
Manipulation of Insect Vectors’ Host Selection Behavior by Barley Yellow Dwarf Virus Is Dependent on the Host Plant Species and Viral Co-Infection


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Insect Colonies and Virus Inoculation
2.3. Gene Expression Analysis
2.4. Aphid Preference Tests
2.5. Data Analysis
3. Results
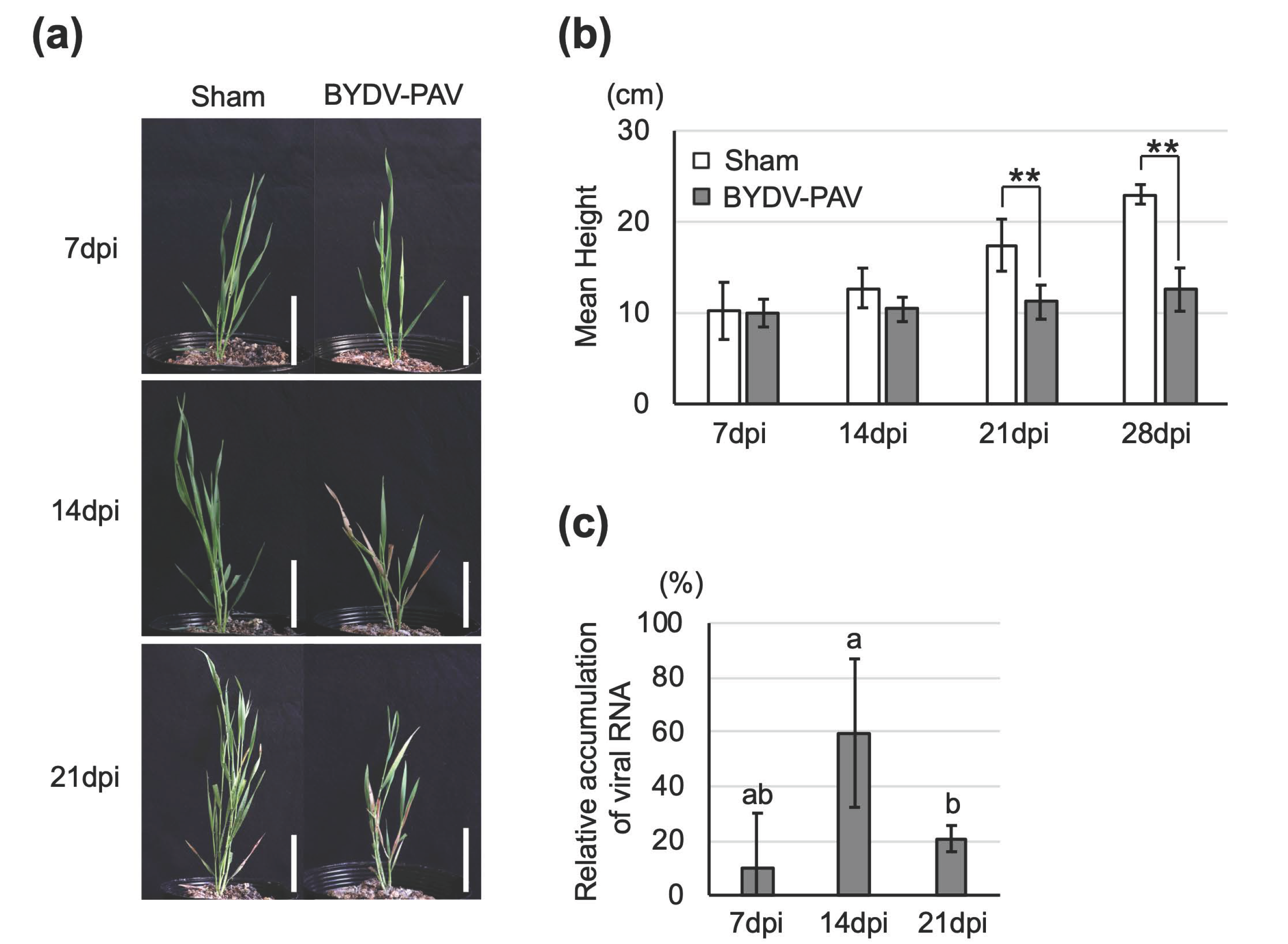
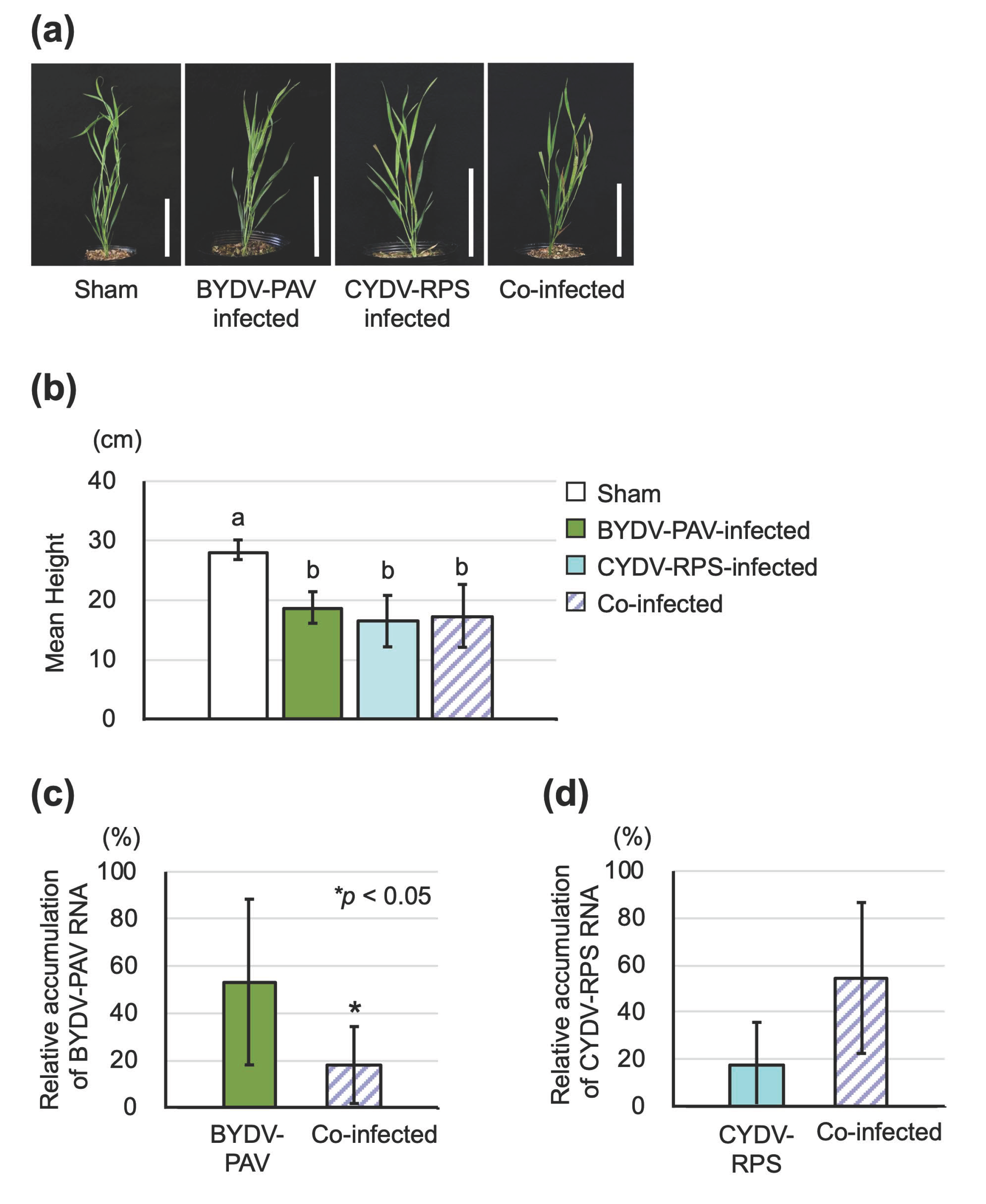
3.1. BYDV-PAV and CYDV-RPS Infect Brachypodium Distachyon via an Aphid Vector
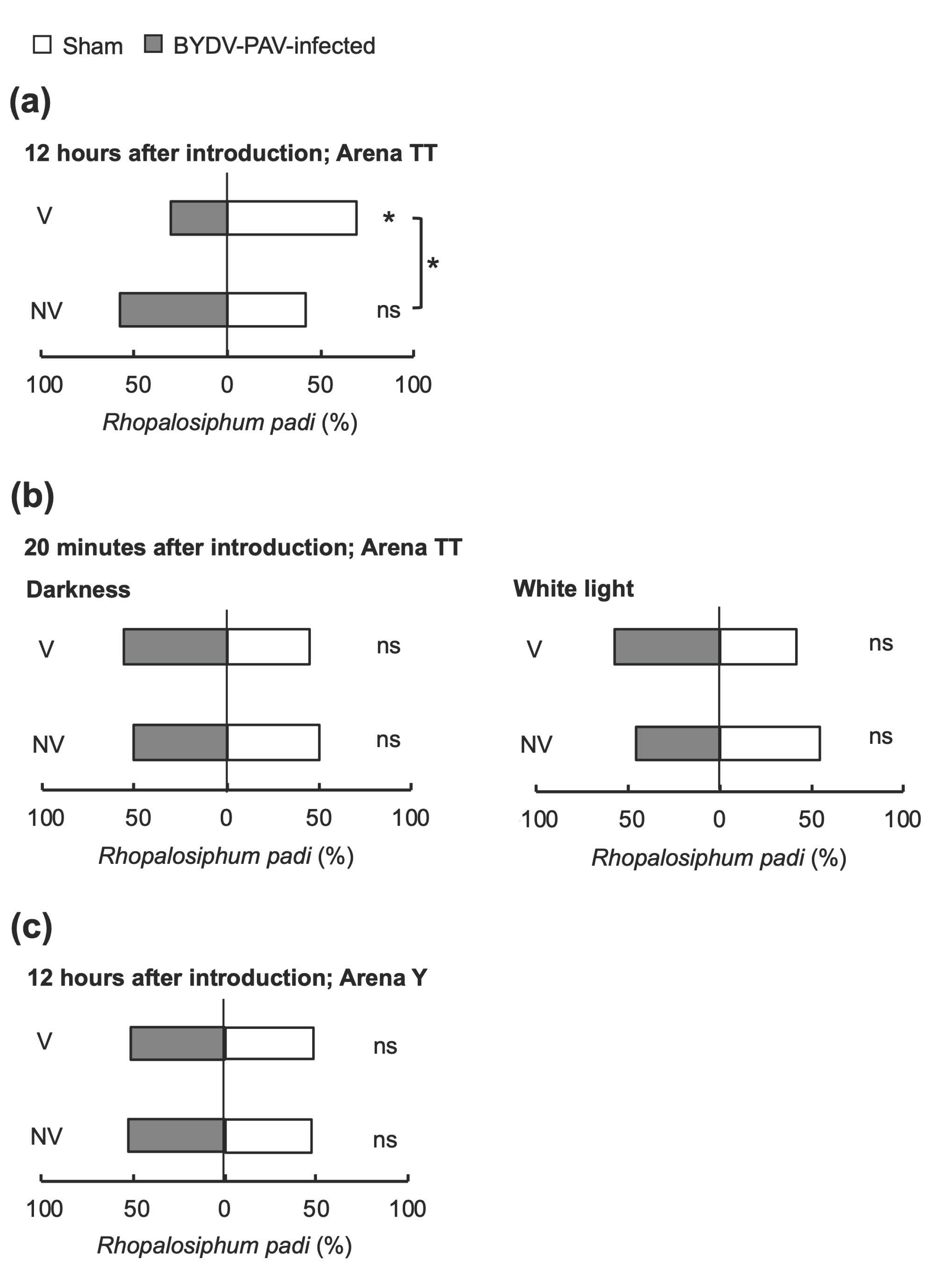
3.2. BYDV-PAV Affects the Host Preference of the Aphid Vector R. padi on Wheat
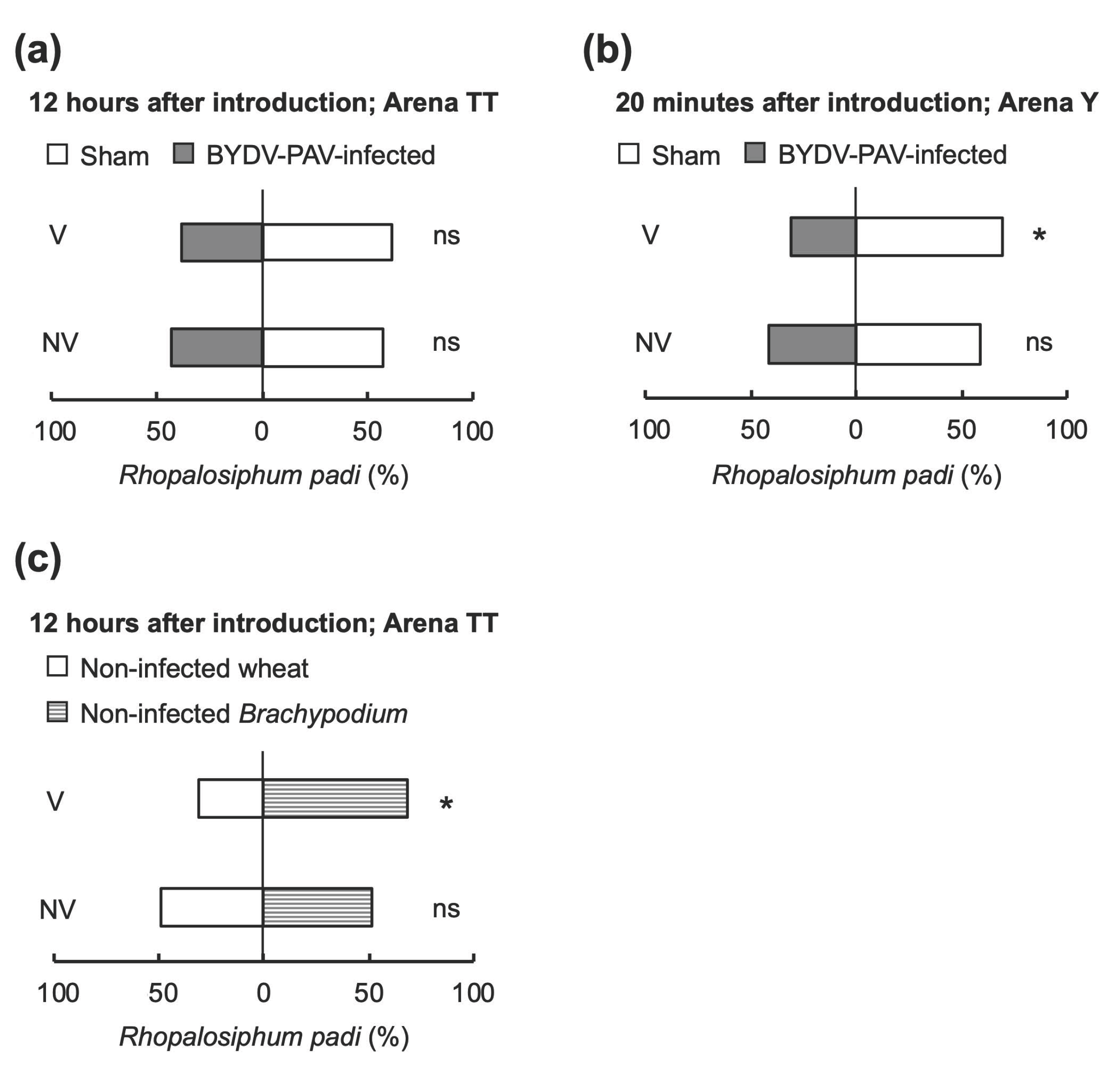
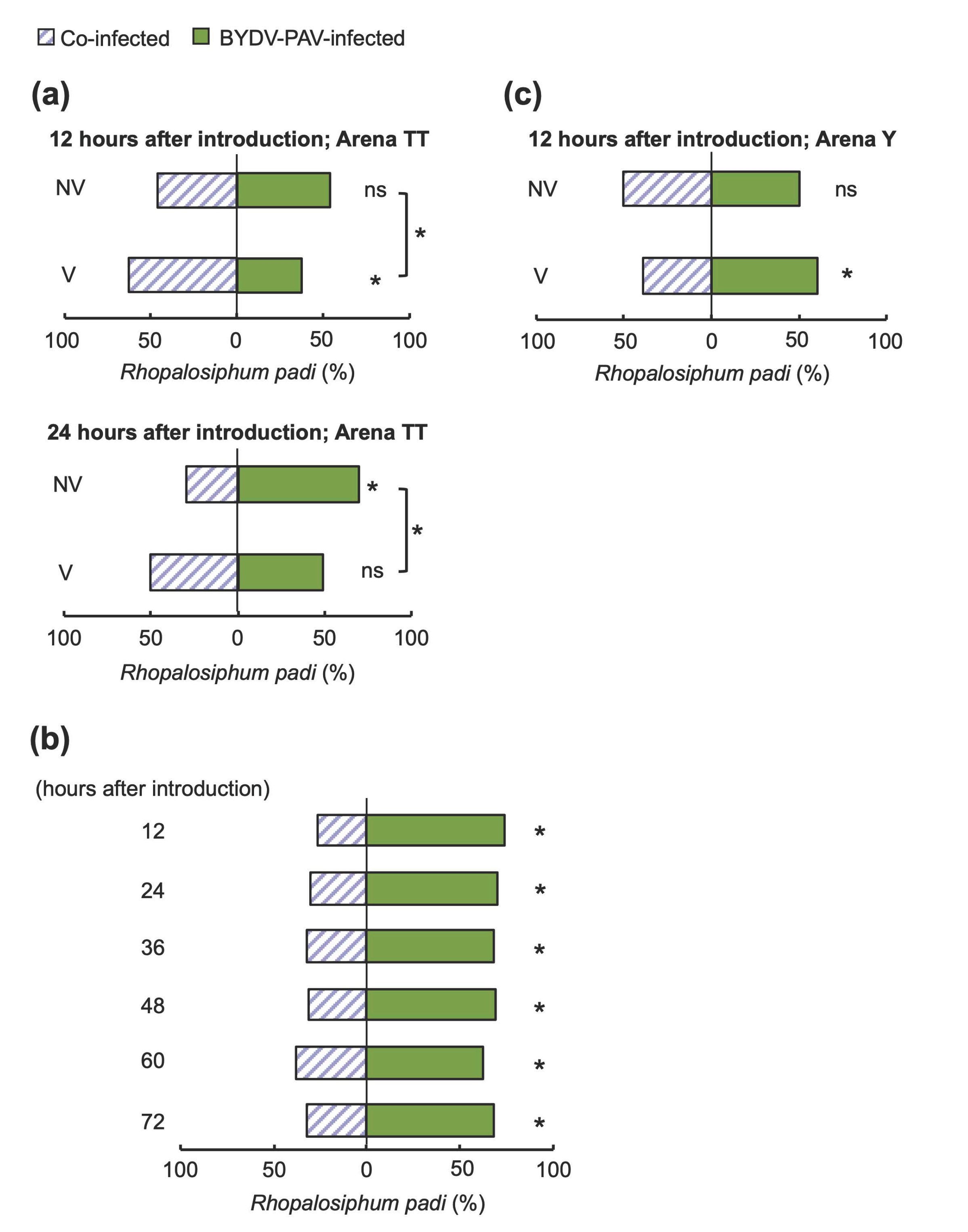
3.3. Alteration of Aphids’ Host Preference by BYDV-PAV Depends on the Host Poaceae Species
3.4. Insect Vectors Harboring Only BYDV-PAV Facilitated the Co-Infection of Multiple Viruses on Wheat
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hogenhout, S.A.; Ammar, E.-D.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauck, K.; Bosque-Pérez, N.A.; Eigenbrode, S.D.; De Moraes, C.M.; Mescher, M.C. Transmission mechanisms shape pathogen effects on host–vector interactions: Evidence from plant viruses. Funct. Ecol. 2012, 26, 1162–1175. [Google Scholar] [CrossRef]
- Eigenbrode, S.D.; Bosque-Pérez, N.A.; Davis, T.S. Insect-borne plant pathogens and their vectors: Ecology, evolution, and complex interactions. Annu. Rev. Entomol. 2018, 63, 169–191. [Google Scholar] [CrossRef] [PubMed]
