
The Effect of Enterococcus faecium AL41 on the Acute Phase Proteins and Selected Mucosal Immune Molecules in Broiler Chickens

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Scheme
Animals
2.2. Preparing of Probiotic Strain
2.3. Laboratory Analyses
2.3.1. Homogenization of Jejunal Samples and Isolation of Total RNA of IgA, MUC-2 and IGF-2 (Growth Factor) Gene
2.3.2. Relative Expression of Genes in Quantitative Real-Time PCR (qRT-PCR)
2.4. Mucus Production
2.5. Microbiology
2.6. Statistical Analyses
3. Results
3.1. Laboratory Analyses
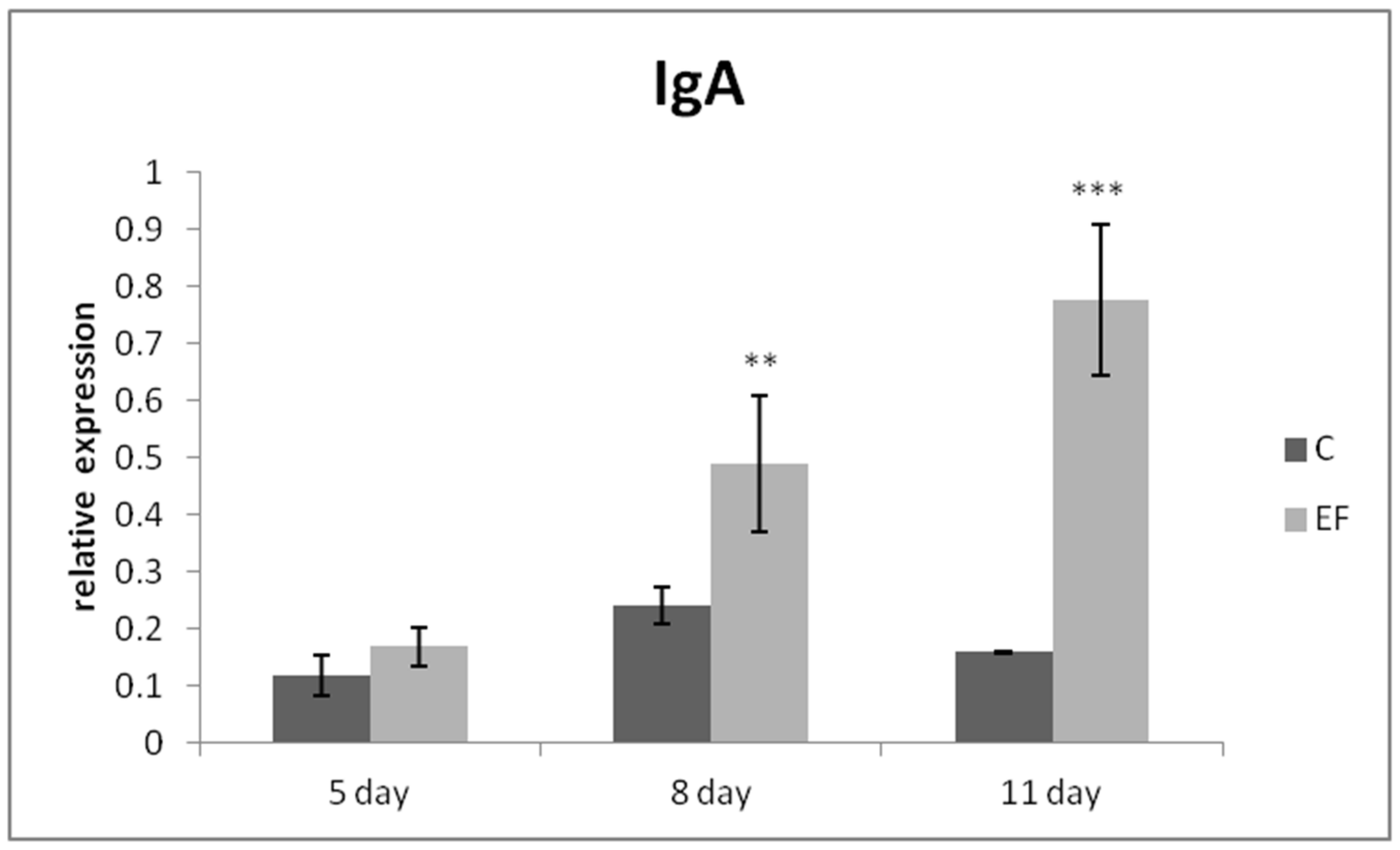
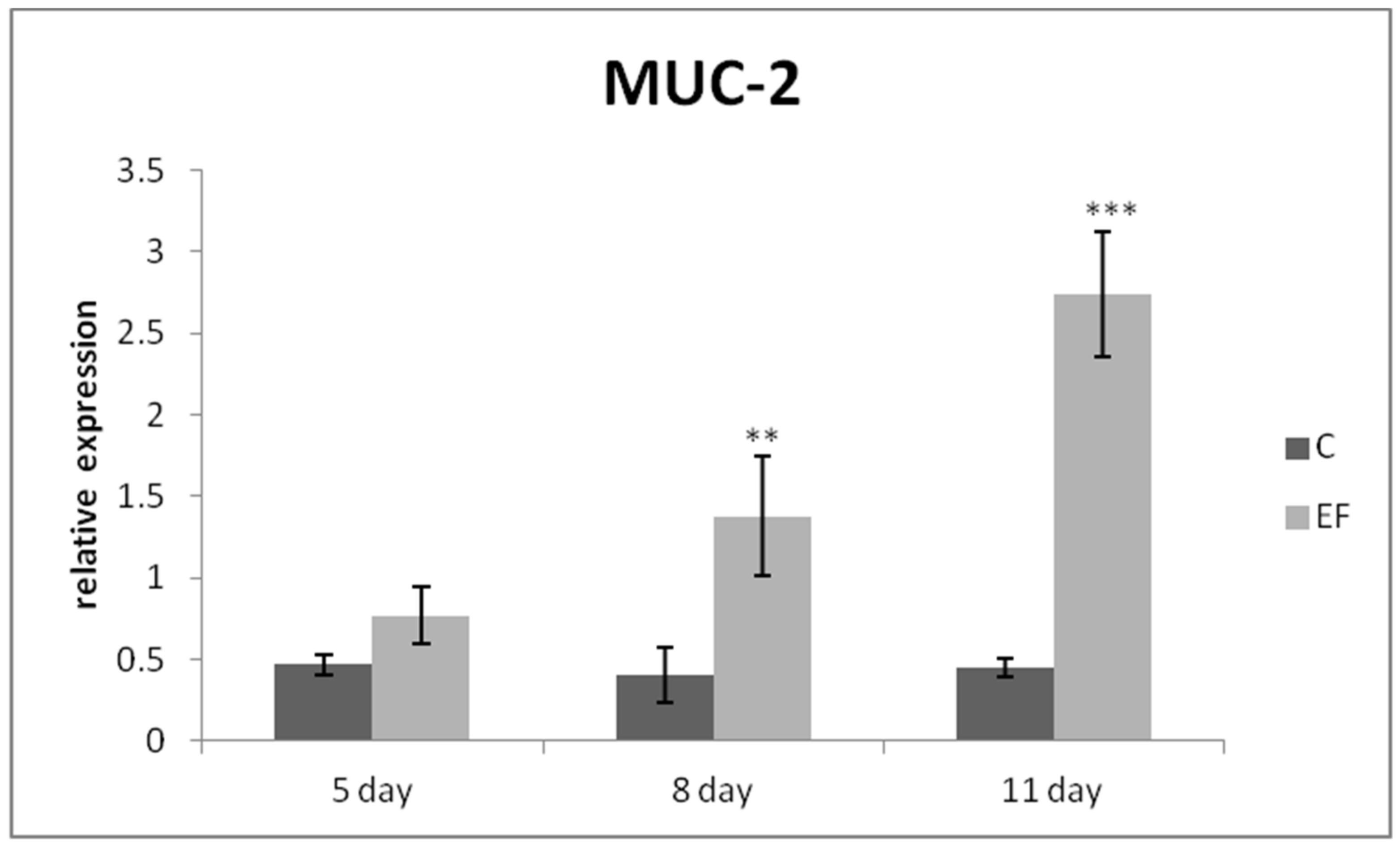
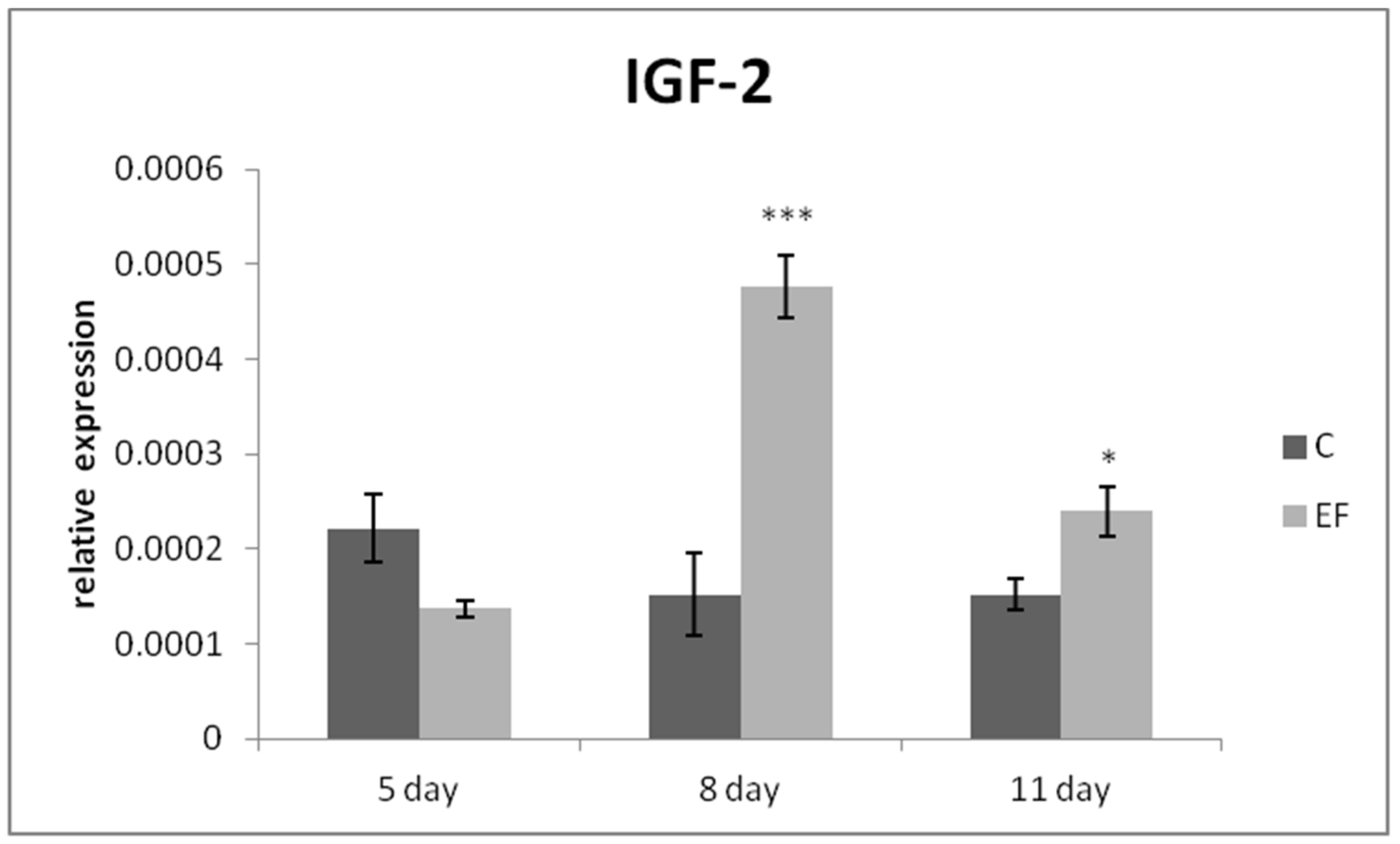
3.2. Relative Expression of Genes in qRT-PCR
3.3. Mucus Production
3.4. Microbial Screening
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Nochi, T.; Jansen, C.A.; Toyomizu, M.; van Eden, W. The well-developed mucosal immune systems of birds and mammals allow for similar approaches of mucosal vaccination in both types of animals. Front. Nutr. 2018, 5, 60. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Eicher, S.D.; Applegate, T.J. Development of intestinal mucin 2, IgA, and polymeric Ig receptor expressions in broiler chickens and Pekin ducks. Poult. Sci. 2015, 94, 172–180. [Google Scholar] [CrossRef]
- Everett, M.L.; Palestrant, D.; Miller, S.E.; Bollinger, R.R.; Parker, W. Immune exclusion and immune inclusion: A new model of host-bacterial interactions in the gut. Clin. Appl. Immunol. Rev. 2004, 4, 321–332. [Google Scholar] [CrossRef]
- Sun, R.C.; Choi, P.M.; Guo, J.; Erwin, C.R.; Warner, B.W. Insulin-like growth factor 2 and its enterocyte receptor are not required for adaptation in response to massive small bowel resection. J. Pediatr. Surg. 2014, 49, 966–970. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, S.; Suchodolski, J. Understanding the canine intestinal microbiota and its modification by pro-, pre- and synbiotics—what is the evidence? Vet. Med. Sci. 2016, 2, 71–94. [Google Scholar] [CrossRef]
- Duangnumsawang, Y.; Zentek, J.; Goodarzi Boroojeni, F. Development and Functional Properties of Intestinal Mucus Layer in Poultry. Front. Immunol. 2021, 12, 745849. [Google Scholar] [CrossRef]
