

Role of WRKY Transcription Factors in Regulation of Abiotic Stress Responses in Cotton

Abstract
:1. Introduction
2. WRKY Transcription Factors
3. Cotton WRKY Transcription Factors
4. Functions of WRKY TFs in Cotton against Abiotic Stresses
4.1. Drought and Heat Stresses
4.2. Salt Stress
4.3. Cold Stress
4.4. Other Abiotic Stresses
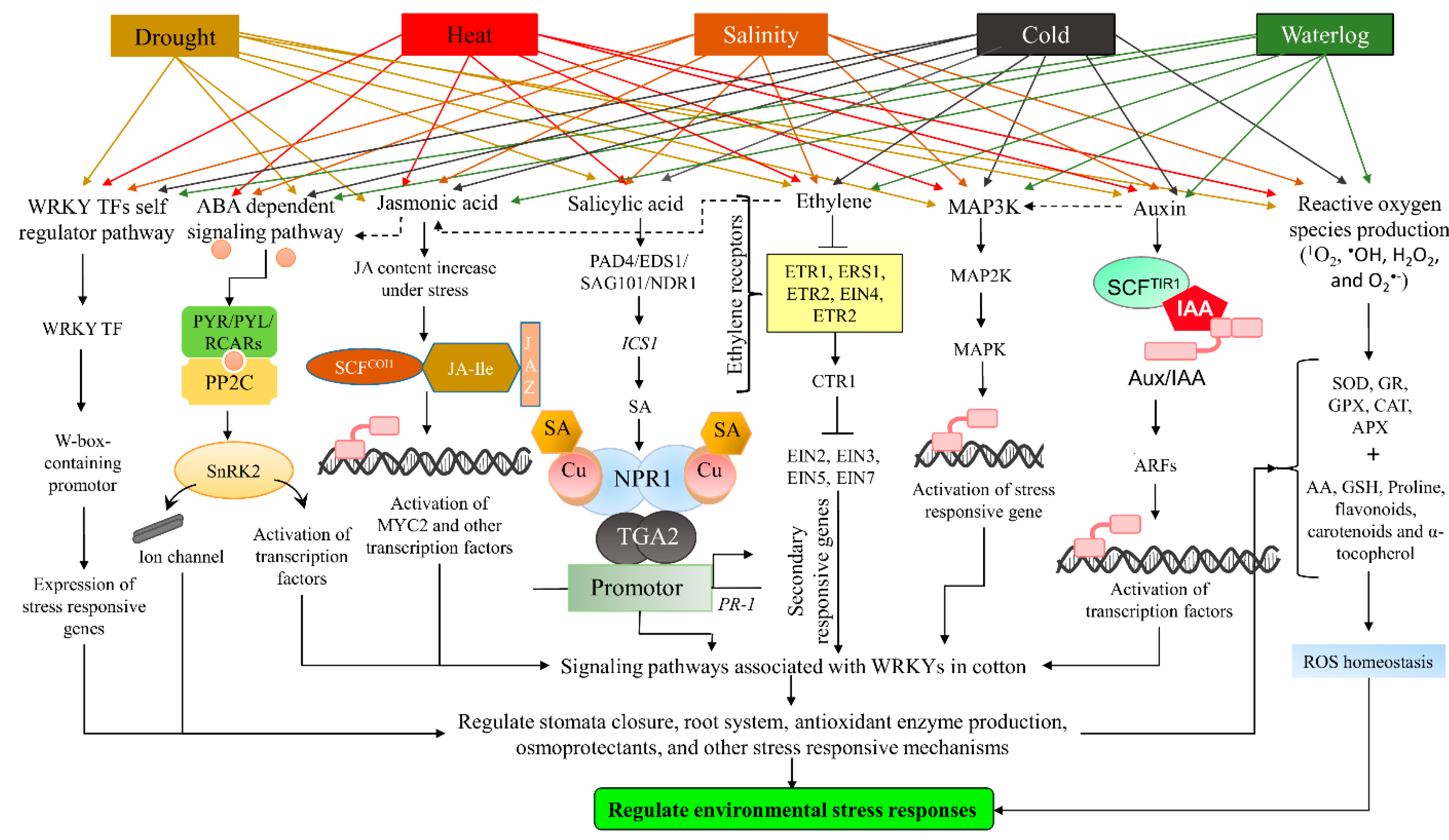
5. Signaling Pathways Associated with WRKY TFs
5.1. Self-Regulatory Pathway
5.2. Abscisic Acid
5.3. Jasmonic Acid, Ethylene, and Salicylic Acid
5.4. Scavenging of ROS
5.5. Kinases (MAPK)
6. Cotton WRKY TFs Genetically Engineered Plants

{kind=link}
| WRKY Gene (Sub Group) | Stress | Cellular Localization | Expression in Plant | Traits Regulated | Reference |
|---|---|---|---|---|---|
| GbWRKY1 (I) | Reduced Phosphorus starvation, and tolerance to drought and salt stresses | Nucleus | Overexpressed in Arabidopsis and cotton | Overexpressing lines reduced the accumulation of anthocyanin and enhanced the activity of phosphatase, MDA content, ion leakage and root inhibition | [68,109] |
| GhWRKY3 (I) | Expressed under drought, salt and low temperature | Nucleus | NA | NA | [110] |
| GarWRKY5 (III) | Enhanced salt tolerance | NA | Silenced in cotton and overexpressed in Arabidopsis | Overexpressing lines exhibited higher activities of SOD, POD and enhanced root length | [55] |
| GhWRKY6 | Regulated drought and salt stresses | NA | Silenced in cotton and overexpressed in Arabidopsis | Overexpressing plants showed shorter root length, larger stomatal aperture, increased H2O2 and MDA level, reduced proline accumulation and participated in ABA signaling pathway | [111] |
| GhWRKY6-like | Improved salt and drought tolerance | Nucleus | Overexpressed in Arabidopsis and silenced in cotton | In overexpressing lines, MDA and H2O2 content were reduced and proline, SOD and POD contents were increased. Expression pattern of ABA signaling pathway genes was also reported higher in overexpressing lines. | [58] |
| GhWRKY15 (IId) | Increased resistant against wounding Viral and fungal infection | Nucleus | Overexpressed in Tobacco | Increased POD, APX and expression of stress related genes | [12] |
| GhWRKY17 (IId) | Enhanced drought and salt sensitivity | Nucleus | Overexpressed in Nicotiana benthamiana | Reduced root length, seed germination, stomatal closure, chlorophyll content and expression pattern of marker genes involved in ABA signaling pathway Increased water loss rate, electrolyte leakage and accumulation of O2−, H2O2, MDA content | [48] |
| GhWRKY25 (I) | Enhanced salt tolerance but reduced drought tolerance and plant defense against fungal pathogen | Nucleus | Overexpressed in Nicotiana benthamiana | Increased MDA, O2−, and H2O2 content Decreased SOD, POD and CAT activities, inhibited root length under drought stress | [56] |
| GhWRKY27a (III) | Reduced drought tolerance | Nucleus | Overexpressed in Nicotiana benthamiana and silenced in cotton | Overexpressing lines exhibited short roots, closed stomata, high content O2−, and H2O2, and low level of drought related genes expression | [49] |
| GhWRKY33 (III) | Reduced drought tolerance | Nucleus | Overexpressed in Arabidopsis | Inhibited seed germination, early seedling growth and root length, reduced sensitivity to ABA | [24] |
