

Blocking of Caspases Exerts Anti-Inflammatory Effects on Periodontal Cells

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Murine ST2 Cells, Human Gingival Fibroblasts and HSC2 Oral Squamous Carcinoma Cells
2.2. Isolation and Culture of Murine Bone Marrow-Derived Macrophages and RAW264.7 Cells
2.3. qRT-PCR Analysis and Immunoassay
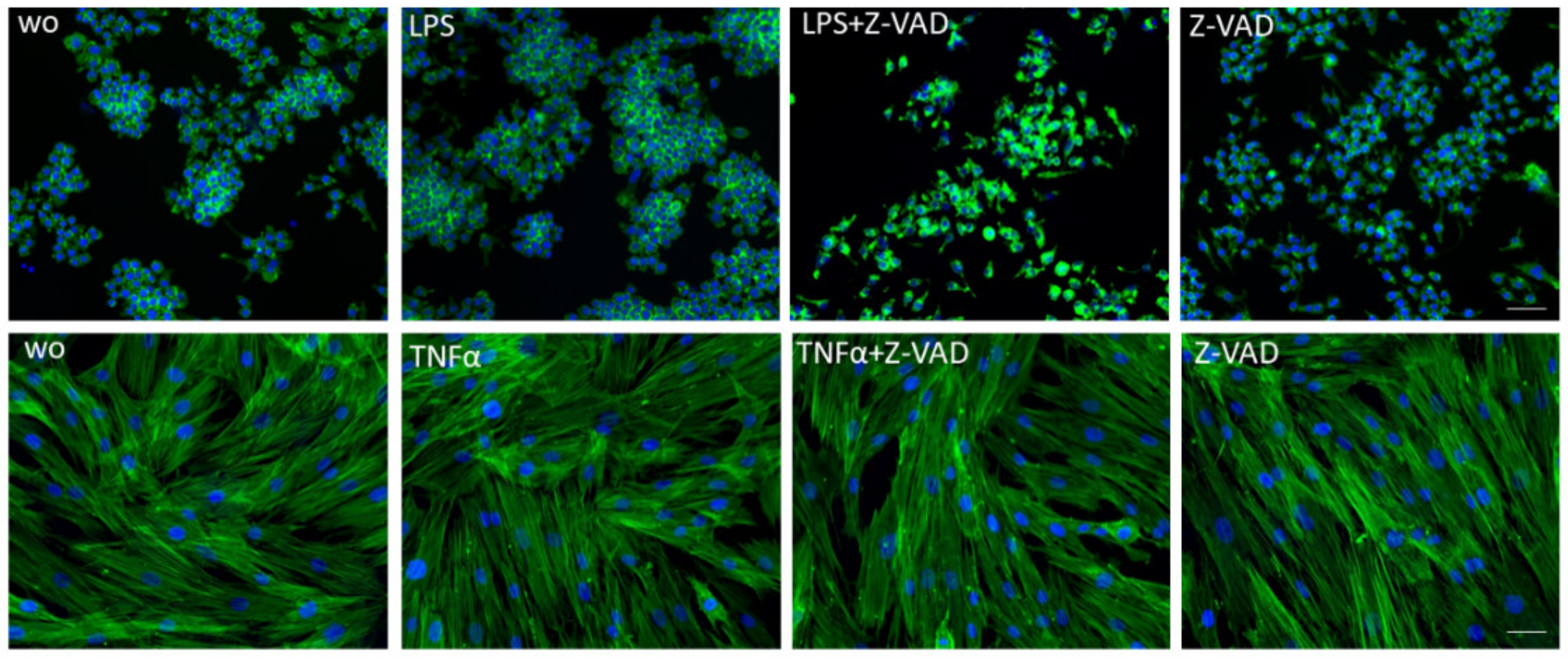
2.4. Fluorescence Microscopy
2.5. Caspase-8 Activity Assay
2.6. Statistical Analysis
3. Results
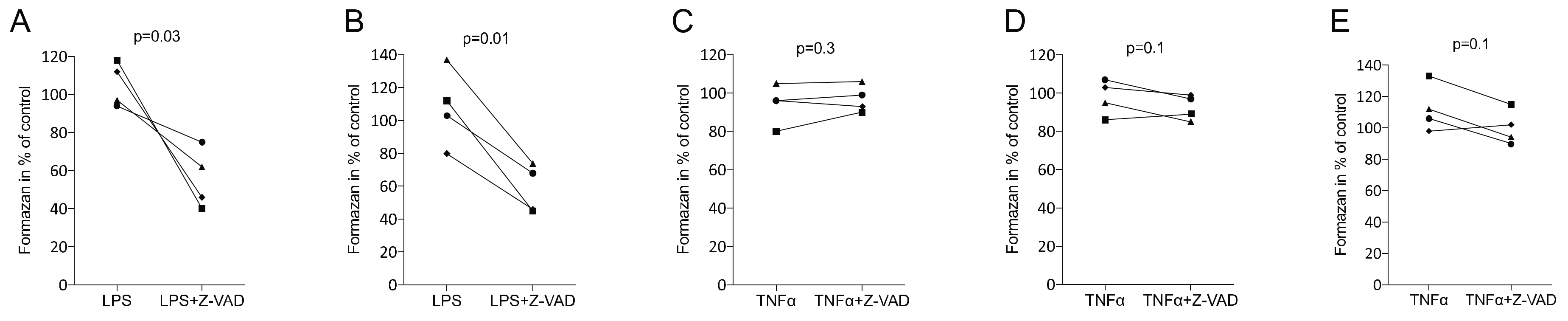
3.1. Z-VAD Decreased the Metabolic Activity of Stimulated Macrophages but Not of Mesenchymal and Epithelial Cells
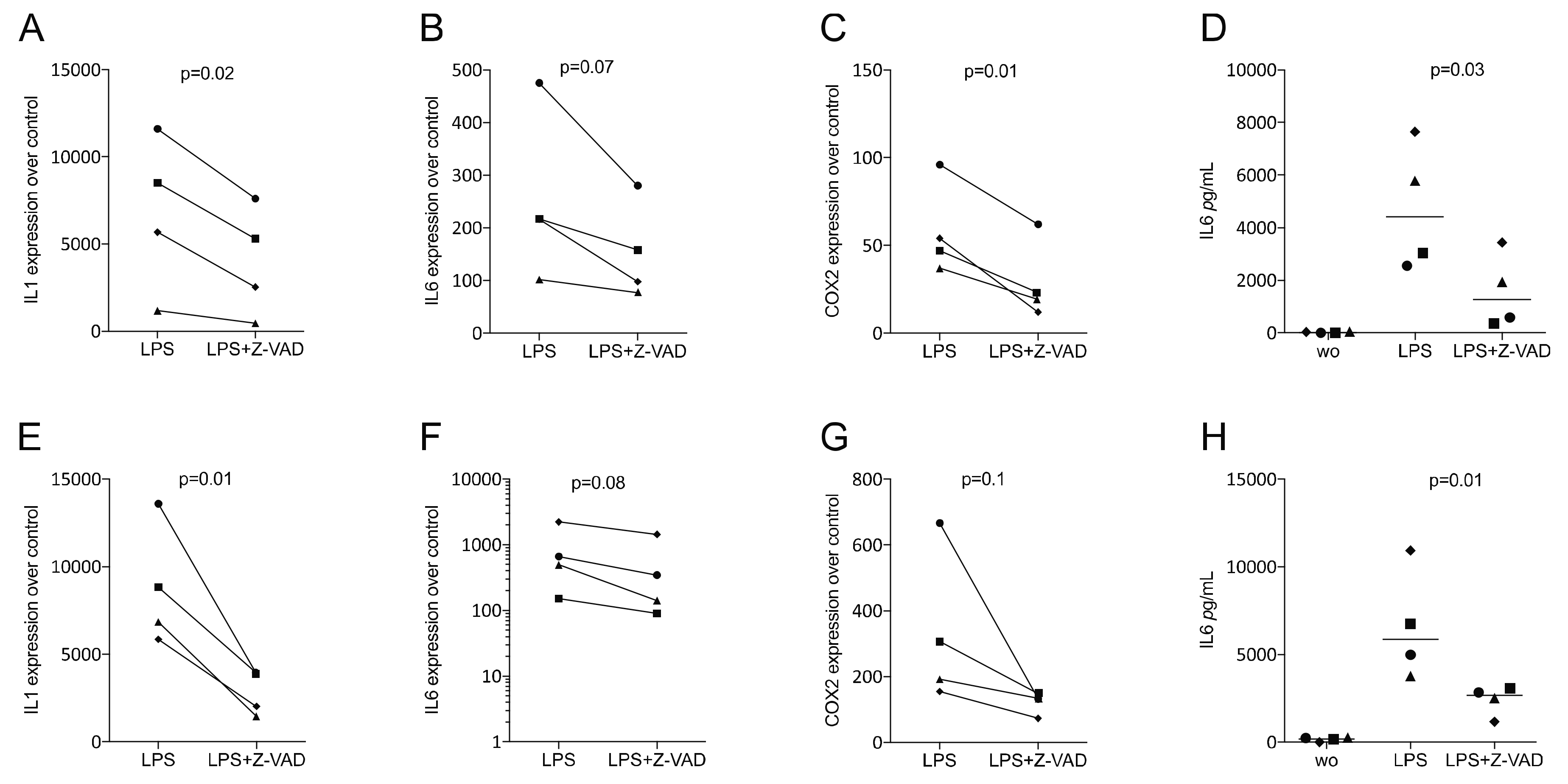
3.2. Z-VAD Decreased the Inflammatory Response of LPS-Stimulated RAW 264.7 Cells and Primary Macrophages
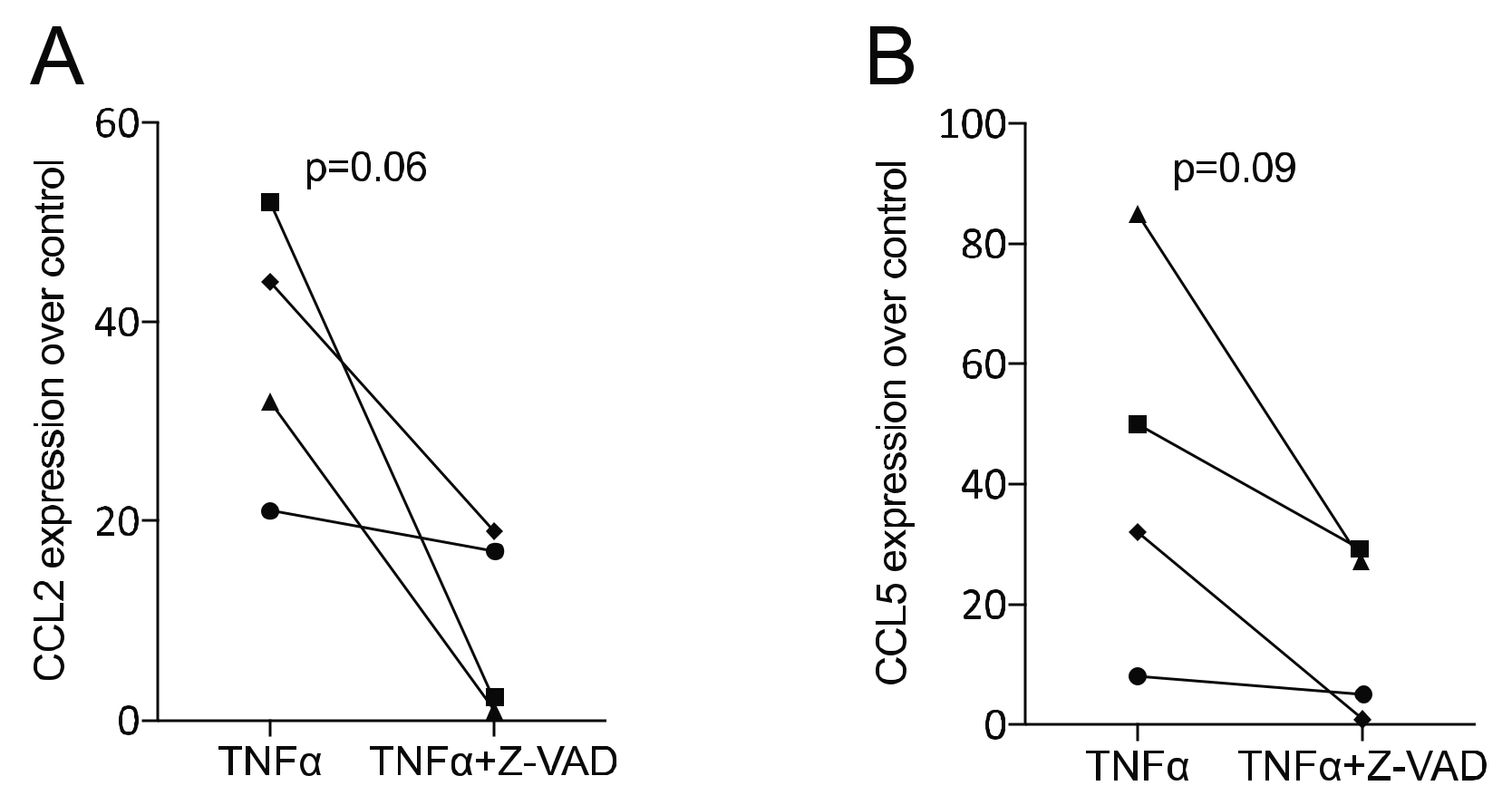
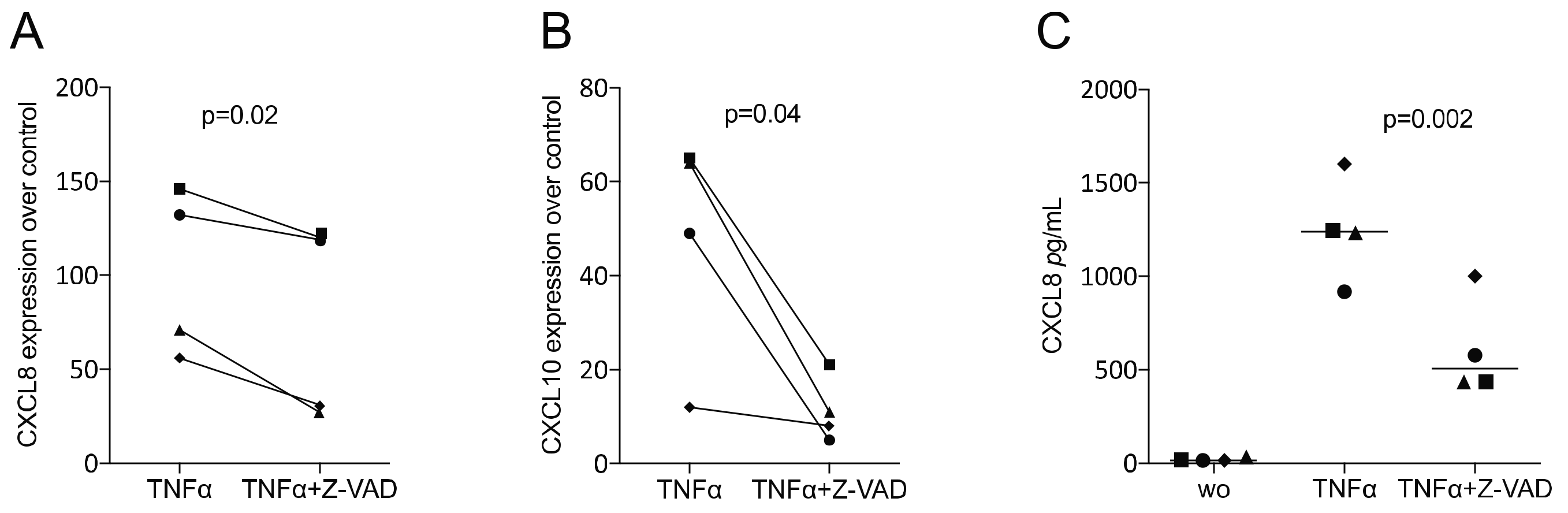
3.3. Z-VAD Decreased the Inflammatory Response of TNFα-Stimulated ST2 Mesenchymal Cells and Gingival Fibroblasts
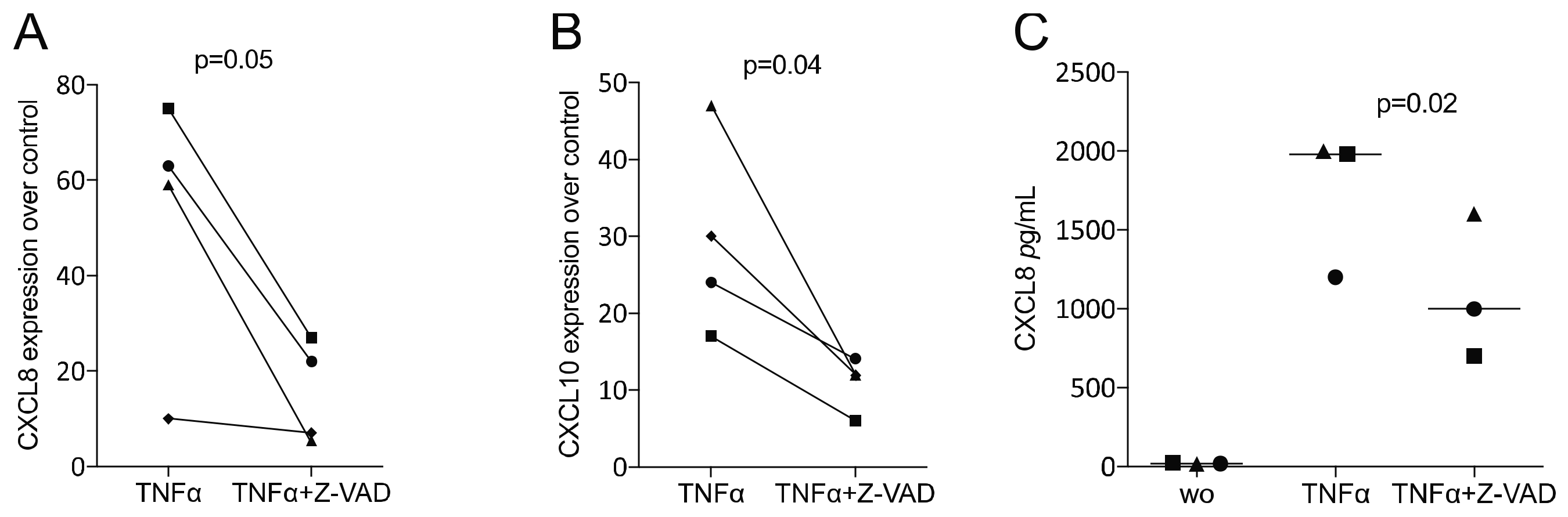
3.4. Z-VAD Decreased the Inflammatory Response of TNFα-Stimulated HSC2 Oral Squamous Carcinoma Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kinane, D.F.; Stathopoulou, P.G.; Papapanou, P.N. Periodontal diseases. Nat. Rev. Dis. Primers 2017, 3, 17038. [Google Scholar] [CrossRef] [PubMed]
- Pihlstrom, B.L.; Michalowicz, B.S.; Johnson, N.W. Periodontal diseases. Lancet 2005, 366, 1809–1820. [Google Scholar] [CrossRef] [Green Version]
