3.1. Construction of High-Pressure Reactor and Performance of HPR-30
The principle of HPR-30 is based on our previous HFR, which was originally designed for monitoring the millisecond-to-second time scale under hydrothermal conditions. The original HFR consists of a water reservoir, high-pressure pump, sample injector, high-temperature reactor with heating block, cooling bath, pressure controller, and sampling port. In the heating block, a narrow tubing reactor is settled for performing high-temperature reactions, and this was applied for the present HPR-30. The pressure inside the reactor tubing is controlled with the pressure regulator so as to be higher than the vapor pressure of water, in order to keep water in a liquid state. The residence time of the sample exposed to the high temperature is easily regulated by the flow rate of the pump or by using different sizes of tubing reactor. This enables short-time monitoring of the reactions, such as in the millisecond-to-second time scale (0.002–120 s) using different sizes of tubing (inner diameter: 0.015–0.25 mm). For instance, ca. 3–120 s residence times can be imposed using stainless-steel tubing of 0.25 mm inner-diameter and 2 m in length, which possesses ca. 0.1 mL inner volume, by regulating the flow rate to be within ca. 2 mL/min–0.05 mL/min. The system was initially built for monitoring short time scale reactions so that the residence time is suitable for hydrothermal reactions with relatively fast reaction rates. However, this short time scale is not well suited to the reaction time scale of ASBVd(−):HHR, which generally proceeds within the time scale from a few minutes to a few hours [
19]. It is necessary to be able to work under different conditions for the measurement of ribozyme reactions.
Initially, we attempted to adjust the flow rate to a very low value. The longest residence time exposed at high pressure is limited to 100 min, corresponding to the minimum adjustable flow rate of 0.001 mL/min. However, controlling the flow rate at 0.001 mL/min is not practical because pressure regulation is difficult and the accuracy of flow rate control decreases at a very low flow rate. Thus, we adopted the method mentioned in the experimental section, which is applicable in principle to much longer reaction times at high pressures than those tested in this study, of a few hours and longer (
Figure 1). The flow path to the high-pressure tubing is switched to isolate the high-pressure tubing from the main flow of the system, as shown in the left of
Figure 1b, which enables the high-pressure tubing to be a high-pressure vessel which is maintained for the long reaction time scale. There is no upper limit on the residence time unless the sample leaks from the flow reactor tubing.
The reaction pressure in HFR, for instance at 250 °C, is normally controlled at 5–10 MPa since the vapor pressure of water at 250 °C is ca. 4 MPa. We have previously determined that this kind of system is resistant up to 400 °C with a pressure normally adjusted to below 30 MPa. According to our previous studies regarding the hydrolytic degradations of simpler biomolecules, we did not observe any pressure influence under hydrothermal conditions in such cases [
7,
9,
16]. It is reasonable to think that the conformation of simple molecules in the activated state is less dependent on pressure. In the present study, the performance of the system needed to be evaluated with a reaction system using complex organic molecules, such as ribozyme cleavage, for which the pressure influence has already been identified in previous studies [
8,
10,
15].
The maximum pressure which can be imposed is normally determined by the limit pressure of the system components. In the present system, these components consist of the pump, the sample injector, the valve, the pressure regulator, etc. Thus, we built a system which can be used at up to 30 MPa by considering the upper-limit pressure of the pump. In other words, this fact indicates that the maximum pressure could be increased by using components which can be used at higher pressures as a future improvement.
The timing at which the sample solutions reach the inlet and the outlet of the high-pressure tubing was determined by injecting 0.1 M HCl solution and checking the color change of pH test paper. The timing for recovering the samples exposed to high pressure after the switching valve was changed to the main flow was also determined in the same way. In addition, we confirmed that we can recover the RNA molecules by switching the 6-port valves properly. According to our previous study regarding the diffusion of sample solutions, it was estimated that the sample solution is diluted by a factor of 1.55 [
25] in our typical hydrothermal flow reactors. From the measurement of the elution of HCl solution in the present study, the sample is approximately diluted by a factor of 2 at a flow rate of 0.2 mL/min. This indicates that the 0.1 mL sample solution should be recovered in within 1 min. However, we confirmed that the recovery of 0.4–0.6 mL samples gives better results for the ASBVd(−):HHR cleavage reaction. This seems to be due to the low concentration of the ASBVd(−):HHR, resulting in stronger dilution as compared to the 0.1 M HCl solution. Thus, we adjusted the recovery of 0.4–0.6 mL samples at the sampling port for the 0.1 mL injection of ASBVd(−):HHR solution. Furthermore, for the sample analysis by HPLC, we found that the concentration was occasionally useful for sample analysis by using a centrifugation evaporator to adapt the sample volume for HPLC analysis. The concentration did not affect the cleavage reactions provided that the reaction was stopped by the addition of EDTA.