- Jha, R.; Das, R.; Oak, S.; Mishra, P. Probiotics (Direct-Fed Microbials) in Poultry Nutrition and Their Effects on Nutrient Utilization, Growth and Laying Performance, and Gut Health: A Systematic Review. Animals 2020, 10, 1863. [Google Scholar] [CrossRef]
- Forte, C.; Acuti, G.; Manuali, E.; Casagrande Proietti, P.; Pavone, S.; Trabalza-Marinucci, M.; Moscati, L.; Onofri, A.; Lorenzetti, C.; Franciosini, M.P. Effects of two different probiotics on microflora, morphology, and morphometry of gut in organic laying hens. Poult. Sci. 2016, 95, 2528–2535. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Long, S.; Mahfuz, S.; Wu, D.; Wang, X.; Wei, X.; Piao, X. Effects of Probiotics as Antibiotics Substitutes on Growth Performance, Serum Biochemical Parameters, Intestinal Morphology, and Barrier Function of Broilers. Animals 2019, 9, 985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pogány Simonová, M.; Chrastinová, Ľ.; Lauková, A. Effect of Enterococcus faecium AL41 (CCM8558) and its enterocin m on the physicochemical properties and mineral content of rabbit meat. Agriculture 2021, 11, 1045. [Google Scholar] [CrossRef]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The Genus Enterococcus: Between probiotic potential and safety concerns-an update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef]
- Hanifeh, M.; Spillmann, T.; Huhtinen, M.; Sclivagnotis, Y.S.; Grönthal, T.; Hynönen, U. Ex-Vivo adhesion of Enterococcus faecalis and Eenterococcus faecium to the intestinal mucosa of healthy beagles. Animals 2021, 11, 3283. [Google Scholar] [CrossRef]
- Lauková, A.; Kandričáková, A.; Ščerbová, J. Use of bacteriocin-producing, probiotic strain Enterococcus faecium AL41 to control intestinal microbiota in farm ostriches. Lett. Appl. Microbiol. 2015, 60, 531–535. [Google Scholar] [CrossRef]
- Mareková, M.; Lauková, A.; Skaugen, M.; Nes, I. Isolation and characterization of a new bacteriocin, termed enterocin M, produced by environmental isolate Enterococcus faecium AL41. J. Ind. Microbiol. Biotechnol. 2007, 34, 533–537. [Google Scholar] [CrossRef]