- Song, B.; Zhang, Y.L.; Chen, L.J.; Zhou, T.; Huang, W.K.; Zhou, X.; Shao, L.Q. The role of Toll-like receptors in periodontitis. Oral. Dis. 2017, 23, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A.; Chawla, A.; Pollard, J.W. Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–455. [Google Scholar] [CrossRef]
- Panahipour, L.; Tabatabaei, A.A.; Gruber, R. Hypoallergenic infant formula lacks transforming growth factor beta activity and has a lower anti-inflammatory activity than regular infant formula. J. Dairy. Sci. 2020, 103, 6771–6781. [Google Scholar] [CrossRef] [PubMed]
- Ba, X.; Bacsi, A.; Luo, J.; Aguilera-Aguirre, L.; Zeng, X.; Radak, Z.; Brasier, A.R.; Boldogh, I. 8-oxoguanine DNA glycosylase-1 augments proinflammatory gene expression by facilitating the recruitment of site-specific transcription factors. J. Immunol. 2014, 192, 2384–2394. [Google Scholar] [CrossRef]
- Wilson, M.; Reddi, K.; Henderson, B. Cytokine-inducing components of periodontopathogenic bacteria. J. Periodontal Res. 1996, 31, 393–407. [Google Scholar] [CrossRef]
- Panahipour, L.; Nasserzare, S.; Amer, Z.; Brucke, F.; Stahli, A.; Kreissl, A.; Haiden, N.; Gruber, R. The anti-inflammatory effect of milk and dairy products on periodontal cells: An in vitro approach. Clin. Oral Investig. 2019, 23, 1959–1966. [Google Scholar] [CrossRef] [Green Version]