- Mauck, K.E.; Chesnais, Q.; Shapiro, L.R. Evolutionary determinants of host and vector manipulation by plant viruses. Adv. Virus Res. 2018, 101, 189–250. [Google Scholar]
- Mauck, K.E.; Chesnais, Q. A synthesis of virus-vector associations reveals important deficiencies in studies on host and vector manipulation by plant viruses. Virus Res. 2020, 285, 197957. [Google Scholar] [CrossRef]
- Ingwell, L.L.; Eigenbrode, S.D.; Bosque-Pérez, N.A. Plant viruses alter insect behavior to enhance their spread. Sci. Rep. 2012, 2, 578. [Google Scholar] [CrossRef] [Green Version]
- Rajabaskar, D.; Bosque-Pérez, N.A.; Eigenbrode, S.D. Preference by a virus vector for infected plants is reversed after virus acquisition. Virus Res. 2014, 186, 32–37. [Google Scholar] [CrossRef]
- Mauck, K.E. Variation in virus effects on host plant phenotypes and insect vector behavior: What can it teach us about virus evolution? Curr. Opin. Virol. 2016, 21, 114–123. [Google Scholar] [CrossRef]
- Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Effects of pathogens on sensory-mediated interactions between plants and insect vectors. Curr. Opin. Plant Biol. 2016, 32, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Ziegler-Graff, V. Molecular insights into host and vector manipulation by plant viruses. Viruses 2020, 12, 263. [Google Scholar] [CrossRef] [Green Version]