- Huang, L.; Luo, L.; Zhang, Y.; Wang, Z.; Xia, Z. Effects of the dietary probiotic, Enterococcus faecium NCIMB11181, on the intestinal barrier and system immune status in Escherichia coli O78-challenged broiler chickens. Probiotics Antimicrob. Proteins 2019, 11, 946–956. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, S.; Asif, M.; Bhutta, Z.A.; Kulyar, M.F.; Hussain, R.; Ramzan, A.; Shafeeq, S.; Shakir, M.Z.; Sarfaraz, M.T.; Li, K. A comprehensive review on acute phase proteins in chicken. Eur. Poult. Sci. 2021, 85, 344. [Google Scholar]
- Skeldon, N. Interpreting protein electrophoresis in practice. Practice 2018, 40, 183–193. [Google Scholar] [CrossRef]
- Karaffová, V.; Bobíková, K.; Husáková, E.; Levkut, M.; Herich, R.; Revajová, V.; Levkutová, M.; Levkut, M. Interaction of TGF-β4 and IL-17 with IgA secretion in the intestine of chickens fed with E. faecium AL41 and challenged with S. enteritidis. Res. Vet. Sci. 2015, 100, 75–79. [Google Scholar] [CrossRef]
- Letnická, A.; Karaffová, V.; Levkut, M.; Revajová, V.; Herich, R. Influence of oral application of Enterococcus faecium AL41 on TGF-β4 and IL-17 expression and immunocompetent cell distribution in chickens challenged with Campylobacter jejuni. Acta Vet. Hung. 2017, 65, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Cobb-Vantress Inc. Cobb500 Slow Feather Breeder Management Supplement; Cobb-Vantress: Siloam Springs, AR, USA, 2020. [Google Scholar]
- Karaffová, V.; Marcinková, E.; Bobíková, K.; Herich, R.; Revajová, V.; Stašová, D.; Kavuľová, A.; Levkutová, M.; Levkut, M., Jr.; Lauková, A.; et al. TLR4 and TLR21 expression, MIF, IFN-β, MD-2, CD14 activation, and sIgA production in chickens administered with EFAL41 strain challenged with Campylobacter jejuni. Folia Microbiol. 2017, 62, 89–97. [Google Scholar]
- Nagy, O.; Tóthová, C.; Nagyová, V.; Kováč, G. Comparison of serum protein electrophoretic pattern in cows and small ruminants. Acta Vet. Brno 2015, 84, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Karaffová, V.; Bobíková, K.; Levkut, M.; Revajová, V.; Ševčíková, Z.; Levkut, M. The influence of Farmatan® and Flimabend® on the mucosal immunity of broiler chicken. Poult. Sci. 2019, 98, 1161–1166. [Google Scholar] [CrossRef]
