
5-HT3 Receptors in Rat Dorsal Root Ganglion Neurons: Ca2+ Entry and Modulation of Neurotransmitter Release


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Primary Neuronal Cultures
2.2. Expression of Human 5-HT3A and 5-HT3B in GH4C1 Cells
2.3. Electrophysiology
2.4. Ca2+ Measurements
2.5. Solutions and Chemicals
2.6. Data Analysis and Statistics
3. Results
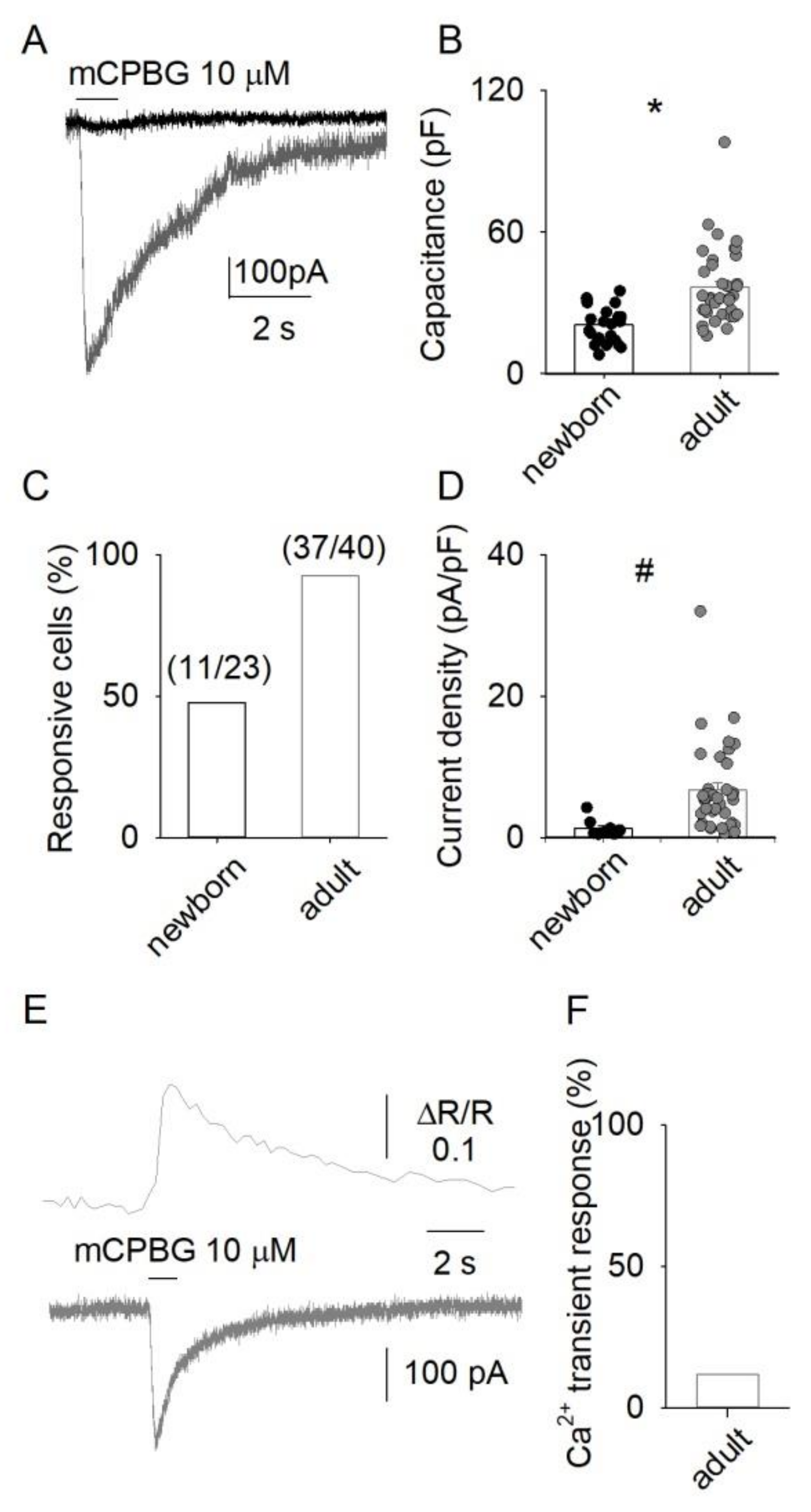
3.1. 5-HT3 Receptor-Mediated Responses in DRG Neurons
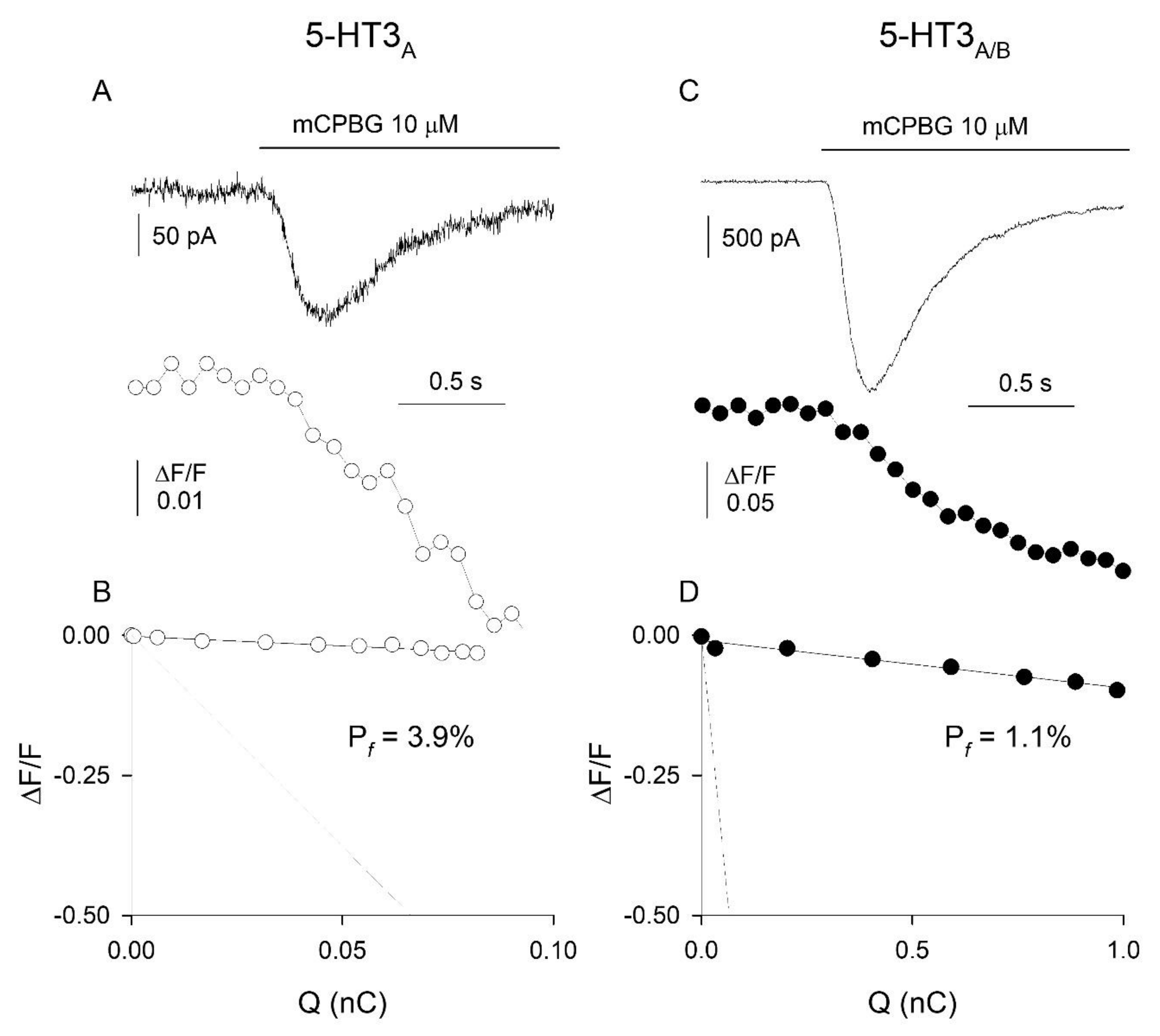
3.2. Ca2+ Permeability of Somatic 5-HT3Rs in Adult DRG Neurons
3.3. Modulatory Role of 5-HT3Rs in Neurotransmission
3.4. Modulatory Role of Ca2+ Entry through 5-HT3Rs
4. Discussion
4.1. Expression of 5-HT3Rs in DRG Neurons
4.2. Presynaptic 5-HT3Rs in DRG Neurons
4.3. Ca2+ Permeability of 5-HT3Rs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Richardson, B.P. Serotonin and nociception. Ann. N. Y. Acad. Sci. 1990, 600, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Dray, A. Inflammatory mediators of pain. Br. J. Anaesth. 1995, 75, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Sorkin, L.S.; McAdoo, D.J.; Willis, W.D. Raphe magnus stimulation-induced antinociception in the cat is associated with release of amino acids as well as serotonin in the lumbar dorsal horn. Brain Res. 1993, 618, 95–108. [Google Scholar] [CrossRef]
- Millan, M.J. Descending control of pain. Prog. Neurobiol. 2002, 66, 355–474. [Google Scholar] [CrossRef]
- De Kort, A.N.; Joosten, E.A.J.; Patijn, J.; Tibboel, D.; van den Hoogen, N.J. The development of descending serotonergic modulation of the spinal nociceptive network: A life span perspective. Pediatr. Res. 2022, 91, 1361–1369. [Google Scholar] [CrossRef]
- Nicholson, R.; Small, J.; Dixon, A.K.; Spanswick, D.; Lee, K. Serotonin receptor mRNA expression in rat dorsal root ganglion neurons. Neurosci. Lett. 2003, 337, 119–122. [Google Scholar] [CrossRef]
- Doak, G.J.; Sawynok, J. Formalin-induced nociceptive behavior and edema: Involvement of multiple peripheral 5-hydroxytryptamine receptor subtypes. Neuroscience 1997, 80, 939–949. [Google Scholar] [CrossRef]
- Robertson, B.; Bevan, S. Properties of 5-hydroxytryptamine3 receptor-gated currents in adult rat dorsal root ganglion neurones. Br. J. Pharmacol. 1991, 102, 272–276. [Google Scholar] [CrossRef] [Green Version]