- Ohta, K.; Shigeishi, H.; Taki, M.; Nishi, H.; Higashikawa, K.; Takechi, M.; Kamata, N. Regulation of CXCL9/10/11 in oral keratinocytes and fibroblasts. J. Dent. Res. 2008, 87, 1160–1165. [Google Scholar] [CrossRef]
- Hosokawa, I.; Hosokawa, Y.; Ozaki, K.; Nakae, H.; Matsuo, T. Adrenomedullin suppresses tumour necrosis factor alpha-induced CXC chemokine ligand 10 production by human gingival fibroblasts. Clin. Exp. Immunol. 2008, 152, 568–575. [Google Scholar] [CrossRef]
- Franco, C.; Patricia, H.R.; Timo, S.; Claudia, B.; Marcela, H. Matrix Metalloproteinases as Regulators of Periodontal Inflammation. Int. J. Mol. Sci. 2017, 18, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maciejczyk, M.; Pietrzykowska, A.; Zalewska, A.; Knas, M.; Daniszewska, I. The Significance of Matrix Metalloproteinases in Oral Diseases. Adv. Clin. Exp. Med. 2016, 25, 383–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, R. Osteoimmunology: Inflammatory osteolysis and regeneration of the alveolar bone. J. Clin. Periodontol. 2019, 46 (Suppl. 21), 52–69. [Google Scholar] [CrossRef] [Green Version]
- Hienz, S.A.; Paliwal, S.; Ivanovski, S. Mechanisms of Bone Resorption in Periodontitis. J. Immunol. Res. 2015, 2015, 615486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, R.C.; Cullen, S.P.; Martin, S.J. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell. Biol. 2008, 9, 231–241. [Google Scholar] [CrossRef]
- Sordi, M.B.; Magini, R.S.; Panahipour, L.; Gruber, R. Pyroptosis-Mediated Periodontal Disease. Int. J. Mol. Sci. 2021, 23, 372. [Google Scholar] [CrossRef]