- Porras, M.F.; Navas, C.A.; Marden, J.H.; Mescher, M.C.; De Moraes, C.M.; Pincebourde, S.; Sandoval-Mojica, A.; Raygoza-Garay, J.A.; Holguin, G.A.; Rajotte, E.G. Enhanced heat tolerance of viral-infected aphids leads to niche expansion and reduced interspecific competition. Nat. Commun. 2020, 11, 1184. [Google Scholar] [PubMed]
- Burnett, P.A. World Perspectives on Barley Yellow Dwarf: Proceedings of the International Workshop July 6–11, 1987, Undine Italy; CIMMYT: Veracruz, Mexico, 1990. [Google Scholar]
- İLbaĞI, H.; Aydin, Ö. Firs t Report of Yellow Dwarf Viruses (YDVs) in the Rice Fields in the Trakya Region of Turkey. Ekin J. Crop Breed. Genet. 2020, 6, 96–101. [Google Scholar]
- Ilbaği, H.; Çitir, A.; Kara, A.; Uysal, M.; Olden, F.A. First report of Barley Yellow Dwarf Viruses (BYDVs) on dicotyledonous weed hosts in Turkey. Cereal Res. Commun. 2019, 47, 292–303. [Google Scholar]
- Riedell, W.E.; Kieckhefer, R.W.; Langham, M.A.C.; Hesler, L.S. Root and shoot responses to bird cherry-oat aphids and barley yellow dwarf virus in spring wheat. Crop Sci. 2003, 43, 1380–1386. [Google Scholar]
- Scholthof, K.B.G.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahlquist, P. Top 10 plant viruses in molecular plant pathology. Mol. Plant Pathol. 2011, 12, 938–954. [Google Scholar]