Based on the reaction profiles using HPR-30, we confirmed that the HPR-30 showed good performance for monitoring reactions at high pressures at temperatures up to 65 °C and a residence time of 120 min at 30 MPa. Theoretically, the residence time of the sample exposed to high pressure and high temperature is to be controlled for a very long time as compared to our previous methods [
7,
9,
16] and literature reports using flow systems [
18]. On the other hand, the upper temperature limit should be 150 °C since that is the limit for the switching 6-port valves. The accuracy of the reaction curves in the next section indicates that an influence of the dead volume connecting between the high-pressure tubing and the switching 6-port valves was not observed.
3.2. Cleavage of ASBVd(−):HHR at 0.1 MPa and 30 MPa at 45–70 °C
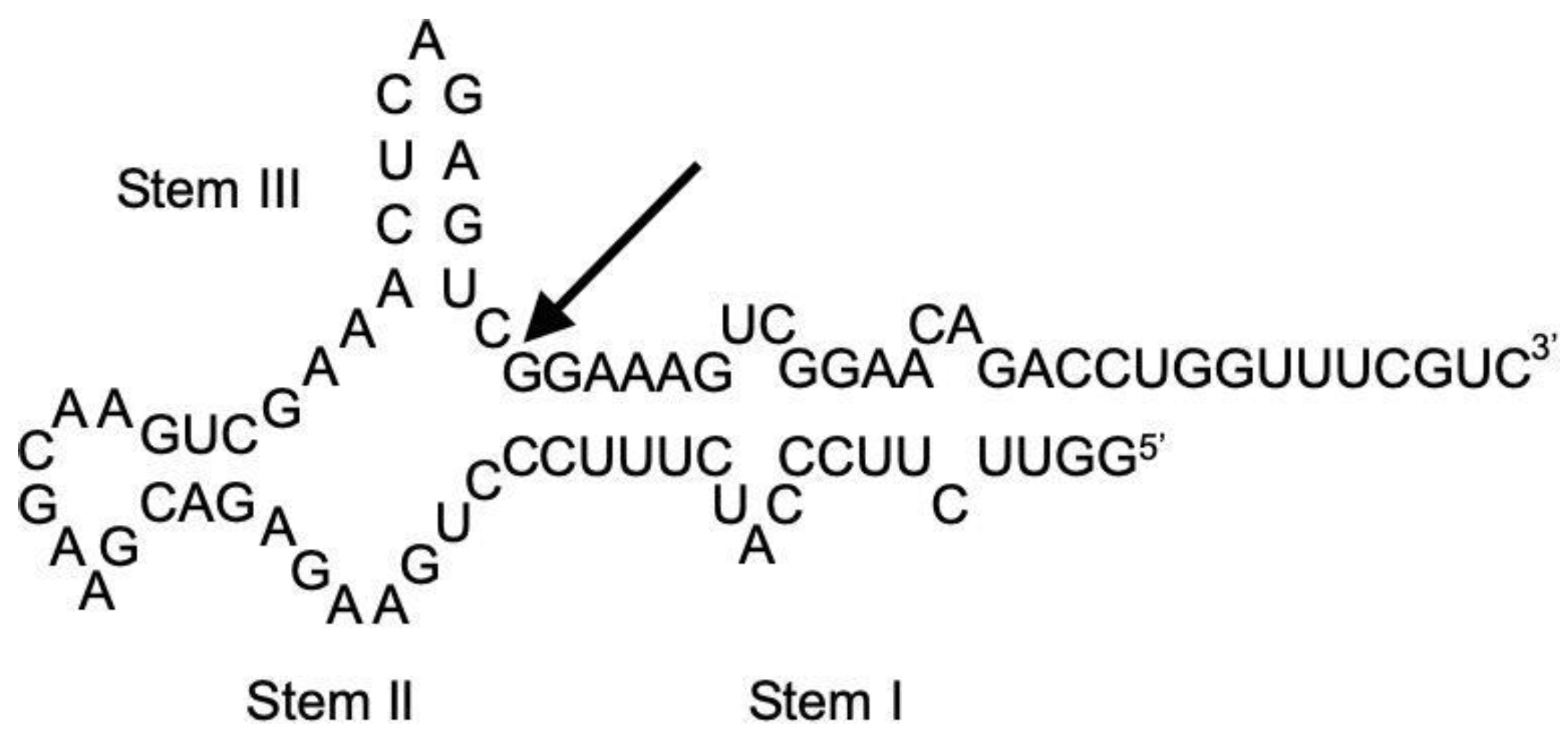
The cleavage of ASBVd(−):HHR using PCR test tubing was carried out at temperatures of 45–70 °C at 0.1 MPa. Reaction curves are shown in
Figure 3. According to our previous study of ASBVd(−):HHR, the self-cleavage of ribozyme proceeds to form two oligonucleotides with 28 and 51 nucleotide units (28 nt and 51 nt). The self-cleavage reaction obeys first-order kinetics, so the lower concentration should basically not affect the reaction behavior (
Supplementary data, Figure S1). The HPLC conditions were optimized for separation of ASBVd(−):HHR (79nt) and its two cleavage products (51nt, 28nt) by using a gradient mode for HPLC using NaCl in the presence of 7.5 M Urea. Three main peaks were observed, one of which corresponds to the ASBVd(−):HHR, with a retention time of ~20 min on the column with 4.6 mm diameter. A treatment for denaturing of ASBVd(−):HHR prior to HPLC analysis did not affect the retention time of ASBVd(−):HHR. According to the literature [
21,
22], running HPLC at high temperatures seems to be useful. However, this is not the case for the present system, so it was possible to run HPLC at 35 °C. The retention time of one of the two reaction products (~12 min) was confirmed as being identical to that of the oligonucleotide with 28 nucleotide units, so it is reasonable that the other peak at ~18 min corresponds to the 51 nt fragmentation product. Thus, we are able to follow the self-cleavage reaction straightforwardly by using the present HPLC system.