- Bergsbaken, T.; Fink, S.L.; Cookson, B.T. Pyroptosis: Host cell death and inflammation. Nat. Rev. Microbiol. 2009, 7, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Weinlich, R.; Oberst, A.; Beere, H.M.; Green, D.R. Necroptosis in development, inflammation and disease. Nat. Rev. Mol. Cell. Biol. 2017, 18, 127–136. [Google Scholar] [CrossRef]
- Kesavardhana, S.; Malireddi, R.K.S.; Kanneganti, T.D. Caspases in Cell Death, Inflammation, and Pyroptosis. Annu. Rev. Immunol. 2020, 38, 567–595. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Li, J. Caspase blockade induces RIP3-mediated programmed necrosis in Toll-like receptor-activated microglia. Cell Death Dis. 2013, 4, e716. [Google Scholar] [CrossRef] [Green Version]
- Bantel, H.; Beikler, T.; Flemmig, T.F.; Schulze-Osthoff, K. Caspase activation is involved in chronic periodontitis. FEBS Lett. 2005, 579, 5559–5564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, F.; Shu, R.; Xie, Y. The expression of NLRP3, NLRP1 and AIM2 in the gingival tissue of periodontitis patients: RT-PCR study and immunohistochemistry. Arch. Oral Biol. 2015, 60, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Bostanci, N.; Emingil, G.; Saygan, B.; Turkoglu, O.; Atilla, G.; Curtis, M.A.; Belibasakis, G.N. Expression and regulation of the NALP3 inflammasome complex in periodontal diseases. Clin. Exp. Immunol. 