- Todorovic, S.M.; Scroggs, R.S.; Anderson, E.G. Cationic modulation of 5-HT2 and 5-HT3 receptors in rat sensory neurons: The role of K+, Ca2+ and Mg2+. Brain Res. 1997, 765, 291–300. [Google Scholar] [CrossRef]
- Derkach, V.; Surprenant, A.; North, R.A. 5-HT3 receptors are membrane ion channels. Nature 1989, 339, 706–70942. [Google Scholar] [CrossRef]
- Maricq, A.V.; Peterson, A.S.; Brake, A.J.; Myers, R.M.; Julius, D. Primary structure and functional expression of the 5-HT3 receptor, a serotonin-gated ion channel. Science 1991, 254, 432–437. [Google Scholar] [CrossRef]
- Faerber, L.; Drechsler, S.; Ladenburger, S.; Gschaidmeier, H.; Fischer, W. The neuronal 5-HT3 receptor network after 20 years of research--evolving concepts in management of pain and inflammation. Eur. J. Pharmacol. 2007, 560, 1–8. [Google Scholar] [CrossRef]
- Kayser, V.; Elfassi, I.E.; Aubel, B.; Melfort, M.; Julius, D.; Gingrich, J.A.; Hamon, M.; Bourgoin, S. Mechanical, thermal and formalin-induced nociception is differentially altered in 5-HT1A−/−, 5-HT1B−/−, 5-HT2A−/−, 5-HT3A−/− and 5-HTT−/− knock-out male mice. Pain 2007, 130, 235–248. [Google Scholar] [CrossRef]
- Niesler, B.; Frank, B.; Kapeller, J.; Rappold, G.A. Cloning, physical mapping and expression analysis of the human 5-HT3 serotonin receptor-like genes HTR3C, HTR3D and HTR3E. Gene 2003, 310, 101–111. [Google Scholar] [CrossRef]
- Davies, P.A.; Pistis, M.; Hanna, M.C.; Peters, J.A.; Lambert, J.J.; Hales, T.G.; Kirkness, E.F. The 5-HT3B subunit is a major determinant of serotonin-receptor function. Nature 1999, 397, 359–363. [Google Scholar] [CrossRef]
- Dubin, A.E.; Huvar, R.; D’Andrea, M.R.; Pyati, J.; Zhu, J.Y.; Joy, K.C.; Wilson, S.J.; Galindo, J.E.; Glass, C.A.; Luo, L.; et al. The pharmacological and functional characteristics of the serotonin 5-HT(3A) receptor are specifically modified by a 5-HT(3B) receptor subunit. J. Biol. Chem. 1999, 274, 30799–30810. [Google Scholar] [CrossRef] [Green Version]
- Hapfelmeier, G.; Tredt, C.; Haseneder, R.; Zieglgansberger, W.; Eisensamer, B.; Rupprecht, R.; Rammes, G. Co-expression of the 5-HT3B serotonin receptor subunit alters the biophysics of the 5-HT3 receptor. Biophys. J. 2003, 84, 1720–1733. [Google Scholar] [CrossRef] [Green Version]
- Morales, M.; Wang, S.D. Differential composition of 5-hydroxytryptamine3 receptors synthesized in the rat CNS and peripheral nervous system. J. Neurosci. 2002, 22, 6732–6741. [Google Scholar] [CrossRef] [Green Version]