- Lammers, A.; Wieland, W.H.; Kruijt, L.; Jansma, A.; Straetemans, T.; Schots, A.; den Hartog, G.; Parmentier, H.K. Successive immunoglobulin and cytokine expression in the small intestine of juvenile chicken. Dev. Comp. Immunol. 2010, 34, 1254–1262. [Google Scholar] [CrossRef]
- Smirnov, A.; Tako, E.; Ferket, P.R.; Uni, Z. Mucin gene expression and mucin content in the chicken intestinal goblet cells are affected by in ovo feeding of carbohydrates. Poult. Sci. 2006, 85, 669–673. [Google Scholar] [CrossRef]
- Mudroňová, D.; Karaffová, V.; Košcová, J.; Bartkovský, M.; Marcincáková, D.; Popelka, P.; Klempová, T.; Certík, M.; Mačanga, J.; Marcincák, S. Effect of fungal gamma-linolenic acid and beta-carotene containing prefermented feed on immunity and gut of broiler chicken. Poult. Sci. 2018, 97, 4211–4218. [Google Scholar] [CrossRef]
- De Boever, S.; Vangestel, C.; De Backer, P.; Croubels, S.; Sys, S.U. Identification and validation of housekeeping genes as internal control for gene expression in an intravenous LPS inflammation model in chickens. Vet. Immunol. Immunopathol. 2008, 122, 312–317. [Google Scholar] [CrossRef]
- Faixová, Z.; Piešová, E.; Maková, Z.; Levkut, M., Jr.; Pistl, J.; Lauková, A.; Faix, Š.; Levkut, M. Effect of dietary probiotic and plant extract supplementation on mucin dynamics in the chicken intestine and on performance of chickens. Folia Vet. 2012, 56, 15–16. [Google Scholar]
- Alkhalf, A.; Alhaj, M.; Al-Homidan, I. Influence of probiotic supplementation on blood parameters and growth performance in broiler chickens. Saudi J. Biol. Sci. 2010, 17, 219–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.W.; Lee, S.H.; Lillehoj, H.S.; Li, G.X.; Jang, S.I.; Babu, U.S.; Park, M.S.; Kim, D.K.; Lillehoj, E.P.; Neumann, A.P.; et al. Effects of direct-fed microbials on growth performance, gut morphometry, and immune characteristics in broiler chickens. Poult. Sci. 2010, 89, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Chamanza, R.; Toussaint, J.M.; Van Edeeren, A.M.; Van Veen, L.; Hulskamp-Koch, C.; Fabri, T.H.F. Serum amyloid A and transferrin in chicken. A preliminary investigation of using acute phase variables to assess diseases in chickens. Vet. Q. 1999, 21, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Kefal, S.; Toker, N.Y. Effects of probiotics on some acute phase proteins in broilers exposed to Salmonella typhimurium lipopolysaccharides. Arch. Geflügelk. 2006, 70, 270–277. [Google Scholar]