| GhWRKY34 (III) | Enhanced salt tolerance | Nucleus | Overexpressed in Arabidopsis | Seed germination, root length, chlorophyll content and expression pattern of stress related genes was higher | [42] |
| GhWRKY39 (IId) | Enhanced resistance to pathogen infection and salinity | Nucleus | Overexpressed in Nicotiana benthamiana | Reduced hydrogen peroxide accumulation and increased the level of APX, CAT, GST and SOD | [112] |
| GhWRKY39-1 (IId) | Enhanced resistance to pathogen infection and salinity | Nucleus | Overexpressed in Nicotiana benthamiana | Enhanced root length and expression pattern of SOD, GST, APX, and CAT. Decreased H2O2 content | [57] |
| GhWRKY40 (IIa) | Enhanced tolerance against wounding | Nucleus | Overexpressed in Nicotiana benthamiana | Decreased level of O2− and H2O2 Transcript level of JA and SA associated genes in overexpressing plants was decreased | [71] |
| GhWRKY41 (III) | Enhanced drought and salinity tolerance | Nucleus | Overexpressed in Nicotiana benthamiana | Reduced MDA, H2O2 content and ABA dependent stomatal opening. Accumulated increased level of SOD, POD and CAT | [41] |
| GhWRKY42 (IId) | Induced drought and salinity stress | Nucleus | Overexpressed in Arabidopsis | Increased senescence | [33] |
| GhWRKY68 (IIc) | Reduced drought and salt tolerance | Nucleus | Overexpressed in Nicotiana benthamiana | Stomatal opening, O2−, H2O2, and MDA content were increased. Reduced total chlorophyll, CAT, SOD and POD content, root length, expression pattern of ABA-dependent pathway genes | [113] |
| GhWRKY91 (IIe) | Enhanced drought tolerance and delayed senescence | NA | Overexpressed in Arabidopsis | Expression pattern of marker genes involved in drought tolerance was significantly higher. | [77] |
7. Concluding Remarks, Complications, and Recommendations
Author Contributions
Funding
Conflicts of Interest
References
- Wang, K.; Wang, Z.; Li, F.; Ye, W.; Wang, J.; Song, G.; Yue, Z.; Cong, L.; Shang, H.; Zhu, S.; et al. The draft genome of a diploid cotton Gossypium raimondii. Nat. Genet. 2012, 4410, 1098–1103. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J.; Scheffler, B.E.; Dennis, E.; Triplett, B.A.; Zhang, T.; Guo, W.; Chen, X.; Stelly, D.M.; Rabinowicz, P.D.; Town, C.D.; et al. Toward sequencing cotton Gossypium genomes. Plant Physiol. 2007, 1454, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Egbuta, M.A.; McIntosh, S.; Waters, D.L.; Vancov, T.; Liu, L. Biological Importance of Cotton By-Products Relative to Chemical Constituents of the Cotton Plant. Molecules 2017, 221, 93. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Y.; Jiang, W.; Fang, L.; Guan, X.; Chen, J.; Zhang, J.; Saski, C.A.; Scheffler, B.E.; Stelly, D.M.; et al. Sequencing of allotetraploid cotton Gossypium hirsutum L. acc. TM–1 provides a resource for fiber improvement. Nat. Biotechnol. 2015, 335, 531–537. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, A.; Li, J.; Zhang, J.; Zhang, B.; Ge, Q.; Jamshed, M.; Lu, Q.; Li, S.; Xiang, X.; et al. Dissecting the genetic basis of fiber quality and yield traits in interspecific backcross populations of Gossypium hirsutum× Gossypium barbadense. Mol. Genet. Genom. 2019, 294, 1385–1402. [Google Scholar] [CrossRef]
- Statista. 2022. Available online: https://www.statista.com/statistics/263055/cotton-production-worldwide-by-top-countries/ (accessed on 12 February 2022).