2009, 157, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Li, J.; Su, W.; Zhao, S.; Li, H.; Lei, L. Loss of periodontal ligament fibroblasts by RIPK3-MLKL-mediated necroptosis in the progress of chronic periodontitis. Sci. Rep. 2019, 9, 2902. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ke, X.; Yan, F.; Lei, L.; Li, H. Necroptosis in the periodontal homeostasis: Signals emanating from dying cells. Oral Dis. 2018, 24, 900–907. [Google Scholar] [CrossRef]
- Li, X.; Yao, X.; Zhu, Y.; Zhang, H.; Wang, H.; Ma, Q.; Yan, F.; Yang, Y.; Zhang, J.; Shi, H.; et al. The Caspase Inhibitor Z-VAD-FMK Alleviates Endotoxic Shock via Inducing Macrophages Necroptosis and Promoting MDSCs-Mediated Inhibition of Macrophages Activation. Front. Immunol. 2019, 10, 1824. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.O.; Ono, K.; Han, J. Apoptosis by pan-caspase inhibitors in lipopolysaccharide-activated macrophages. Am. J. Physiol. Lung Cell. Mol. Physiol. 2001, 281, L1095–L1105. [Google Scholar] [CrossRef]
- Kim, S.O.; Han, J. Pan-caspase inhibitor zVAD enhances cell death in RAW246.7 macrophages. J. Endotoxin Res. 2001, 7, 292–296. [Google Scholar] [CrossRef]
- Schwarze, U.Y.; Strauss, F.J.; Gruber, R. Caspase inhibitor attenuates the shape changes in the alveolar ridge following tooth extraction: A pilot study in rats. J. Periodontal Res. 2021, 56, 101–107. [Google Scholar] [CrossRef]