- Nayak, S.V.; Ronde, P.; Spier, A.D.; Lummis, S.C.; Nichols, R.A. Calcium changes induced by presynaptic 5-hydroxytryptamine-3 serotonin receptors on isolated terminals from various regions of the rat brain. Neuroscience 1999, 91, 107–117. [Google Scholar] [CrossRef]
- Rondé, P.; Nichols, R.A. High calcium permeability of serotonin 5-HT3 receptors on presynaptic nerve terminals from rat striatum. J. Neurochem. 1998, 70, 1094–1103. [Google Scholar] [CrossRef] [Green Version]
- Zazpe, A.; Artaiz, I.; Del Rio, J. Role of 5-HT3 receptors in basal and K+-evoked dopamine release from rat olfactory tubercle and striatal slices. Br. J. Pharmacol. 1994, 113, 968–972. [Google Scholar] [CrossRef] [Green Version]
- Glaum, S.R.; Brooks, P.A.; Spyer, K.M.; Miller, R.J. 5-Hydroxytryptamine-3 receptors modulate synaptic activity in the rat nucleus tractus solitarius in vitro. Brain Res. 1992, 589, 62–68. [Google Scholar] [CrossRef]
- Kawamata, T.; Omote, K.; Toriyabe, M.; Yamamoto, H.; Namiki, A. The activation of 5-HT3 receptors evokes GABA release in the spinal cord. Brain Res. 2003, 978, 250–255. [Google Scholar] [CrossRef]
- Koyama, S.; Matsumoto, N.; Kubo, C.; Akaike, N. Presynaptic 5-HT3 receptor-mediated modulation of synaptic GABA release in the mechanically dissociated rat amygdala neurons. J. Physiol. 2000, 529, 373–383. [Google Scholar] [CrossRef]
- Ropert, N.; Guy, N. Serotonin facilitates GABAergic transmission in the CA1 region of rat hippocampus in vitro. J. Physiol. 1991, 441, 121–136. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ramage, A.G.; Jordan, D. Presynaptic 5-HT3 receptors evoke an excitatory response in dorsal vagal preganglionic neurones in anaesthetized rats. J. Physiol. 1998, 509, 683–694. [Google Scholar] [CrossRef]
- Hicks, G.A.; Coldwell, J.R.; Schindler, M.; Ward, P.A.B.; Jenkins, D.; Lynn, P.A.; Humphrey, P.P.A.; Blackshaw, L.A. Excitation of rat colonic afferent fibres by 5-HT3 receptors. J. Physiol. 2002, 544, 861–869. [Google Scholar] [CrossRef]
- Morales, M.; McCollum, N.; Kirkness, E.F. 5-HT3-receptor subunits A and B are co-expressed in neurons of the dorsal root ganglion. J. Comp. Neurol. 2001, 438, 163–172. [Google Scholar] [CrossRef]
- Zeitz, K.P.; Guy, N.; Malmberg, A.B.; Dirajlal, S.; Martin, W.J.; Sun, L.; Bonhaus, D.W.; Stucky, C.L.; Julius, D.; Basbaum, A.I. The 5-HT3 subtype of serotonin receptor contributes to nociceptive processing via a novel subset of myelinated and unmyelinated nociceptors. J. Neurosci. 2002, 22, 1010–1019. [Google Scholar] [CrossRef] [Green Version]