- He, M.; He, C.Q.; Ding, N.Z. Abiotic Stresses: General Defences of Land Plants and Chances for Engineering Multi stress Tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar] [CrossRef]
- Khan, A.; Tan, D.K.Y.; Afridi, M.Z.; Luo, H.; Tung, S.; Ajab, M.; Fahad, S. Nitrogen fertility and abiotic stresses management in cotton crop: A review. Environ. Sci. Pollut. Res. 2017, 24, 14551–14566. [Google Scholar] [CrossRef]
- Ullah, A.; Sun, H.; Yang, X.; Zhang, X. Drought coping strategies in cotton: Increased crop per drop. Plant Biotechnol. J. 2017, 153, 271–284. [Google Scholar] [CrossRef]
- Lata, R.; Chowdhury, S.; Gond, S.K.; White, J.F., Jr. Induction of abiotic stress tolerance in plants by endophytic microbes. Lett. Appl. Microbiol. 2018, 664, 268–276. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, A. WRKY proteins: Signaling and regulation of expression during abiotic stress responses. Scient. World J. 2015, 2015, 807560. [Google Scholar] [CrossRef] [Green Version]
- Gull, A.; Lone, A.A.; Wani, N.U.I. Biotic and Abiotic Stresses in Plants. In Abiotic and Biotic Stress in Plants; IntechOpen: London, UK, 2019. [Google Scholar]
- Abdelraheem, A.; Esmaeili, N.; O’Connell, M.; Zhang, J. Progress and perspective on drought and salt stress tolerance in cotton. Ind. Crops Prod. 2019, 130, 118–129. [Google Scholar] [CrossRef]
- Roy, S. 2016. Function of MYB domain transcription factors in abiotic stress and epigenetic control of stress response in plant genome. Plant Signal. Behav. 2016, 111, e1117723. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Xu, Y.; Lu, P.; Wang, X.; Li, Z.; Cai, X.; Zhou, Z.; Wang, Y.; Zhang, Z.; Lin, Z.; et al. Salt stress responsiveness of a wild cotton species Gossypium klotzschianum based on transcriptomic analysis. PLoS ONE 2015, 125, e0178313. [Google Scholar] [CrossRef]
- Chen, F.; Hu, Y.; Vannozzi, A.; Wu, K.; Cai, H.; Qin, Y.; Mullis, A.; Lin, Z.; Zhang, L. The WRKY transcription factor family in model plants and crops. Crit. Rev. Plant Sci. 2017, 365, 311–335. [Google Scholar] [CrossRef]
- Yu, F.; Huaxia, Y.; Lu, W.; Wu, C.; Cao, X.; Guo, X. GhWRKY15, a member of the WRKY transcription factor family identified from cotton Gossypium hirsutum L., is involved in disease resistance and plant development. BMC Plant Biol. 2012, 121, 144–161. [Google Scholar] [CrossRef]
- Wani, S.H.; Anand, S.; Singh, B.; Bohra, A.; Joshi, R. WRKY transcription factors and plant defense responses: Latest discoveries and future prospects. Plant Cell Rep. 2021, 40, 1071–1085. [Google Scholar] [CrossRef]
- Bakshi, M.; Oelmüller, R. WRKY transcription factors: Jack of many trades in plants. Plant Signal. Behav. 2014, 92, e27700. [Google Scholar] [CrossRef]
- Song, H.; Wang, P.; Nan, Z.; Wang, X. The WRKY Transcription Factor Genes in Lotus japonicus. Int. J. Genom. 2014, 2014, 420128. [Google Scholar]
- Xie, T.; Chen, C.; Li, C.; Liu, J.; Liu, C.; He, Y. Genome-wide investigation of WRKY gene family in pineapple: Evolution and expression profiles during development and stress. BMC Genom. 2018, 191, 490. [Google Scholar] [CrossRef]
- Jiang, J.; Ma, S.; Ye, N.; Jiang, M.; Cao, J.; Zhang, J. WRKY transcription factors in plant responses to stresses. J. Integr. Plant Biol. 2017, 592, 86–101. [Google Scholar] [CrossRef]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 155, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.N.; Xu, S.W.; Sun, Y.L.; Liu, D.; Zhou, L.; Li, Y.; Li, X.B. The cotton WRKY transcription factor GhWRKY33 reduces transgenic Arabidopsis resistance to drought stress. Sci. Rep. 2019, 91, 724. [Google Scholar] [CrossRef] [PubMed]
- Schluttenhofer, C.; Yuan, L. Regulation of specialized metabolism by WRKY transcription factors. Plant Physiol. 2015, 1672, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Tian, R.; Ma, D.; Wang, S.; Lium, C. Comparative transcriptome study provides insights into acquisition of embryogenic ability in upland cotton during somatic embryogenesis. J. Cotton Res. 2018, 11, 9. [Google Scholar] [CrossRef]