- Szabó, A.; Mézes, M.; Horn, P.; Sütő, Z.; Bázár, G.Y.; Romvári, R. Developmental dynamics of some blood biochemical parameters in the growing turkey (Meleagris gallopavo). Acta Vet. Hung. 2005, 53, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Zheng, A.; Luo, J.; Meng, K.; Li, J.; Bryden, W.L.; Chang, W.; Zhang, S.; Wang, L.X.N.; Liu, G.; Yao, B. Probiotic (Enterococcus faecium) induced responses of the hepatic proteome improves metabolic efficiency of broiler chickens (Gallus gallus). BMC Genom. 2016, 17, 89. [Google Scholar] [CrossRef] [Green Version]
- Cetin, N.; Güclü, B.K.; Cetin, E. The effects of probiotic and mannanoligosaccharide on some haematological and immunological parameters in turkeys. J. Vet. Med. 2005, 52, 263–267. [Google Scholar] [CrossRef]
- Capitelli, R.; Crosta, L. Overview of psittacine blood analysis and comparative retrospective study of clinical diagnosis, hematology and blood chemistry in selected psittacine species. Vet. Clin. Exot. Anim. 2013, 16, 71–120. [Google Scholar] [CrossRef]
- Stef, L.; Julean, C.; Cean, A.; Mot, D.; Stef, D.S.; Simiz, E.; Simiz, F.; Pet, I.; Marcu, A.; Corcionivoschi, N. Evaluation of the administration effects of probiotics against Campylobacter jejuni on the immune system of broiler chickens. J. Anim. Sci. Biotechnol. 2016, 49, 16–21. [Google Scholar]
- Dev, K.; Mir, N.A.; Biswas, A.; Kannoujja, J.; Begum, J.; Kant, R.; Mandal, A. Dietary symbiotic supplementation improves the growth performance, body antioxidant pool, serum biochemistry, meat quality, and lipid oxidative stability in broiler chickens. Anim. Nutr. 2020, 6, 325–332. [Google Scholar] [CrossRef]
- Ingenbleek, Y.; Young, V. Transthyretin (prealbumin) in health and disease: Nutritional implications. Annu. Rev. Nutr. 1994, 14, 495–533. [Google Scholar] [CrossRef]