- Rybicki, E.P. A Top Ten list for economically important plant viruses. Arch. Virol. 2015, 160, 17–20. [Google Scholar] [CrossRef]
- Toriyama, S.; Yora, K.; Asuyama, H. Occurrence of Barley yellow dwarf virus (BYDV) in Japan. In Proceedings of the Autumn Meeting of the Kanto Division, Annual Meeting of the Phytopathological Society of Japan, Tokyo, Japan; 1968; p. 374. [Google Scholar]
- Kojima, M.; Matsubara, A.; Yanase, S.; Toriyama, S. The occurrence of barley yellow dwarf disease in Japan. Jpn. J. Phytopathol. 1983, 49, 338–346. [Google Scholar] [CrossRef]
- Usugi, T.; Nakano, M.; Shinkai, A. Occurrence of barley yellow dwarf virus and wheat yellow leaf virus in Kyushu. Proc. Assoc. Plant Prot. Kyushu 1987, 33, 21–23. [Google Scholar]
- Fan, Y.-J.; Namba, S.; Yamashita, S.; Doi, Y. Some properties of barley yellow dwarf virus (BYDV) from maize and some cereal plants. Jpn. J. Phytopathol. 1994, 60, 725–728. [Google Scholar]
- Trebicki, P. Climate change and plant virus epidemiology. Virus Res. 2020, 286, 198059. [Google Scholar] [CrossRef]
- McElhany, P.; Real, L.A.; Power, A.G. Vector preference and disease dynamics: A study of barley yellow dwarf virus. Ecology 1995, 76, 444–457. [Google Scholar] [CrossRef]