The measurements using either the ribozyme prepared by in vitro transcription in our laboratory or that prepared commercially in FASMAC did not show any difference in their cleavage behavior. According to our previous study, the self-cleavage is dominant at temperatures up to 60 °C, while partially random degradative hydrolysis of phosphodiester bonds within the ribozyme sequence occurs with increasing temperature, especially over 65 °C [
19]. We confirmed that the reaction was observable up to 70 °C in batch reactions, that it still followed first-order kinetics, and that there was no notable influence of the random degradation of the ribozyme on the cleavage analysis. The first-order rate constants at temperatures of 45–70 °C are summarized in
Table 1. The influence of random hydrolysis is not notable, so the determination of the first-order rate constants was based on the ratio of 79 nt among the total amount of 28 nt, 51 nt, and 79 nt as per our previous study [
19]. The temperature dependence of the rate constants shows that they are maximum at 55 °C. This is consistent with our previous study [
19]. In addition, this is reasonable since the cleavage rate of ASBVd was also maximum at 55 °C [
15].
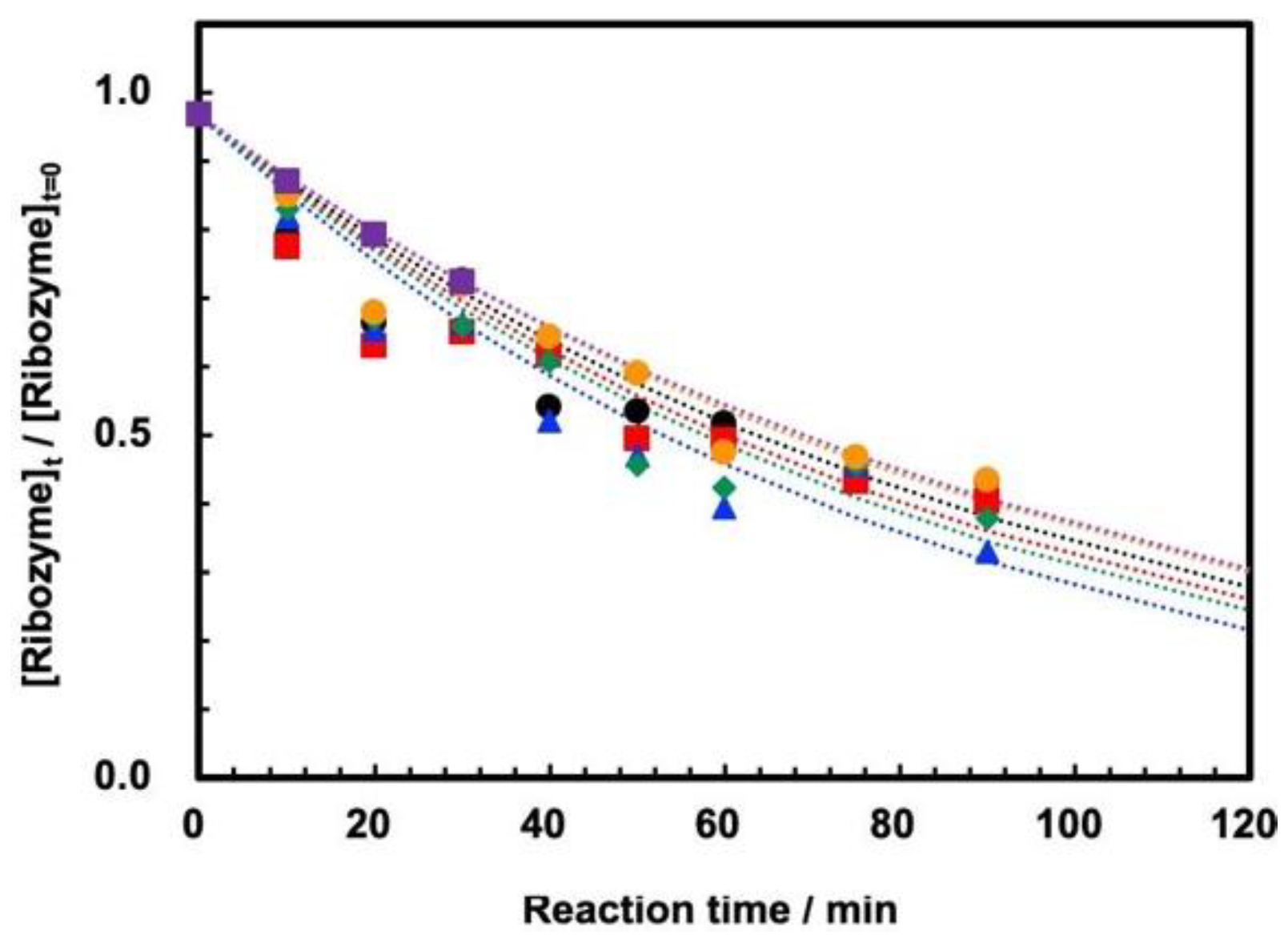
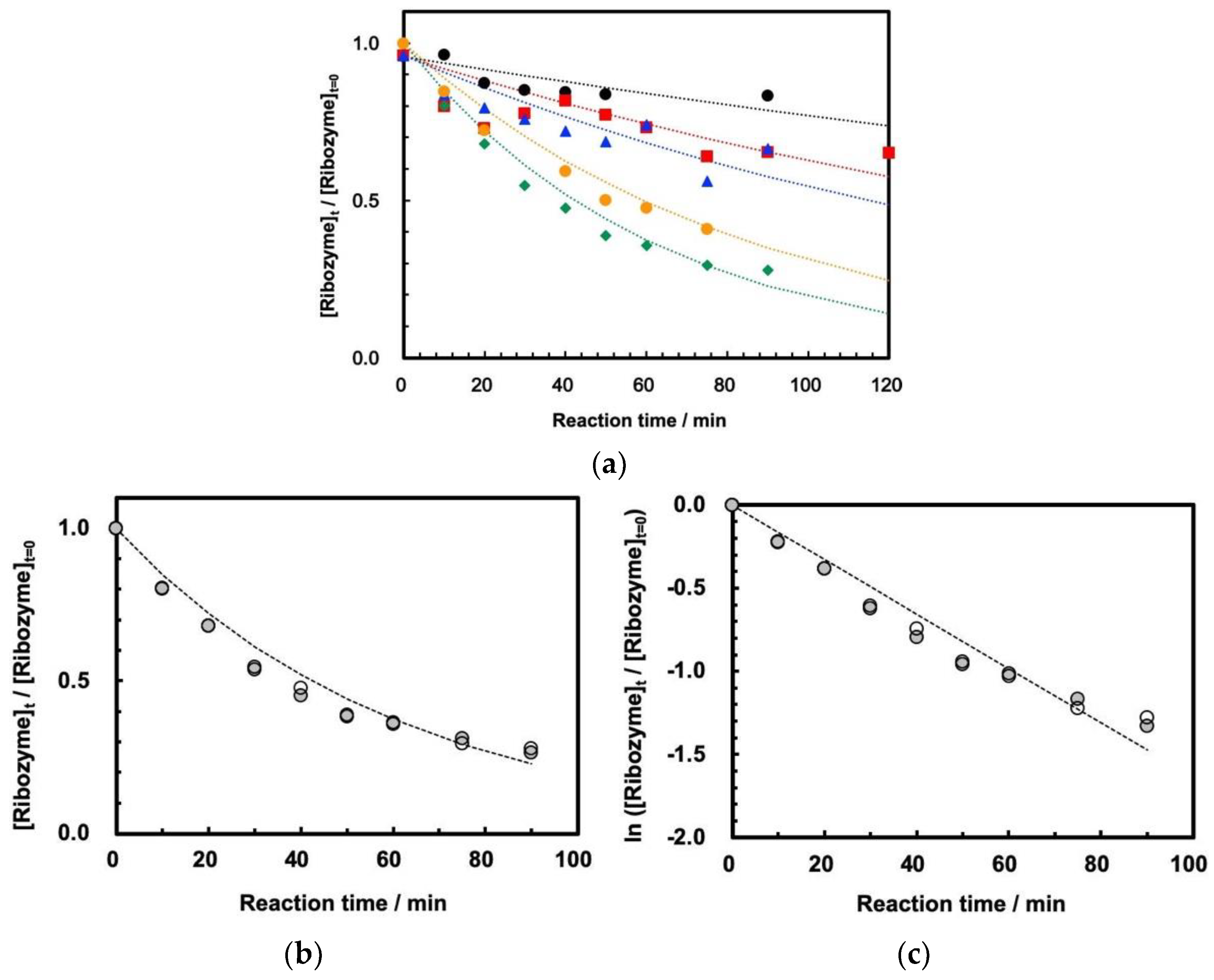
On the other hand, the measurements of self-cleavage of ASBVd(−):HHR were successfully measured for the first time at 30 MPa at temperatures of 45–65 °C by using HPR-30. The reaction curves at temperatures of 45–65 °C are shown in
Figure 4a. We confirmed that the reaction profile is well reproducible by duplicating the experiments at 60 °C (
Figure 4b). This indicates that ribozyme cleavage proceeded well inside the tubing reactor at 30 MPa at 45–65 °C, that the reaction was stopped completely by the addition of EDTA solution, and that evaporation of the sample mixture did not affect the cleavage reaction. The first-order kinetic plots show that the reaction basically follows first-order kinetics at 30 MPa (
Figure 4c), although the plots seem to diverge from the estimated first-order kinetics curve at longer reaction times. The first-order rate plots at 30 MPa at 45–65 °C are summarized in
Figure S2 (Supplementary Materials), and the rate constants are shown in
Table 1 for comparison with the rate constants at 0.1 MPa.