- Bogusławska-Tryk, M.; Ziółkowska, E.; Sławinska, A.; Siwek, M.; Bogucka, J. Modulation of intestinal histology by probiotics, prebiotics and synbiotics delivered in ovo in distinct chicken genotypes. Animals 2021, 11, 3293. [Google Scholar] [CrossRef]
- El-Hack, M.E.A.; El-Saadony, M.T.; Shafi, M.E.; Qattan, S.Y.A.; Batiha, G.E.; Khafaga, A.F.; Abdel-Moneim, A.E.; Alagawany, M. Probiotics in poultry feed: A comprehensive review. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1835–1850. [Google Scholar] [CrossRef]
- Sicard, J.F.; Le Bihan, G.; Vogeleer, P.; Jacques, M.; Harel, J. Interactions of intestinal bacteria with components of the intestinal mucus. Front. Cell Infect. Microbiol. 2017, 7, 387. [Google Scholar] [CrossRef] [PubMed]
- Ohland, C.L.; MacNaughton, W.K. Probiotic bacteria and intestinal epithelial barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G807–G819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levkut, M., Jr.; Karaffová, V.; Levkutová, M.; Seman, V.; Revajová, V.; Ševčíková, Z.; Herich, R.; Levkut, M. Influence of Lacto-Immuno-Vital on growth performance and gene expression of IgA, MUC-2, and growth factor IGF-2 in the jejunum of broiler chickens. Poult. Sci. 2020, 99, 6569–6575. [Google Scholar] [CrossRef] [PubMed]
- Aliakbarpour, H.; Chamani, M.; Rahimi, G.; Sadeghi, A.; Qujeq, D. The Bacillus subtilis and lactic acid bacteria probiotics influences intestinal mucin gene expression, histomorphology and growth performance in broilers. Anim. Biosci. 2012, 25, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Freier, S.; Eran, M.; Reinus, C.; Ariel, I.; Faber, J.; Wilschanski, M.; Braverman, D. Relative expression and localization of the insulin-like growth factor system components in the fetal, child and adult intestine. J. Pediatr. Gastroenterol. Nutr. 2005, 40, 202–209. [Google Scholar] [CrossRef]
- Wu, Y.; Zhen, W.; Geng, Y.; Wang, Z.; Guo, Y. Pretreatment with probiotic Enterococcus faecium NCIMB 11181 ameliorates necrotic enteritis-induced intestinal barrier injury in broiler chickens. Sci. Rep. 2019, 9, 10256. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ingredients g/kg | BR1 |
|---|---|
| Wheat | 290 |