- Jiménez-Martínez, E.S.; Bosque-Pérez, N.A.; Berger, P.H.; Zemetra, R.S.; Ding, H.; Eigenbrode, S.D. Volatile cues influence the response of Rhopalosiphum padi (Homoptera: Aphididae) to Barley yellow dwarf virus–infected transgenic and untransformed wheat. Environ. Entomol. 2004, 33, 1207–1216. [Google Scholar] [CrossRef] [Green Version]
- Medina-Ortega, K.J.; Bosque-Pérez, N.A.; Ngumbi, E.; Jiménez-Martínez, E.S.; Eigenbrode, S.D. Rhopalosiphum padi (Hemiptera: Aphididae) responses to volatile cues from barley yellow dwarf virus–infected wheat. Environ. Entomol. 2009, 38, 836–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porras, M.; De Moraes, C.M.; Mescher, M.C.; Rajotte, E.G.; Carlo, T.A. A plant virus (BYDV) promotes trophic facilitation in aphids on wheat. Sci. Rep. 2018, 8, 11709. [Google Scholar] [PubMed] [Green Version]
- Seabloom, E.W.; Hosseini, P.R.; Power, A.G.; Borer, E.T. Diversity and composition of viral communities: Coinfection of barley and cereal yellow dwarf viruses in California grasslands. Am. Nat. 2009, 173, E79–E98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Khine, M.O.; Zhang, P.; Fu, Y.; Wang, X. Incidence and distribution of insect-transmitted cereal viruses in wheat in China from 2007 to 2019. Plant Dis. 2020, 104, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Laney, A.G.; Acosta-Leal, R.; Rotenberg, D. Optimized yellow dwarf virus multiplex PCR assay reveals a common occurrence of Barley yellow dwarf virus-PAS in Kansas winter wheat. Plant Health Prog. 2018, 19, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Mordecai, E.A.; Hindenlang, M.; Mitchell, C.E. Differential impacts of virus diversity on biomass production of a native and an exotic grass host. PLoS ONE 2015, 10, e0134355. [Google Scholar]
- Malmstrom, C.M.; Bigelow, P.; Trębicki, P.; Busch, A.K.; Friel, C.; Cole, E.; Abdel-Azim, H.; Phillippo, C.; Alexander, H.M. Crop-associated virus reduces the rooting depth of non-crop perennial native grass more than non-crop-associated virus with known viral suppressor of RNA silencing (VSR). Virus Res. 2017, 241, 172–184. [Google Scholar]
- Wang, H.; Xu, D.; Pu, L.; Zhou, G. Southern rice black-streaked dwarf virus alters insect vectors’ host orientation preferences to enhance spread and increase rice ragged stunt virus co-infection. Phytopathology 2014, 104, 196–201. [Google Scholar]
- Lightle, D.; Lee, J. Raspberry viruses affect the behavior and performance of A mphorophora agathonica in single and mixed infections. Entomol. Exp. Appl. 2014, 151, 57–64. [Google Scholar] [CrossRef]
- Peñaflor, M.F.G.V.; Mauck, K.E.; Alves, K.J.; De Moraes, C.M.; Mescher, M.C. Effects of single and mixed infections of Bean pod mottle virus and Soybean mosaic virus on host-plant chemistry and host–vector interactions. Funct. Ecol. 2016, 30, 1648–1659. [Google Scholar] [CrossRef]