- Ross, C.A.; Liu, Y.; Shen, Q.X.J. The WRKY gene family in rice Oryza sativa. J. Integr. Plant Biol. 2007, 496, 827–842. [Google Scholar] [CrossRef]
- Song, Y.; Gao, J. Genome–wide analysis of WRKY gene family in Arabidopsis lyrata and comparison with Arabidopsis thaliana and Populus trichocarpa. Chin. Sci. Bull. 2014, 598, 754–765. [Google Scholar] [CrossRef]
- Huang, X.; Li, K.; Xu, X.; Yao, Z.; Jin, C.; Zhang, S. Genome-wide analysis of WRKY transcription factors in white pear Pyrus bretschneideri reveals evolution and patterns under drought stress. BMC Genom. 2015, 161, 1104. [Google Scholar] [CrossRef]
- Waqas, M.; Azhar, M.T.; Rana, I.A.; Azeem, F.; Ali, M.A.; Nawaz, M.A.; Chung, G.; Atif, R.M. Genome-wide identification and expression analyses of WRKY transcription factor family members from chickpea Cicer arietinum L. reveal their role in abiotic stress-responses. Genes Genom. 2019, 414, 467–481. [Google Scholar] [CrossRef]
- Dou, L.; Zhang, X.; Pang, C.; Song, M.; Wei, H.; Fan, S.; Yu, S. Genome-wide analysis of the WRKY gene family in cotton. Mol. Genet. Genom. 2014, 2896, 1103–1121. [Google Scholar] [CrossRef]
- Ding, M.; Chen, J.; Jiang, Y.; Lin, L.; Cao, Y.; Wang, M.; Zhang, Y.; Rong, J.; Ye, W. Genome-wide investigation and transcriptome analysis of the WRKY gene family in Gossypium. Mol. Genom. 2014, 290, 151–171. [Google Scholar] [CrossRef]
- Gu, L.; Wang, H.; Wei, H.; Sun, H.; Li, L.; Chen, P.; Elasad, M.; Su, Z.; Zhang, C.; Ma, L.; et al. Identification, Expression, and Functional Analysis of the Group IId WRKY Subfamily in Upland Cotton Gossypium hirsutum L. Front. Plant Sci. 2018, 9, 1684. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Wendel, J.F.; Gundlach, H.; Guo, H.; Jenkins, J.; Jin, D.; Llewellyn, D.; Showmaker, K.C.; Shu, S.; Udall, J.; et al. Repeated polyploidization of Gossypium genomes and the evolution of spinnable cotton fibres. Nature 2012, 4927429, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Niu, E.; Du, H.; Zhao, L.; Feng, Y.; Guo, W. Genome-wide analysis of the WRKY transcription factor gene family in Gossypium raimondii and the expression of orthologs in cultivated tetraploid cotton. Crop J. 2014, 2, 87–101. [Google Scholar] [CrossRef]
- Fan, X.; Guo, Q.; Xu, P.; Gong, Y.; Shu, H.; Yang, Y.; Ni, W.; Zhang, X.; Shen, X. Transcriptome-wide identification of salt-responsive members of the WRKY gene family in Gossypium aridum. PLoS ONE 2015, 10, e0216148. [Google Scholar]
- Hande, A.S.; Katageri, I.S.; Jadhav, M.P.; Adiger, S.; Gamanagatti, S.; Padmalatha, K.V.; Dhandapani, G.; Kanakachari, M.; Kumar, P.A.; Reddy, V.S. Transcript profiling of genes expressed during fibre development in diploid cotton Gossypium arboreum L. BMC Genom. 2017, 181, 675. [Google Scholar] [CrossRef]
- Gu, L.; Wei, H.; Wang, H.; Su, J.; Yu, S. Characterization and functional analysis of GhWRKY42, a group IId WRKY gene, in upland cotton Gossypium hirsutum L. BMC Genet. 2018, 191, 48. [Google Scholar] [CrossRef]
- Gu, L.; Dou, L.; Guo, Y.; Wang, H.; Li, L.; Wang, C.; Chen, P.; Elasad, M.; Su, Z.; Zhang, C.; et al. The WRKY transcription factor GhWRKY27 coordinates the senescence regulatory pathway in upland cotton Gossypium hirsutum L. BMC Plant Biol. 2019, 191, 116. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; He, S.P.; Gao, Y.; Wang, N.N.; Lu, R.; Li, X.B. A cotton Gossypium hirsutum WRKY transcription factor GhWRKY22 participates in regulating anther/pollen development. Plant Physiol. Biochem. 2019, 141, 231–239. [Google Scholar] [CrossRef]
