Zang, D.; Wang, J.; Zhang, X.; Liu, Z.; Wang, Y. Arabidopsis heat shock transcription factor HSFA7b positively mediates salt stress tolerance by binding to an E-box-like motif to regulate gene expression. J. Exp. Bot. 2019, 70, 5355–5374. [Google Scholar] [CrossRef] [PubMed]
- Qamer, Z.; Chaudhary, M.T.; Du, X.; Hinze, L.; Azhar, M.T. Review of oxidative stress and antioxidative defense mechanisms in Gossypium hirsutum L. in response to extreme abiotic conditions. Cotton Res. 2021, 4, 9. [Google Scholar] [CrossRef]
- Ul Haq, S.; Khan, A.; Ali, M.; Khattak, A.M.; Gai, W.X.; Zhang, H.X.; Wei, A.M.; Gong, Z.H. Heat shock proteins: Dynamic biomolecules to counter plant biotic and abiotic stresses. Int. J. Mol. Sci. 2019, 20, 5321. [Google Scholar] [CrossRef]
- Lee, K.-W.; Rahman, M.; Choi, G.J.; Kim, K.-Y.; Ji, H.C.; Hwang, T.Y.; Lee, S.-H.; Sciences, P. Expression of small heat shock protein 23 enhanced heat stress tolerance in transgenic alfalfa plants. J. Anim. Plant Sci. 2017, 27, 1238–1244. [Google Scholar]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef]
- Nadarajah, K.K. ROS homeostasis in abiotic stress tolerance in plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef]
- Kabir, A.H.; Das, U.; Rahman, M.A.; Lee, K.-W. Silicon induces metallochaperone-driven cadmium binding to the cell wall and restores redox status through elevated glutathione in Cd-stressed sugar beet. Physiol Plant. 2021, 173, 352–368. [Google Scholar] [CrossRef]
- Raven, E.L.; Lad, L.; Sharp, K.H.; Mewies, M.; Moody, P.C.E. Defining substrate specificity and catalytic mechanism in ascorbate peroxidase. Biochem. Soc. Symp. 2004, 71, 27–38. [Google Scholar] [CrossRef]
- Sevilla, F.; Camejo, D.; Ortiz-Espín, A.; Calderón, A.; Lázaro, J.J.; Jiménez, A. The thioredoxin/peroxiredoxin/sulfiredoxin system: Current overview on its redox function in plants and regulation by reactive oxygen and nitrogen species. J. Exp. Bot. 2015, 66, 2945–2955. [Google Scholar] [CrossRef] [PubMed]
- Ullah Qadir, S.; Raja, V.; Siddiqui, W.A.; Shah, T.; Alansi, S.; El-Sheikh, M.A. Ascorbate glutathione antioxidant system alleviates fly ash stress by modulating growth physiology and biochemical responses in Solanum lycopersicum. Saudi J. Biol. Sci. 2022, 29, 1322–1336. [Google Scholar] [CrossRef] [PubMed]
- Koh, Y.S.; Wong, S.K.; Ismail, N.H.; Zengin, G.; Duangjai, A.; Saokaew, S.; Phisalprapa, P.; Tan, K.W.; Goh, B.H.; Tang, S.Y. Mitigation of environmental stress-impacts in plants: Role of sole and combinatory exogenous application of glutathione. Front. Plant Sci. 2021, 12, 791205. [Google Scholar] [CrossRef]
- McDonagh, J.; O’Donovan, M.; McEvoy, M.; Gilliland, T.J. Genetic gain in perennial ryegrass (Lolium perenne) varieties 1973 to 2013. Euphytica 2016, 212, 187–199. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, Y.; Tao, X.; Wang, J.-Z.; Cheng, H.-Y.; Yang, H.; Ma, X.-R. Heat shock factor genes of tall fescue and perennial ryegrass in response to temperature stress by RNA-Seq analysis. Front. Plant Sci. 2016, 6, 1226. [Google Scholar] [CrossRef]
- Rahman, M.A.; Alam, I.; Sharmin, S.A.; Kabir, A.H.; Kim, Y.-G.; Liu, G.; Lee, B.-H. Physiological and proteomic analyses reveal the protective roles of exogenous hydrogen peroxide in alleviating drought stress in soybean plants. Plant Biotechnol. Rep. 2021, 15, 805–818. [Google Scholar] [CrossRef]
- Peever, T.L.; Higgins, V.J. Electrolyte leakage, lipoxygenase, and lipid peroxidation induced in tomato leaf tissue by specific and nonspecific elicitors from Cladosporium fulvum. Plant Physiol. Rep. 1989, 90, 867–875. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Soliman, W.S.; Fujimori, M.; Tase, K.; Sugiyama, S.-i. Oxidative stress and physiological damage under prolonged heat stress in C3 grass Lolium perenne. Grassl. Sci 2011, 57, 101–106. [Google Scholar] [CrossRef]