| Maize | 300 |
| Soybean meal | 320 |
| Rapeseed oil | 40 |
| Fish meal | 20 |
| Limestone | 12 |
| Dicalcium phosphate | 10 |
| Sodium chloride | 2 |
| DL-methionine | 1 |
| Vitamin-mineral mix * | 5 |
| Composition by analysis (g/kg) | |
| dry matter | 899.9 |
| crude protein | 232.7 |
| fat | 64.5 |
| dietary fibre | 22.7 |
| ash | 53 |
| Ca (calcium) | 90.4 |
| P (phosphorus) total | 69.6 |
| Primer | Sequence 5′–3′ | Annealing/Temperature Time | References |
|---|---|---|---|
| IgA Fw | GTCACCGTCACCTGGACTACA | 59 °C for 30 s | [24] |
| IgA Rev | ACCGATGGTCTCCTTCACATC | ||
| MUC-2 Fw | GCTGATTGTCACTCACGCCTT | 54 °C for 1 min | [25] |
| MUC-2 Rev | ATCTGCCTGAATCACAGGTGC | ||
| IGF-2 Fw | CTCTGCTGGAAACCTACTGT | 55 °C/30 s | [26] |
| IGF-2 Rev | GAGTACTTGGCATGAGATGG | ||
| GAPDH Fw | CCTGCATCTGCCCATTT | 59 °C/30 s | [27] |
| GAPDH Rev | GGCACGCCATCACTATC |
| Parameter | Groups of Animals | ||||
|---|---|---|---|---|---|
| C (Control) | EF | ||||
| Day 0 | Day 11 | Day 0 | Day 11 | ||
| Hp (mg/mL) | 0.052 ± 0.071 | 0.025 ± 0.033 | 0.014 ± 0.013 | 0.022 ± 0.029 | |
| SAA (ng/mL) | 34.30 ± 11.38 | 48.62 ± 17.16 | 35.41 ± 12.30 | 44.22 ± 6.81 | |
| TP (g/L) | 26.7 ± 2.06 | 29.7 ± 1.46 b | 26.8 ± 2.19 | 29.1 ± 1.78 a | |
| prealb | % | 1.46 ± 0.30 | 1.36 ± 0.30 | 1.53 ± 0.29 | 1.77 ± 0.49 |
| g/L | 0.39 ±0.09 | 0.40 ± 0.08 | 0.41 ± 0.11 | 0.51 ± 0.13 | |
| alb | % | 43.9 ± 1.63 | 43.2 ± 2.49 | 43.7 ± 1.75 | 42.1 ± 2.72 |
| g/L | 11.7 ± 0.62 | 12.8 ± 0.76 b | 11.7 ± 1.23 | 12.3 ± 1.10 | |
| α1- | % | 4.6 ± 0.65 | 4.7 ± 0.67 | 4.2 ± 0.62 | 4.0 ± 0.52 |
| g/L | 1.2 ± 0.28 | 1.4 ± 0.17 | 1.1 ± 0.22 | 1.2 ± 0.14 | |
| α2- | % | 29.6 ± 0.90 | 27.9 ± 1.44 a | 29.5 ± 1.64 | 27.6 ± 1.11 a |
| g/L | 7.8 ± 0.73 | 8.3 ± 0.41 | 7.9 ± 0.53 | 8.0 ± 0.70 | |
| β- | % | 6.6 ± 0.50 | 6.6 ± 0.67 | 6.4 ± 1.14 | 7.3 ± 0.77 |
| g/L | 1.8 ± 0.11 | 2.0 ± 0.22 | 1.7 ± 0.30 | 2.1 ± 0.12 b | |
| γ- | % | 13.8 ± 0.98 | 16.2 ± 3.05 | 14.7 ± 1.26 | 17.2 ± 2.00 b |
| g/L | 3.7 ± 0.55 | 4.8 ± 1.12 a | 3.9 ± 0.41 | 5.0 ± 0.69 b | |
| A/G | 1.00 ± 0.08 | 0.94 ± 0.15 | 1.00 ± 0.10 | 0.94 ± 0.12 | |
| Mucus Production (g ± SD) | 5th Day of Experiment | 8th Day of Experiment | 11th Day of Experiment |