- Kidd, E.J.; Laporte, A.M.; Langlois, X.; Fattaccini, C.-M.; Doyen, C.; Lombard, M.; Gozlan, H.; Hamon, M. 5-HT3 receptors in the rat central nervous system are mainly located on nerve fibres and terminals. Brain Res. 1993, 612, 289–298. [Google Scholar] [CrossRef]
- Khasabov, S.G.; Lopez-Garcia, J.A.; Asghar, A.U.; King, A.E. Modulation of afferent-evoked neurotransmission by 5-HT3 receptors in young rat dorsal horn neurones in vitro: A putative mechanism of 5-HT3 induced anti-nociception. Br. J. Pharmacol. 1999, 127, 843–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudomin, P.; Schmidt, R.F. Presynaptic inhibition in the vertebrate spinal cord revisited. Exp. Brain Res. 1999, 129, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, A.C.; Lummis, S.C.; Taylor, C.W. Ca2+ permeability of cloned and native 5-hydroxytryptamine type 3 receptors. Mol. Pharmacol. 1994, 46, 1120–1128. [Google Scholar] [PubMed]
- Yang, J. Ion permeation through 5-hydroxytryptamine-gated channels in neuroblastoma N18 cells. J. Gen. Physiol. 1990, 96, 1177–1198. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Neher, E. Calcium permeability of nicotinic acetylcholine receptor channels in bovine adrenal chromaffin cells. Pflug. Arch. 1993, 425, 511–517. [Google Scholar] [CrossRef]
- Egan, T.M.; Khakh, B.S. Contribution of calcium ions to P2X channel responses. J. Neurosci. 2004, 24, 3413–3420. [Google Scholar] [CrossRef] [Green Version]
- Noam, Y.; Wadman, W.J.; van Hooft, J.A. On the voltage-dependent Ca2+ block of serotonin 5-HT3 receptors: A critical role of intracellular phosphates. J. Physiol. 2008, 586, 3629–3638. [Google Scholar] [CrossRef]
- Fucile, S.; Sucapane, A.; Eusebi, F. Ca2+ permeability of nicotinic acetylcholine receptors from rat dorsal root ganglion neurones. J. Physiol. 2005, 565, 219–228. [Google Scholar] [CrossRef]
- Kilpatrick, G.J.; Bunce, K.T.; Tyers, M.B. 5-HT3 receptors. Med. Res. Rev. 1990, 10, 441–475. [Google Scholar] [CrossRef]
- Fucile, S. Ca2+ permeability of nicotinic acetylcholine receptors. Cell Calcium 2004, 35, 1–8. [Google Scholar] [CrossRef]
- Gu, J.G.; MacDermott, A.B. Activation of ATP P2X receptors elicits glutamate release from sensory neuron synapses. Nature 1997, 389, 749–753. [Google Scholar] [CrossRef]