- Homsi, E.; Janino, P.; de Faria, J.B. Role of caspases on cell death, inflammation, and cell cycle in glycerol-induced acute renal failure. Kidney Int. 2006, 69, 1385–1392. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Johnson, T.S.; Haylor, J.L.; Wagner, B.; Watson, P.F.; El Kossi, M.M.; Furness, P.N.; El Nahas, A.M. Effects of caspase inhibition on the progression of experimental glomerulonephritis. Kidney Int. 2003, 63, 2050–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandri, B.; Nishioka, T.; Heimann, A.; Bullock, R.M.; Kempski, O. Caspase-dependent cell death involved in brain damage after acute subdural hematoma in rats. Brain Res. 2006, 1111, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Eipel, C.; Bordel, R.; Nickels, R.M.; Menger, M.D.; Vollmar, B. Impact of leukocytes and platelets in mediating hepatocyte apoptosis in a rat model of systemic endotoxemia. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G769–G776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhani, S.; Zhao, Y.; Zhivotovsky, B. A long way to go: Caspase inhibitors in clinical use. Cell Death Dis. 2021, 12, 949. [Google Scholar] [CrossRef]
- Park, J.; Kim, H.D.; Lee, S.H.; Kwak, C.H.; Chang, Y.C.; Lee, Y.C.; Chung, T.W.; Magae, J.; Kim, C.H. Ascochlorin induces caspase-independent necroptosis in LPS-stimulated RAW 264.7 macrophages. J. Ethnopharmacol. 2019, 239, 111898. [Google Scholar] [CrossRef] [PubMed]