- Chu, X.; Wang, C.; Chen, X.; Lu, W.; Li, H.; Wang, X.; Hao, L.; Guo, X. The Cotton WRKY Gene GhWRKY41 Positively Regulates Salt and Drought Stress Tolerance in Transgenic Nicotiana benthamiana. PLoS ONE 2015, 1011, e0143022. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, N.; Gong, S.; Lu, R.; Li, Y.; Li, X. Overexpression of a cotton Gossypium hirsutum WRKY gene, GhWRKY34, in Arabidopsis enhances salt–tolerance of the transgenic plants. Plant Physiol. Biochem. 2015, 96, 311–320. [Google Scholar] [CrossRef]
- Index Mundi. 2020. Available online: https://www.indexmundi.com/agriculture/?country=pk&commodity=cotton&graph=production (accessed on 12 April 2020).
- Shakeel, A. Dawn Newspaper. 2020. Available online: https://www.dawn.com/news/1529289 (accessed on 12 June 2020).
- Loka, D.A.; Oosterhuis, D.M. Water Stress and Reproductive Development in Cotton; Department of Crop, Soil, and Environmental Sciences University of Arkansas: Fayetteville, AR, USA, 2012. [Google Scholar]
- Ullah, A.; Manghwar, H.; Shaban, M.; Khan, A.H.; Akbar, A.; Ali, U.; Ali, E.; Fahad, S. Phytohormones enhanced drought tolerance in plants: A coping strategy. Environ. Sci. Pollut. Res. 2018, 2533, 33103–33118. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, W.; Zhu, Y.M.; Chen, Y.; Qiu, C.W.; Zhu, S.; Wu, F. Genotypic differences in leaf secondary metabolism, plant hormones and yield under alone and combined stress of drought and salinity in cotton genotypes. Physiol. Plant. 2019, 1652, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Jia, H.; Chen, X.; Hao, L.; An, H.; Guo, X. The cotton WRKY transcription factor GhWRKY17 functions in drought and salt stress in transgenic Nicotiana benthamiana through ABA signaling and the modulation of reactive oxygen species production. Plant Cell Physiol. 2014, 5512, 2060–2076. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Jia, H.; Wang, F.; Wang, C.; Liu, S.; Guo, X. Overexpression of GhWRKY27a reduces tolerance to drought stress and resistance to Rhizoctonia solani infection in transgenic Nicotiana benthamiana. Front. Physiol. 2015, 6, 265. [Google Scholar] [CrossRef]
- Hu, Q.; Ao, C.; Wang, X.; Wu, Y.; Du, X. GhWRKY1-like, a WRKY transcription factor, mediates drought tolerance in Arabidopsis via modulating ABA biosynthesis. BMC Plan Biol. 2021, 21, 458. [Google Scholar] [CrossRef]
- Ranjan, A.; Sawant, S. Genome-wide transcriptomic comparison of cotton Gossypium herbaceum leaf and root under drought stress. 3 Biotech 2015, 54, 585–596. [Google Scholar] [CrossRef]
- Negrao, S.; Schmockel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 1191, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sharif, I.; Aleem, S.; Farooq, J.; Rizwan, M.; Younas, A.; Sarwar, G.; Chohan, S.M. Salinity stress in cotton: Effects, mechanism of tolerance and its management strategies. Physiol. Mol. Biol. Plants 2019, 254, 807–820. [Google Scholar] [CrossRef]
- Ashraf, J.; Zuo, D.; Wang, Q.; Malik, W.; Zhang, Y.; Abid, M.A.; Cheng, H.; Yang, Q.; Song, G. Recent insights into cotton functional genomics: Progress and future perspectives. Plant Biotechnol. J. 2018, 163, 699–713. [Google Scholar] [CrossRef]
- Guo, Q.; Zhao, L.; Fan, X.; Xu, P.; Xu, Z.; Zhang, X.; Meng, S.; Shen, X. Transcription Factor GarWRKY5 Is Involved in Salt Stress Response in Diploid Cotton Species Gossypium aridum L. Int. J. Mol. Sci. 2021, 2021, 5244. [Google Scholar] [CrossRef]
- Liu, X.; Song, Y.; Xing, F.; Wang, N.; Wen, F.; Zhu, C. GhWRKY25, a group I WRKY gene from cotton, confers differential tolerance to abiotic and biotic stresses in transgenic Nicotiana benthamiana. Protoplasma 2016, 2535, 1265–1281. [Google Scholar] [CrossRef]
- Shi, W.; Hao, L.; Li, J.; Liu, D.; Guo, X.; Li, H. The Gossypium hirsutum WRKY gene GhWRKY39–1 promotes pathogen infection defense responses and mediates salt stress tolerance in transgenic Nicotiana benthamiana. Plant Cell Rep. 2014, 33, 483–498. [Google Scholar] [CrossRef]
- Ullah, A.; Sun, H.; Yang, X.; Zhang, X. A novel cotton WRKY gene, GhWRKY6—Like, improves salt tolerance by activating the ABA signaling pathway and scavenging of reactive oxygen species. Physiol. Plant. 2018, 1624, 439–454. [Google Scholar] [CrossRef]