- Bleakman, D.; Bowman, D.; Bath, C.P.; Brust, P.F.; Johnson, E.C.; Deal, C.R.; Miller, R.J.; Ellis, S.B.; Harpold, M.M.; Hans, M.; et al. Characteristics of a human N-type calcium channel expressed in HEK293 cells. Neuropharmacology 1995, 34, 753–765. [Google Scholar] [CrossRef]
- Fucile, S. The distribution of charged amino acid residues and the Ca2+ permeability of nicotinic acetylcholine receptors: A predictive model. Front. Mol. Neurosci. 2017, 10, 155. [Google Scholar] [CrossRef] [Green Version]
- Burnashev, N. Calcium permeability of ligand-gated channels. Cell Calcium 1998, 24, 325–332. [Google Scholar] [CrossRef]
- Linhart, O.; Obreja, O.; Kress, M. The inflammatory mediators serotonin, prostaglandin E2 and bradykinin evoke calcium influx in rat sensory neurons. Neuroscience 2003, 118, 69–74. [Google Scholar] [CrossRef]
- Liu, X.Y.; Wu, S.X.; Wang, Y.Y.; Wang, W.; Zhou, L.; Li, Y.Q. Changes of 5-HT receptor subtype mRNAs in rat dorsal root ganglion by bee venom-induced inflammatory pain. Neurosci. Lett. 2005, 375, 42–46. [Google Scholar] [CrossRef]
- Giordano, J.; Dyche, J. Differential analgesic actions of serotonin 5-HT3 receptor antagonists in the mouse. Neuropharmacology 1989, 28, 423–427. [Google Scholar] [CrossRef]
- Giordano, J.; Rogers, L.V. Peripherally administered serotonin 5-HT3 receptor antagonists reduce inflammatory pain in rats. Eur. J. Pharmacol. 1989, 170, 83–86. [Google Scholar] [CrossRef]
- Orwin, J.M.; Fozard, J.R. Blockade of the flare response to intradermal 5-hydroxytryptamine in man by MDL 72.222, a selective antagonist at neuronal 5-hydroxytryptamine receptors. Eur. J. Clin. Pharmacol. 1986, 30, 209–212. [Google Scholar] [CrossRef]
- Kia, H.K.; Miquel, M.C.; McKernan, R.M.; Laporte, A.-M.; Lombard, M.-C.; Bourgoin, S.; Hamon, M.; Vergé, D. Localization of 5-HT3 receptors in the rat spinal cord: Immunohistochemistry and in situ hybridization. Neuroreport 1995, 6, 257–261. [Google Scholar] [CrossRef]
- Inoue, A.; Hashimoto, T.; Hide, I.; Nishio, H.; Nakata, Y. 5-Hydroxytryptamine-facilitated release of substance P from rat spinal cord slices is mediated by nitric oxide and cyclic GMP. J. Neurochem. 1997, 68, 128–133. [Google Scholar] [CrossRef]
- Saria, A.; Javorsky, F.; Humpel, C.; Gamse, R. 5-HT3 receptor antagonists inhibit sensory neuropeptide release from the rat spinal cord. Neuroreport 1990, 1, 104–106. [Google Scholar] [CrossRef]