- Burns, E.; Bachrach, G.; Shapira, L.; Nussbaum, G. Cutting Edge: TLR2 is required for the innate response to Porphyromonas gingivalis: Activation leads to bacterial persistence and TLR2 deficiency attenuates induced alveolar bone resorption. J. Immunol. 2006, 177, 8296–8300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulos, G.; Weinberg, E.O.; Massari, P.; Gibson, F.C., 3rd; Wetzler, L.M.; Morgan, E.F.; Genco, C.A. Macrophage-specific TLR2 signaling mediates pathogen-induced TNF-dependent inflammatory oral bone loss. J. Immunol. 2013, 190, 1148–1157. [Google Scholar] [CrossRef]
- Koike, A.; Hanatani, M.; Fujimori, K. Pan-caspase inhibitors induce necroptosis via ROS-mediated activation of mixed lineage kinase domain-like protein and p38 in classically activated macrophages. Exp. Cell Res. 2019, 380, 171–179. [Google Scholar] [CrossRef]
- Saeed, W.K.; Jun, D.W.; Jang, K.; Oh, J.H.; Chae, Y.J.; Lee, J.S.; Koh, D.H.; Kang, H.T. Decrease in fat de novo synthesis and chemokine ligand expression in non-alcoholic fatty liver disease caused by inhibition of mixed lineage kinase domain-like pseudokinase. J. Gastroenterol. Hepatol. 2019, 34, 2206–2218. [Google Scholar] [CrossRef]
- Zelic, M.; Roderick, J.E.; O’Donnell, J.A.; Lehman, J.; Lim, S.E.; Janardhan, H.P.; Trivedi, C.M.; Pasparakis, M.; Kelliher, M.A. RIP kinase 1-dependent endothelial necroptosis underlies systemic inflammatory response syndrome. J. Clin. Investig. 2018, 128, 2064–2075. [Google Scholar] [CrossRef]