- Sanghera, G.S.; Wani, S.H.; Hussain, W.; Singh, N.B. Engineering cold stress tolerance in crop plants. Curr. Genom. 2011, 121, 30. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.K.; Sunkar, R. Gene Regulation during Cold Stress Acclimation in Plants. In Plant Stress Tolerance; Humana Press: Totowa, NJ, USA, 2011; pp. 39–55. [Google Scholar]
- Yuan, P.; Yang, T.; Poovaiah, B.W. Calcium Signaling-Mediated Plant Response to Cold Stress. Int. J. Mol. Sci. 2018, 1912, 3896. [Google Scholar] [CrossRef]
- Vyse, K.; Pagter, M.; Zuther, E.; Hincha, D.K. Deacclimation after cold acclimation-a crucial, but widely neglected part of plant winter survival. J. Exp. Bot. 2019, 7018, 4595–4604. [Google Scholar] [CrossRef]
- Zhu, Y.N.; Shi, D.Q.; Ruan, M.B.; Zhang, L.L.; Meng, Z.H.; Liu, J.; Yang, W.C. Transcriptome analysis reveals crosstalk of responsive genes to multiple abiotic stresses in cotton Gossypium hirsutum L. PLoS ONE 2013, 811, e80218. [Google Scholar] [CrossRef] [PubMed]
- Barrero-Sicilia, C.; Silvestre, S.; Haslam, R.P.; Michaelson, L.V. Lipid remodelling: Unravelling the response to cold stress in Arabidopsis and its extremophile relative Eutrema salsugineum. Plant Sci. 2017, 263, 194–200. [Google Scholar] [CrossRef]
- Pareek, A.; Khurana, A.K.; Sharma, A.; Kumar, R. An overview of signaling regulons during cold stress tolerance in plants. Curr. Genom. 2017, 186, 498–511. [Google Scholar] [CrossRef]
- Phukan, U.J.; Jeena, G.S.; Shukla, R.K. 2016 WRKY transcription factors: Molecular regulation and stress responses in plants. Front. Plant Sci. 2016, 7, 760. [Google Scholar] [CrossRef]
- Hasan, M.M.; Hasan, M.M.; da Silva, J.A.T.; Li, X. Regulation of phosphorus uptake and utilization: Transitioning from current knowledge to practical strategies. Cell. Mol. Biol. Lett. 2016, 211, 7. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Jin, L.; Long, L.; Liu, L.; He, X.; Gao, W.; Zhu, L.; Zhang, X. Overexpression of GbWRKY1 positively regulates the Pi starvation response by alteration of auxin sensitivity in Arabidopsis. Plant Cell Rep. 2012, 3112, 2177–2188. [Google Scholar] [CrossRef]
- León, J.; Rojo, E.; Sánchez-Serrano, J.J. Wound signalling in plants. J. Exp. Bot. 2001, 52, 1–9. [Google Scholar] [CrossRef]
- Savatin, D.V.; Gramegna, G.; Modesti, V.; Cervone, F. Wounding in the plant tissue: The defense of a dangerous passage. Front. Plant Sci. 2014, 5, 470. [Google Scholar] [CrossRef]
- Wang, X.; Yan, Y.; Li, Y.; Chu, X.; Wu, C.; Guo, X. GhWRKY40, a multiple stress–responsive cotton WRKY gene, plays an important role in the wounding response and enhances susceptibility to Ralstonia solanacearum infection in transgenic Nicotiana benthamiana. PLoS ONE 2014, 94, e93577. [Google Scholar] [CrossRef]
- Ullah, A.; Heng, S.; Munis, M.F.H.; Fahad, S.; Yang, X. Phytoremediation of heavy metals assisted by plant growth promoting PGP bacteria: A review. Environ. Exp. Bot. 2015, 117, 28–40. [Google Scholar] [CrossRef]
- Ogawa, I.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Time course analysis of gene regulation under cadmium stress in rice. Plant Soil 2009, 325, 97. [Google Scholar] [CrossRef]
- Kaur, R.; Bhatti, S.S.; Singh, S.; Singh, J.; Singh, S. Phytoremediation of heavy metals using cotton plant: A field analysis. Bull. Environ. Contam. Toxicol. 2018, 1015, 637–643. [Google Scholar] [CrossRef]
- Rushton, D.L.; Tripathi, P.; Rabara, R.C.; Lin, J.; Ringler, P.; Boken, A.K.; Langum, T.J.; Smidt, L.; Boomsma, D.D.; Emme, N.J.; et al. WRKY transcription factors: Key components in abscisic acid signalling. Plant Biotechnol. J. 2012, 10, 2–11. [Google Scholar] [CrossRef]
- Besseau, S.; Li, J.; Palva, E.T. WRKY54 and WRKY70 co-operate as negative regulators of leaf senescence in Arabidopsis thaliana. J. Exp. Bot. 2012, 637, 2667–2679. [Google Scholar] [CrossRef]