The present measurements using HPR-30 demonstrate that the HPR-30 system allows us to follow the ribozyme behavior at 30 MPa using a small amount of ribozyme (ca. 3.0 μg) and a small volume of sample solution (100 μL in sample loop volume). The measurements at high pressure for relatively long residence time up to 120 min were also successful, without leaking of the liquid from the lines of the system at 30 MPa. Furthermore, the present system is relatively simple to build, so that high-pressure measurements would be easily applicable in different fields of chemical and biochemical processes.
3.3. Influence of High Pressure on the Cleavage Behavior of ASBVd(−):HHR
Although there are some extensive studies regarding the enzymatic activities of protein enzymes at high pressures [
10,
11,
26,
27,
28,
29], high-pressure studies on ribozymes are scarce [
8,
10,
15]. Here, we confirmed that the cleavage reaction forming two components of 28 nt and 51 nt RNAs proceeded at both 0.1 and 30 MPa at 45–65 °C. This is consistent with our previous study, where ASBVd(−):HHR showed that the cleavage reaction proceeds in the temperature range of 10–70 °C [
15]. The reaction rate constants are relatively small as compared to other hammerhead ribozymes [
30,
31]. It was observed that random hydrolysis of the ribozyme proceeds in parallel with the cleavage reaction, and that the ratio of random hydrolysis to cleavage increases at temperatures over 65 °C. This is also consistent with our previous study [
19]. Although minor products other than 28 nt and 51 nt RNAs were observed by HPLC analysis corresponding to random hydrolysis, the cleavage reaction was dominant under our pressure and temperature conditions.
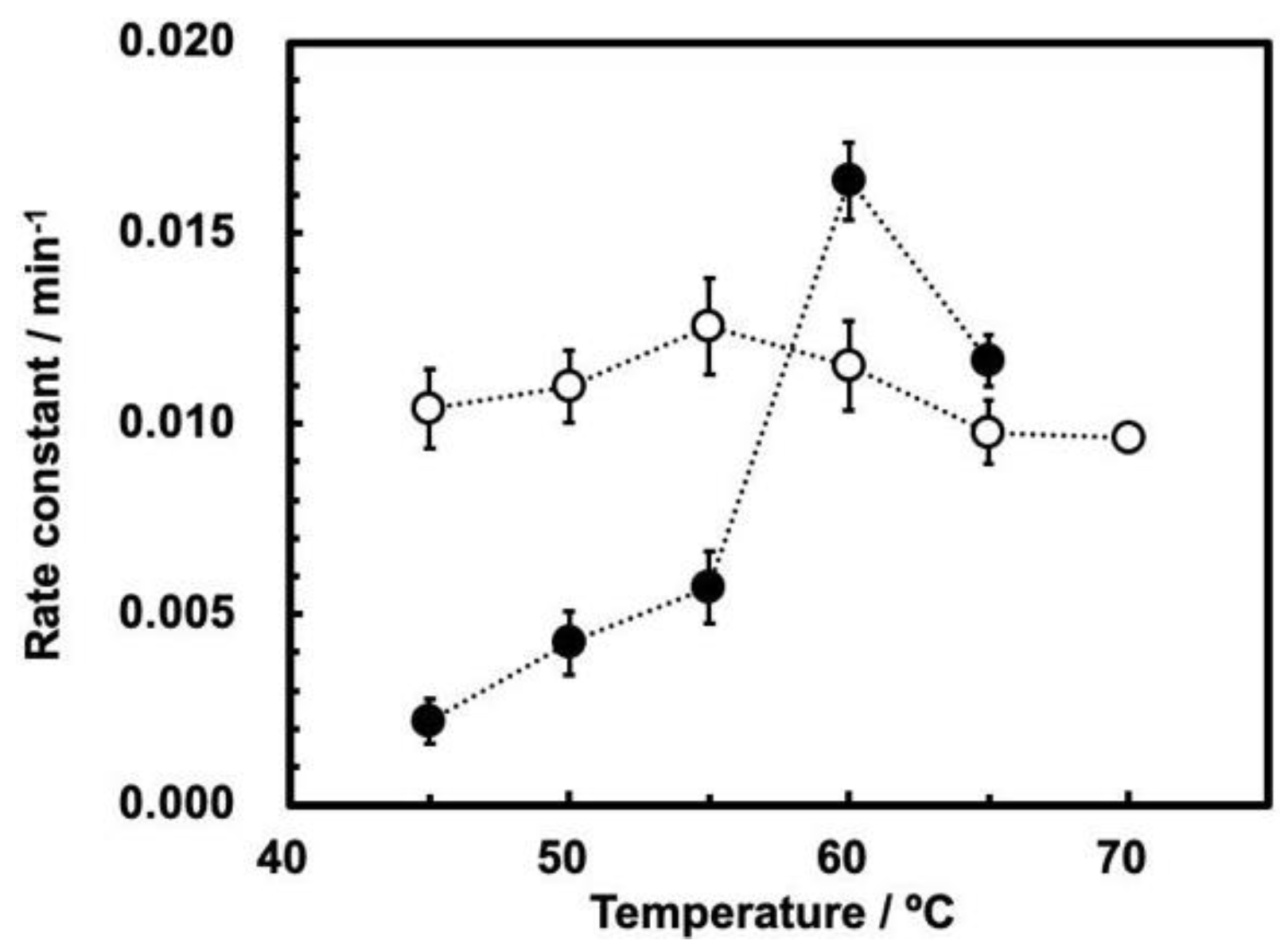
The profiles of the rate constants vs. temperature at 0.1 and 30 MPa (
Figure 5) suggest the following comments.