- Soliman, W.S.; Abbas, A.M.; Novak, S.J.; Fujimori, M.; Tase, K.; Sugiyama, S.I. Inheritance of heat tolerance in perennial ryegrass (Lolium perenne, Poaceae): Evidence from progeny array analysis. PeerJ 2021, 9, e11782. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Shao, K.; Huang, Y.; Lei, Y.; Tan, L.; Chan, Z. Natural variation analysis of perennial ryegrass in response to abiotic stress highlights LpHSFC1b as a positive regulator of heat stress. Environ. Exp. Bot. 2020, 179, 104192. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, X.; Goatley, M.; Ervin, E. Heat shock proteins in relation to heat stress tolerance of creeping bentgrass at different N levels. PLoS ONE 2014, 9, e102914. [Google Scholar] [CrossRef]
- Khan, Z.; Shahwar, D. Role of Heat Shock Proteins (HSPs) and Heat Stress Tolerance in Crop Plants. In Sustainable Agriculture in the Era of Climate Change; Roychowdhury, R., Choudhury, S., Hasanuzzaman, M., Srivastava, S., Eds.; Springer: Cham, Switzerland, 2020; pp. 211–234. [Google Scholar]
- Navrot, N.; Rouhier, N.; Gelhaye, E.; Jacquot, J.-P. Reactive oxygen species generation and antioxidant systems in plant mitochondria. Physiol. Plant. 2007, 129, 185–195. [Google Scholar] [CrossRef]
- Jiuxin, L.; Liebao, H. Progress and challenges in china turfgrass abiotic stress resistance research. Front. Plant Sci. 2022, 13, 922175. [Google Scholar] [CrossRef]
- Harb, A.; Awad, D.; Samarah, N. Gene expression and activity of antioxidant enzymes in barley (Hordeum vulgare L.) under controlled severe drought. J. Plant Interact. 2015, 10, 109–116. [Google Scholar] [CrossRef]
- Shah, K.; Nahakpam, S. Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars. Plant Physiol. Biochem. 2012, 57, 106–113. [Google Scholar] [CrossRef]
- Haque, A.F.M.M.; Tasnim, J.; El-Shehawi, A.M.; Rahman, M.A.; Parvez, M.S.; Ahmed, M.B.; Kabir, A.H. The Cd-induced morphological and photosynthetic disruption is related to the reduced Fe status and increased oxidative injuries in sugar beet. Plant Physiol. Biochem. 2021, 166, 448–458. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward | Reverse | Accession Number |
|---|---|---|---|
| HSP70 | CACGATTGGCCCATTCCAAC | CGTCTCGTGGTGCATCATCT | JF747479 |
| HSP90-6 | AAGCTCGGTTGCATGGAAGA | GGAGCATTTTTGGCACTGCT | XM_047191399 |
| small HSP | GGTGAAGATGCGGTTCGACA | ATGTCGTAGGAGCTGACGC | XM_047229918 |
| HSP26.2 | GGCGAGAAGGATGCATGGAA | CTCGATGGCGATCTGGAACA | XM_047189720 |
| APX | GACGTGATTCCTCGGTTTGC | TCAGAGGATCACGGGTCCAT | JF747449 |
| GR | TGGCTTTGGGTGGACTTACG | ACAGCTTGCCATCCACACTA | XM_047233600 |
| MDHAR | CGGTTCAGATGCTGCAAACA | CCATGCAGTGTTTTTCCGGG | JF747451 |
| DHAR | ACCAACGATGGAACAGAGCA | AGTGAATCTGGGACAGACCA | JF747452 |
| CAT | ACAAGTTCGACTTCGACCCG | CAGCATCTTGTCGTCGGAGT | JF747381 |
| Cu/ZnSOD | CGGTGACACAACTAACGGGT | CCCAACAACTGCATTTGGGC | XP_047061758 |
| POD | CGGTTCCATTCGAGGCATGA | TCTTGCTTGCGGTGGTAGAG | XM_047215214 |
| FTR | CCGTCGTCATCAAGGGACTT | GAAATCGTTGTCGGGGGTGA | XM_047224812 |
| Trx | AACTCCTCTCGTGTCAGAGC | TGGGGCGTTACCTTCCAAAA | EL664102 |
| 2-Cys Prx | GGAGTGGACGATTTGCCGTT | ACTCATCGTGTCTGTCGCTG | XM_047225703 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.A.; Woo, J.H.; Song, Y.; Lee, S.-H.; Hasan, M.M.; Azad, M.A.K.; Lee, K.-W. Heat Shock Proteins and Antioxidant Genes Involved in Heat Combined with Drought Stress Responses in Perennial Rye Grass. Life 2022, 12, 1426. https://doi.org/10.3390/life12091426
Rahman MA, Woo JH, Song Y, Lee S-H, Hasan MM, Azad MAK, Lee K-W. Heat Shock Proteins and Antioxidant Genes Involved in Heat Combined with Drought Stress Responses in Perennial Rye Grass. Life. 2022; 12(9):1426. https://doi.org/10.3390/life12091426
Chicago/Turabian StyleRahman, Md Atikur, Jae Hoon Woo, Yowook Song, Sang-Hoon Lee, Md Mahadi Hasan, Md Abul Kalam Azad, and Ki-Won Lee. 2022. "Heat Shock Proteins and Antioxidant Genes Involved in Heat Combined with Drought Stress Responses in Perennial Rye Grass" Life 12, no. 9: 1426. https://doi.org/10.3390/life12091426