- Vercammen, D.; Beyaert, R.; Denecker, G.; Goossens, V.; Van Loo, G.; Declercq, W.; Grooten, J.; Fiers, W.; Vandenabeele, P. Inhibition of caspases increases the sensitivity of L929 cells to necrosis mediated by tumor necrosis factor. J. Exp. Med. 1998, 187, 1477–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Chen, R.; Xu, Y.; Xia, C. PLCgamma1 inhibition combined with inhibition of apoptosis and necroptosis increases cartilage matrix synthesis in IL-1beta-treated rat chondrocytes. FEBS Open Bio 2021, 11, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Zhang, X.; Liu, N.; Tang, L.; Peng, C.; Chen, X. Pyroptosis: Mechanisms and diseases. Signal Transduct. Target. Ther. 2021, 6, 128. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panahipour, L.; Cervantes, L.C.C.; Oladzad Abbasabadi, A.; Sordi, M.B.; Kargarpour, Z.; Gruber, R. Blocking of Caspases Exerts Anti-Inflammatory Effects on Periodontal Cells. Life 2022, 12, 1045. https://doi.org/10.3390/life12071045
Panahipour L, Cervantes LCC, Oladzad Abbasabadi A, Sordi MB, Kargarpour Z, Gruber R. Blocking of Caspases Exerts Anti-Inflammatory Effects on Periodontal Cells. Life. 2022; 12(7):1045. https://doi.org/10.3390/life12071045
Chicago/Turabian StylePanahipour, Layla, Lara Cristina Cunha Cervantes, Azarakhsh Oladzad Abbasabadi, Mariane Beatriz Sordi, Zahra Kargarpour, and Reinhard Gruber. 2022. "Blocking of Caspases Exerts Anti-Inflammatory Effects on Periodontal Cells" Life 12, no. 7: 1045. https://doi.org/10.3390/life12071045