First, the rate constants of ribozyme cleavage are significantly smaller at 30 MPa than at 0.1 MPa at temperatures up to 55 °C; ribozyme cleavage is inhibited by a high pressure up to 55 °C. Second, the maximum rate constant was observed at 55 °C at 0.1 MPa, and at 60 °C at 30 MPa. This difference suggests that the cleavage of the ribozyme basically follows the same reaction mechanism to the original ASBVd [
15], while the maximum temperatures are somewhat different between 0.1 MPa and 30 MPa. Third, the influence of temperature at 0.1 MPa is moderate, as the ratio between rate constants at 55 °C and 45 °C is ca. 1.25 times for ASBVd(−):HHR. This is similar to the ratio between rate constants at 55 °C and 45 °C for ASBVd at the same pressure [
15]. In contrast, the rate constant at 30 MPa is more sensitive to temperature, as the rate constant at 60 °C is 12 times higher than that at 45 °C. The cleavage reaction rates at 30 MPa is actually higher than those at 0.1 MPa at 60–65 °C. This seems to be at odds with the results published in reference [
15]; as can be seen from
Figure 3a, which shows the effect of temperature on the reaction rates at different pressures, the rates consistently decrease with increasing pressure, and they never become greater (at 65 °C, the rate at 25 MPa is smaller than at 0.1 MPa). Furthermore, the temperature dependence for ASBVd(−) at higher pressures up to 200 MPa was moderate in comparison to the ribozyme [
15]. The ratio between the rate constants at 55 °C and at 45 °C is less than a factor of 3 at all pressures between 0.1–200 MPa.
In addition, as mentioned above, the cleavage reaction does not seem to perfectly fit first-order kinetics in the present study (
Figure 4c). It is known that the cleavage of a hammerhead ribozyme from Chrysanthemum chlorotic mottle viroid consists of a two-component reaction mechanism, including fast and slow reactions [
10]. According to our previous investigation, these are dependent on the interactions of three-dimensional structures due to surrounding sequences of the active center of the ribozyme part of the viroid [
15]. This is also consistent with the assumption that the associate formation prior to the self-cleavage becomes weak with increasing temperature [
32]. Thus, we assumed that the complex kinetics are due to conformational effects of those three-dimensional structures. Thus, the temperature- and pressure-related behaviors of the cleavage of ASBVd(−):HHR in the present study would suggest that the difference between 0.1 and 30 MPa is due to different conformational change of the ribozyme at 60 °C, where the absence of surrounding long sequences in the ribozyme weakens the core structure of the catalytic center at 30 MPa.