- Conte, D.; Legg, E.D.; McCourt, A.C.; Silajdzic, E.; Nagy, G.G.; Maxwell, D.J. Transmitter content, origins and connections of axons in the spinal cord that possess the serotonin (5-hydroxytryptamine) 3 receptor. Neuroscience 2005, 134, 165–173. [Google Scholar] [CrossRef]
- Morales, M.; Battenberg, E.; Bloom, F.E. Distribution of neurons expressing immunoreactivity for the 5-HT3 receptor subtype in the rat brain and spinal cord. J. Comp. Neurol. 1998, 402, 385–401. [Google Scholar] [CrossRef]
- Tecott, L.H.; Maricq, A.V.; Julius, D. Nervous system distribution of the serotonin 5-HT3 receptor mRNA. Proc. Natl. Acad. Sci. USA 1993, 90, 1430–1434. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.J.; Mokler, D.J.; Luebke, J.I. Calcium influx through presynaptic 5-HT3 receptors facilitates GABA release in the hippocampus: In vitro slice and synaptosome studies. Neuroscience 2004, 129, 703–718. [Google Scholar] [CrossRef]
- Gilon, P.; Yakel, J.L. Activation of 5-HT3 receptors expressed in Xenopus oocytes does not increase cytoplasmic Ca2+ levels. Recept. Channels 1995, 3, 83–88. [Google Scholar]
- Yakel, J.L.; Shao, X.M.; Jackson, M.B. The selectivity of the channel coupled to the 5-HT3 receptor. Brain Res. 1990, 533, 46–52. [Google Scholar] [CrossRef]
- Brown, A.M.; Hope, A.G.; Lambert, J.J.; Peters, J.A. Ion permeation and conduction in a human recombinant 5-HT3 receptor subunit (h5-HT3A). J. Physiol. 1998, 507, 653–665. [Google Scholar] [CrossRef]
- Schmidt, B.J.; Jordan, L.M. The role of serotonin in reflex modulation and locomotor rhythm production in the mammalian spinal cord. Brain Res. Bull. 2000, 53, 689–710. [Google Scholar] [CrossRef]
- Oatway, M.A.; Chen, Y.; Weaver, L.C. The 5-HT3 receptor facilitates at-level mechanical allodynia following spinal cord injury. Pain 2004, 110, 259–268. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinello, K.; Sucapane, A.; Fucile, S. 5-HT3 Receptors in Rat Dorsal Root Ganglion Neurons: Ca2+ Entry and Modulation of Neurotransmitter Release. Life 2022, 12, 1178. https://doi.org/10.3390/life12081178
Martinello K, Sucapane A, Fucile S. 5-HT3 Receptors in Rat Dorsal Root Ganglion Neurons: Ca2+ Entry and Modulation of Neurotransmitter Release. Life. 2022; 12(8):1178. https://doi.org/10.3390/life12081178
Chicago/Turabian StyleMartinello, Katiuscia, Antonietta Sucapane, and Sergio Fucile. 2022. "5-HT3 Receptors in Rat Dorsal Root Ganglion Neurons: Ca2+ Entry and Modulation of Neurotransmitter Release" Life 12, no. 8: 1178. https://doi.org/10.3390/life12081178