- Senshu, H.; Yamaji, Y.; Minato, N.; Shiraishi, T.; Maejima, K.; Hashimoto, M.; Miura, C.; Neriya, Y.; Namba, S. A dual strategy for the suppression of host antiviral silencing: Two distinct suppressors for viral replication and viral movement encoded by potato virus M. J. Virol. 2011, 85, 10269–10278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, J.P.; Behpouri, A.; Bird, A.; Ng, C.K.Y. Evaluation of the use of the Polyubiquitin Genes, Ubi4 and Ubi10 as reference genes for expression studies in Brachypodium distachyon. PLoS ONE 2012, 7, e49372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Nadege, S.W.; Huang, C.; Zhang, P.; Song, S.; Sun, L.; Wu, Y. Brachypodium distachyon is a suitable host plant for study of Barley yellow dwarf virus. Virus Genes 2016, 52, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Draper, J.; Mur, L.A.J.; Jenkins, G.; Ghosh-Biswas, G.C.; Bablak, P.; Hasterok, R.; Routledge, A.P.M. Brachypodium distachyon. A new model system for functional genomics in grasses. Plant Physiol. 2001, 127, 1539–1555. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, K.-B.G.; Irigoyen, S.; Catalan, P.; Mandadi, K.K. Brachypodium: A monocot grass model genus for plant biology. Plant Cell 2018, 30, 1673–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Zhang, X.; Gong, Y.; Wang, D.; Xu, D.; Wang, N.; Sun, Y.; Gao, L.; Liu, S.-S.; Deng, X.W. Red-light is an environmental effector for mutualism between begomovirus and its vector whitefly. PLoS Pathog. 2021, 17, e1008770. [Google Scholar] [CrossRef]