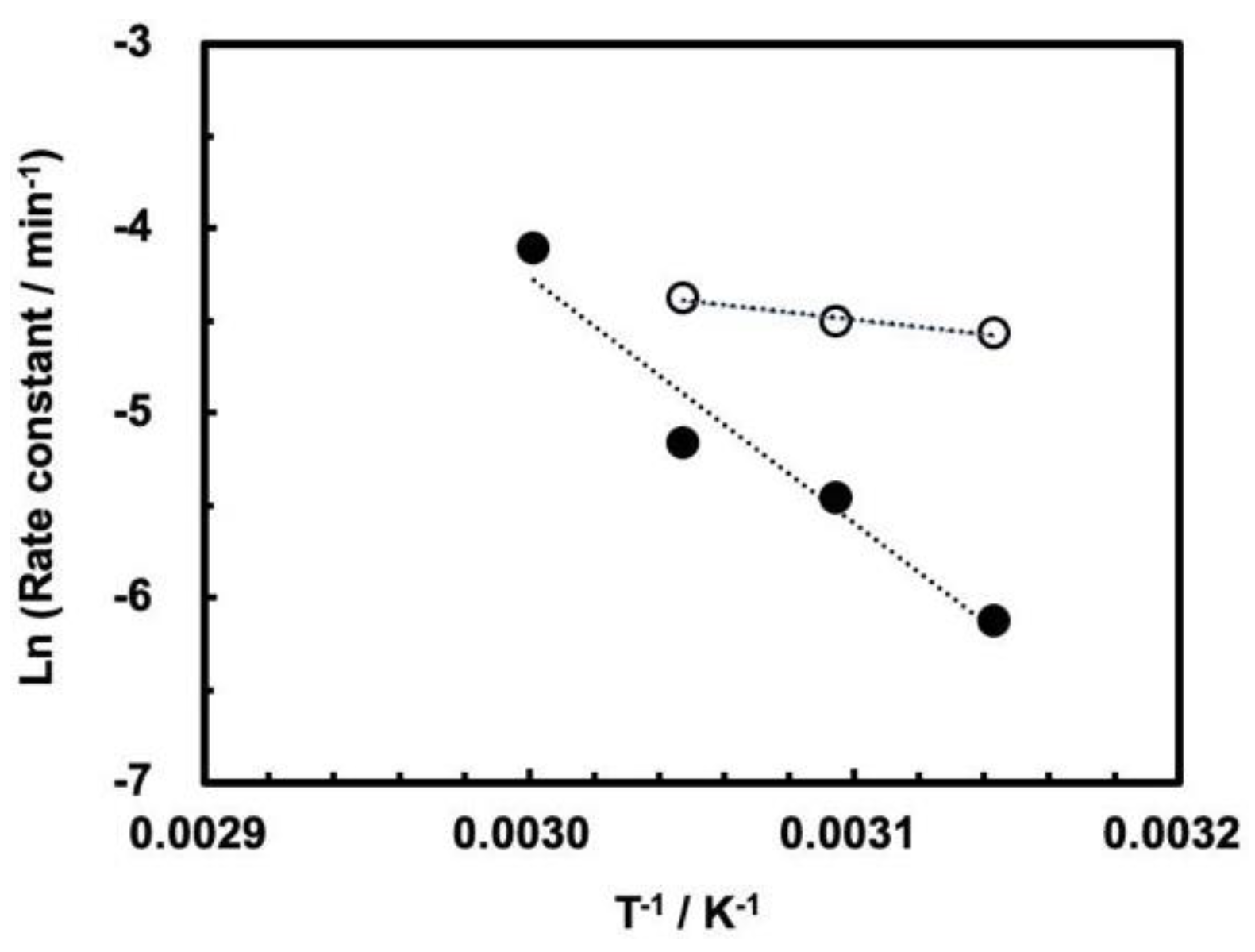
The cleavage reaction obeys the Arrhenius plots for the cleavage rate constants up to the maximum rate, that is, at 55 °C for 0.1 MPa and at 60 °C for 30 MPa, as shown in
Figure 6. The apparent activation energies in the temperature ranges up to the maximum rate have been calculated (
Table 2). Although we do not fully understand the origin of the trends shown in
Figure 5, we assume the following kinetic reasons on the basis of the apparent activation energies. First, the difference between 0.1 MPa and 30 MPa is large as compared to the case of ASBVd(−) viroid, where the behaviors of temperature dependence seems to be parallel for ASBVd(−) viroid at pressures of 0.1–200 MPa. Second, the fact that the activation energy at 30 MPa is fairly high suggests that the enzymatic function for self-cleavage at a specific position is less active at 30 MPa, so the activation energy is not efficiently reduced by the ribozyme. However, the presence of an optimum temperature indicates that the enzymatic function of the ribozyme is partially active at 30 MPa. In addition, this fact is consistent with our previous results showing that self-cleavage is affected by surrounding sequence of the ASBVd(−) viroid [
15,
33]. At the same time, we need to assume that a specific three-dimensional structure is partially maintained in order to ensure cleavage at the particular position -CG- between the 51st and 52nd nucleotide residues within the ribozyme.
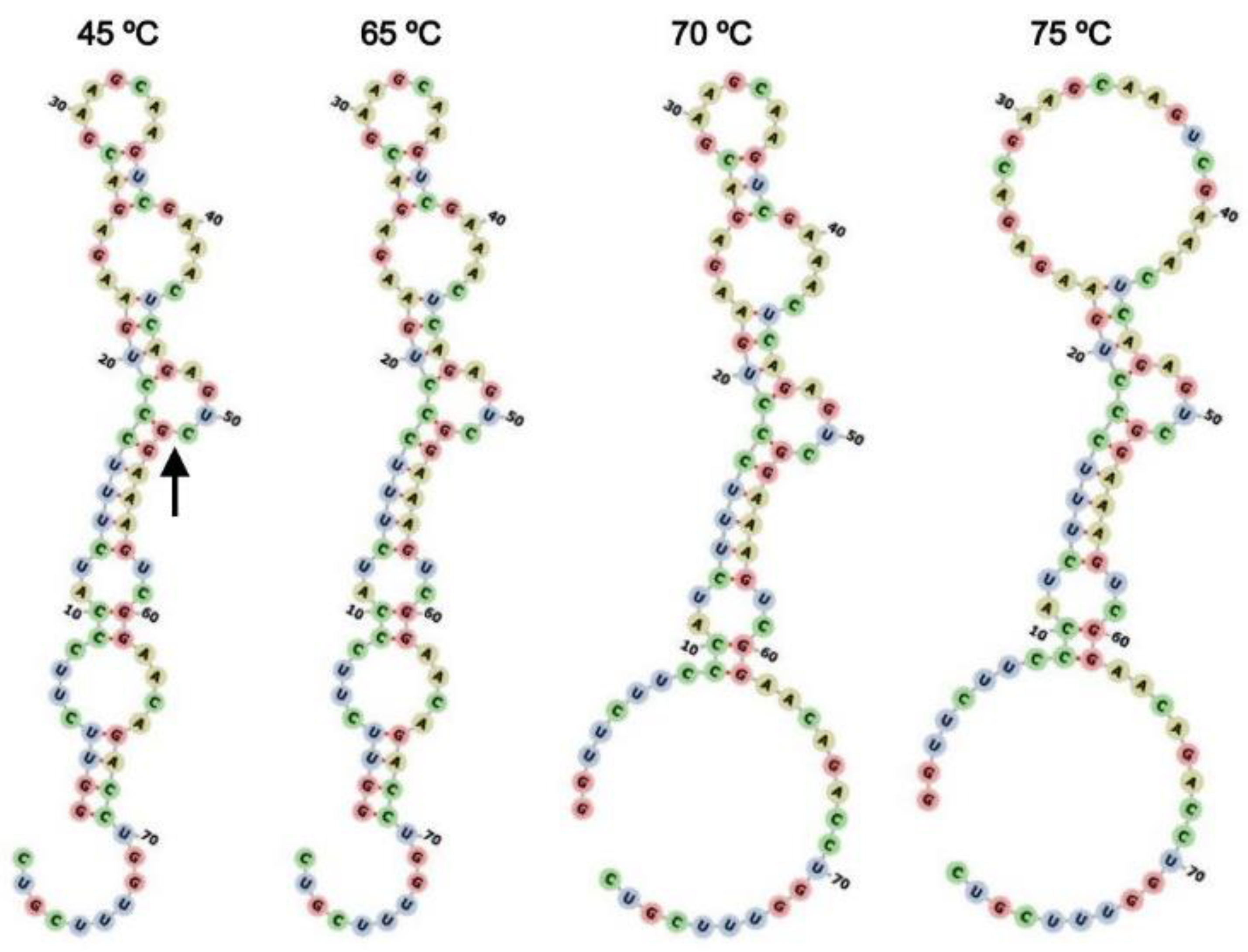
Modeling of secondary structures of the ribozyme (
Figure 7) indicates that the centroid secondary structure of ASBVd(−):HHR starts to change between 65 °C and 70 °C, but the estimation does not account for the pressure influence. In spite of this shortcoming, this is consistent with the optimum reaction rate observed at 55–60 °C for ASBVd(−):HHR at 0.1 and 30 MPa. Although the partial disappearance of the hydrogen bonding within the secondary structure gradually starts at 70–75 °C. These observations would reflect the fact that the cleavage of the ribozyme at 65 °C and 70 °C proceeded dominantly rather than the random hydrolysis of the ASBVd(−):HHR, where the ribozyme self-cleavage is still active at 65 and 70 °C.