|---|---|---|---|
| Duodenum | |||
| EF | 3.66 ± 1.22 | 3.21 ± 1.21 d | 4.44 ± 0.27 a,d |
| C | 2.37 ± 0.69 | 1.41 ± 0.26 | 1.72 ± 0.02 a |
| Jejunum | |||
| EF | 3.27 ± 1.65 | 2.99 ± 0.44 e | 3.91 ± 0.25 b,e |
| C | 2.82 ± 3.38 | 1.82 ± 0.17 | 1.66 ± 0.07 b |
| Ileum | |||
| EF | 3.70 ± 1.71 | 3.77 ± 1.37 | 4.11 ± 0.44 c |
| C | 1.12 ± 4.51 | 1.78 ± 0.18 | 1.33 ± 0.60 c |
| Faeces | Control | EF |
|---|---|---|
| 0 day | ||
| EFAL41 | nt | nt |
| Enterococci | 6.46 (0.48) | 6.43 (0.47) |
| Coliform | 7.1 (0.0) | 6.92 (0.84) |
| 8 day | ||
| EFAL41 | nt | 2.60 (1.0) |
| Enterococci | 6.54 (0.81) | 5.29 (0.23) * |
| Coliform | 6.94 (0.74) | 7.1 (0.0) |
| 11 day | ||
| EFAL41 | nt | 2.17 (0.31) |
| Enterococci | 6.48 (0.81) | 5.09 (0.70) * |
| Coliform | 6.91 (0.84) | 6.98 (0.83) |
| Cecum | Control | EF |
|---|---|---|
| 8 day | ||
| EFAL41 | nt | 2.76 (0.43) |
| Enterococci | 6.40 (0.81) | 5.63 (0.75) * |
| Coliform | 6.79 (0.83) | 6.81 (0.83) |
| 11 day | ||
| EFAL41 | 2.16 (0.33) | |
| Enterococci | 4.61 (2.01) | 4.46 (1.37) |
| Coliform | 5.69 (0.75) | 5.87 (1.42) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karaffová, V.; Tóthová, C.; Szabóová, R.; Revajová, V.; Lauková, A.; Ševčíková, Z.; Herich, R.; Žitňan, R.; Levkut, M.; Levkut, M.; et al. The Effect of Enterococcus faecium AL41 on the Acute Phase Proteins and Selected Mucosal Immune Molecules in Broiler Chickens. Life 2022, 12, 598. https://doi.org/10.3390/life12040598
Karaffová V, Tóthová C, Szabóová R, Revajová V, Lauková A, Ševčíková Z, Herich R, Žitňan R, Levkut M, Levkut M, et al. The Effect of Enterococcus faecium AL41 on the Acute Phase Proteins and Selected Mucosal Immune Molecules in Broiler Chickens. Life. 2022; 12(4):598. https://doi.org/10.3390/life12040598
Chicago/Turabian StyleKaraffová, Viera, Csilla Tóthová, Renáta Szabóová, Viera Revajová, Andrea Lauková, Zuzana Ševčíková, Róbert Herich, Rudolf Žitňan, Martin Levkut, Mikuláš Levkut, and et al. 2022. "The Effect of Enterococcus faecium AL41 on the Acute Phase Proteins and Selected Mucosal Immune Molecules in Broiler Chickens" Life 12, no. 4: 598. https://doi.org/10.3390/life12040598