- Chesnais, Q.; Couty, A.; Uzest, M.; Brault, V.; Ameline, A. Plant infection by two different viruses induce contrasting changes of vectors fitness and behavior. Insect Sci. 2019, 26, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Gao, Y.; Yan, S.; Tang, X.; Zhou, X.; Zhang, D.; Liu, Y. Aphid performance changes with plant defense mediated by Cucumber mosaic virus titer. Virol. J. 2016, 13, 70. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minato, N.; Hatori, S.; Okawa, A.; Nakagawa, K.; Hironaka, M. Manipulation of Insect Vectors’ Host Selection Behavior by Barley Yellow Dwarf Virus Is Dependent on the Host Plant Species and Viral Co-Infection. Life 2022, 12, 644. https://doi.org/10.3390/life12050644
Minato N, Hatori S, Okawa A, Nakagawa K, Hironaka M. Manipulation of Insect Vectors’ Host Selection Behavior by Barley Yellow Dwarf Virus Is Dependent on the Host Plant Species and Viral Co-Infection. Life. 2022; 12(5):644. https://doi.org/10.3390/life12050644
Chicago/Turabian StyleMinato, Nami, Shuichi Hatori, Azusa Okawa, Kai Nakagawa, and Mantaro Hironaka. 2022. "Manipulation of Insect Vectors’ Host Selection Behavior by Barley Yellow Dwarf Virus Is Dependent on the Host Plant Species and Viral Co-Infection" Life 12, no. 5: 644. https://doi.org/10.3390/life12050644