Comparison of the rate constants at 0.1 MPa and 30 MPa demonstrates that the reaction rate decreases with increasing pressure at 55 °C and below, and the reaction rate increases with increasing pressure at 60 °C and 65 °C. The activation volume is positive at 55 °C and below and negative at 60 and 65 °C. The positive activation volume suggests that there is desolvation of water molecules at the activated complex, but the negative activation volume suggests that there is solvation by water molecules within the activated complex. The activation volume can be estimated from the data at 0.1 MP and 30 MPa, although the accuracy of calculation is poor due to only using two different pressures. The roughly estimated values of ΔV
‡ are 141 mL/mol at 45 °C, 87 mL/mol at 50 °C, 61 mL/min at 55 °C, −33 mL/mol at 60 °C, and −17 mL/mol at 65 °C. These values may suggest that a number of water molecules of one or higher contributes to the self-cleavage reaction since the mol volume of water is 18 mL/mol. The increase of the temperature to 60 °C would cause a change of conformation favoring the fixation of water molecules during the formation of the transition state, thus leading to a decrease of the reaction volume, down to negative values; this explains why, at high temperatures, there is an increase of the rate when the pressure increases. We are currently designing a high-pressure flow reactor adapted to run the reactions at pressures higher that 30 MPa so that a detailed analysis of the pressure dependence for ASBVd(−):HHR and inspection of hydration and dehydration [
34] can be carried out.
A pressure of 30 MPa appears frequently in submarine vent systems deep in the ocean, corresponding to 3000 m in depth. In addition, the length of ASBVd(−):HHR is moderate as compared to functional RNA in nature and in synthetic RNA by the in vitro selection technique [
35]. Thus, the fact that the cleavage reaction rate decreases with increasing pressure at temperatures up to 55 °C is useful for considering the compensation effect between pressure and temperature for primitive functional RNA molecules. In addition, the fact that the self-cleavage of ASBVd(−):HHR is fairly sensitive to pressures between 0.1 and 30 MPa suggests that pressure is an important factor for the selection and chemical evolution of RNA molecules in an RNA world compatible with the Hadean Earth environment. Finally, these data prove that our new high-pressure research tool would be very useful for investigating chemical evolution at high pressures at various temperatures since the system can be readily constructed from commercially available equipment and materials, as well as being suitable for a small volume of samples.


 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}