- Gu, L.; Ma, Q.; Zhang, C.; Wang, C.; Wei, H.; Wang, H.; Yu, S. The Cotton GhWRKY91 Transcription Factor Mediates Leaf Senescence and Responses to Drought Stress in Transgenic Arabidopsis thaliana. Front. Plant Sci. 2019, 10, 1352. [Google Scholar] [CrossRef]
- Xiong, L.; Schumaker, K.S.; Zhu, J.K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14 (Suppl. S1), S165–S183. [Google Scholar] [CrossRef] [Green Version]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N.; Yadav, G.; Singh, J.; Mishra, R.K.; Kumar, V.; Verma, R.; Upadhyay, R.G.; Pandey, M.; et al. Abscisic acid signaling and abiotic stress tolerance in plants: A review on current knowledge and future prospects. Front. Plant Sci. 2017, 8, 161. [Google Scholar] [CrossRef]
- Wasilewska, A.; Vlad, F.; Sirichandra, C.; Redko, Y.; Jammes, F.; Valon, C.; Frey, N.F.D.; Leung, J. An update on abscisic acid signaling in plants and more. Mol. Plant 2008, 12, 198–217. [Google Scholar] [CrossRef]
- Ilyas, M.; Nisar, M.; Khan, N.; Hazrat, A.; Khan, A.H.; Hayat, K.; Fahad, S.; Khan, A.; Ullah, A. Drought Tolerance Strategies in Plants: A Mechanistic Approach. J. Plant Growth Regul. 2020, 40, 926–944. [Google Scholar] [CrossRef]
- Kim, T.H.; Böhmer, M.; Hu, H.; Nishimura, N.; Schroeder, J.I. Guard cell signal transduction network: Advances in understanding abscisic acid, CO2, and Ca2+ signaling. Annu. Rev. Plant Biol. 2010, 61, 561–591. [Google Scholar] [CrossRef]
- Yu, F.; Lou, L.; Tian, M.; Li, Q.; Ding, Y.; Cao, X.; Wu, Y.; Belda-Palazon, B.; Rodriguez, P.L.; Yang, S.; et al. ESCRT-I component VPS23A affects ABA signaling by recognizing ABA receptors for endosomal degradation. Mol. Plant 2016, 912, 1570–1582. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Shinozaki, K.Y.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 4436–4442. [Google Scholar] [CrossRef]
- Sun, H.; Chen, L.; Li, J.; Hu, M.; Ullah, A.; He, X.; Yang, X.; Zhang, X. The JASMONATE ZIM-domain gene family mediates JA signaling and stress response in cotton. Plant Cell Physiol. 2017, 5812, 2139–2154. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 462. [Google Scholar] [CrossRef]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.P.; Sun, S.C.; Zhang, X.Y.; Li, Y.J.; Liu, F.; Zhu, Q.H.; Xue, F.; Sun, J. GhWRKY70D13 regulates resistance to Verticillium dahliae in Cotton through the Ethylene and Jasmonic Acid signaling pathways. Front. Plant Sci. 2020, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 905, 856–867. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. The roles of reactive oxygen metabolism in drought: Not so cut and dried. Plant Physiol. 2014, 1644, 1636–1648. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Fichman, Y.; Devireddy, A.R.; Sengupta, S.; Azad, R.K.; Mittler, R. Systemic signaling during abiotic stress combination in plants. Proc. Natl. Acad. Sci. USA 2020, 24, 14551–14566. [Google Scholar] [CrossRef]
- Huang, H.; Ullah, F.; Zhou, D.X.; Yi, M.; Zhao, Y. Mechanisms of ROS Regulation of Plant Development and Stress Responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- de Zelicourt, A.; Colcombet, J.; Hirt, H. 2016 The role of MAPK modules and ABA during abiotic stress signaling. Trends Plant Sci. 2016, 218, 677–685. [Google Scholar] [CrossRef]
- Danquah, A.; de Zelicourt, A.; Colcombet, J.; Hirt, H. The role of ABA and MAPK signaling pathways in plant abiotic stress responses. Biotechnol. Adv. 2014, 321, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.K.; Jaggi, M.; Raghuram, B.; Tuteja, N. Mitogen–activated protein kinase signaling in plants under abiotic stress. Plant Signal. Behav. 2011, 62, 196–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wang, X.; Wang, Y.; Chen, Y.; Luo, J.; Li, D.; Li, X. Genome–wide identification and functional characterization of cotton (Gossypium hirsutum) MAPKKK gene family in response to drought stress. BMC Plant Biol. 2020, 20, 217. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, M.; Wang, P.; Cox Jr, K.L.; Duan, L.; Dever, J.K.; Shan, L.; Li, Z.; He, P. Regulation of cotton Gossypium hirsutum drought responses by mitogen-activated protein MAP kinase cascade-mediated phosphorylation of GhWRKY59. New Phytol. 2017, 2154, 1462–1475. [Google Scholar] [CrossRef]
- Kaleem, F.; Shabir, G.; Aslam, K.; Rasul, S.; Manzoor, H.; Shah, S.M.; Khan, A.R. An overview of the genetics of plant response to salt stress: Present status and the way forward. Appl. Biochem. Biotechnol. 2018, 1862, 306–334. [Google Scholar] [CrossRef]
- Takeda, S.; Matsuoka, M. Genetic approaches to crop improvement: Responding to environmental and population changes. Nat. Rev. Genet. 2008, 96, 444–457. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 5757781, 109–118. [Google Scholar] [CrossRef]
- Newell-McGloughlin, M. Nutritionally improved agricultural crops. Plant Physiol. 2008, 1473, 939–953. [Google Scholar] [CrossRef]
- Kumar, K.; Gambhir, G.; Dass, A.; Tripathi, A.K.; Singh, A.; Jha, A.K.; Yadava, P.; Choudhary, M.; Rakshit, S. Genetically modified crops: Current status and future prospects. Planta 2020, 251, 91. [Google Scholar] [CrossRef]
- ISAA. 2020. Available online: http://www.isaaa.org/gmapprovaldatabase/eventslist/default.asp (accessed on 12 March 2020).
- Chen, L.; Song, Y.; Li, S.; Zhang, L.; Zou, C.; Yu, D. The role of WRKY transcription factors in plant abiotic stresses. Biochim. Biophys. Acta Gene Regul. Mech. 2012, 18192, 120–128. [Google Scholar] [CrossRef]
- Gao, H.; Wang, Y.; Xu, P.; Zhang, Z. Overexpression of a WRKY transcription factor TaWRKY2 enhances drought stress tolerance in transgenic wheat. Front. Plant Sci. 2018, 9, 997. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Li, C.; He, X.; Zhang, X.; Zhu, L. ABA signaling is negatively regulated by GbWRKY1 through JAZ1 and ABI1 to affect salt and drought tolerance. Plant Cell Rep. 2020, 392, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Yu, F.; Gao, Z.; An, H.; Cao, X.; Guo, X. GhWRKY3, a novel cotton (Gossypium hirsutum L.) WRKY gene, is involved in diverse stress responses. Mol. Biol. Rep. 2011, 38, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, L.; Zhou, K.; Zhang, Y.; Han, X.; Din, Y.; Ge, X.; Qin, W.; Wang, P.; Li, F.; et al. GhWRKY6 acts as a negative regulator in both transgenic Arabidopsis and cotton during drought and salt stress. Front. Genet. 2019, 10, 392. [Google Scholar] [CrossRef]
- Shi, W.; Liu, D.; Hao, L.; Wu, C.A.; Guo, X.; Li, H. GhWRKY39, a member of the WRKY transcription factor family in cotton, has a positive role in disease resistance and salt stress tolerance. Plant Cell Tissue Organ Cult. 2014, 1181, 17–32. [Google Scholar] [CrossRef]
- Jia, H.; Wang, C.; Wang, F.; Liu, S.; Li, G.; Guo, X. GhWRKY68 reduces resistance to salt and drought in transgenic Nicotiana benthamiana. PLoS ONE 2015, 10, e0120646. [Google Scholar]
- Ayaz, A.; Huang, H.; Zheng, M.; Zaman, W.; Li, D.; Saqib, S.; Zhao, H.; Lü, S. Molecular Cloning and Functional Analysis of GmLACS2-3 Reveals Its Involvement in Cutin and Suberin Biosynthesis along with Abiotic Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 9175. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Ullah, A.; Siuta, D.; Kukfisz, B.; Iqbal, S. Role of WRKY Transcription Factors in Regulation of Abiotic Stress Responses in Cotton. Life 2022, 12, 1410. https://doi.org/10.3390/life12091410
Guo X, Ullah A, Siuta D, Kukfisz B, Iqbal S. Role of WRKY Transcription Factors in Regulation of Abiotic Stress Responses in Cotton. Life. 2022; 12(9):1410. https://doi.org/10.3390/life12091410
Chicago/Turabian StyleGuo, Xiaoqiang, Abid Ullah, Dorota Siuta, Bożena Kukfisz, and Shehzad Iqbal. 2022. "Role of WRKY Transcription Factors in Regulation of Abiotic Stress Responses in Cotton" Life 12, no. 9: 1410. https://doi.org/10.3390/life12091410