
Trehalose Activates Hepatic and Myocardial Autophagy and Has Anti-Inflammatory Effects in db/db Diabetic Mice

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Biochemical Assays
2.4. Differential Counting of Leukocytes
2.5. Expression of IL-10, TNF-α, and Chitinases
2.6. Isolation of RNA from the Liver and Spleen
2.7. cDNA Synthesis
2.8. qPCR
2.9. Morphological Investigations
2.9.1. Light-Microscopic Examination of the Liver and Heart
2.9.2. Transmission Electron Microscopy
2.9.3. Morphometric Electron-Microscopic Analysis
2.9.4. Morphometric Analysis of Lipid Inclusions
2.10. Statistical Analysis
3. Results
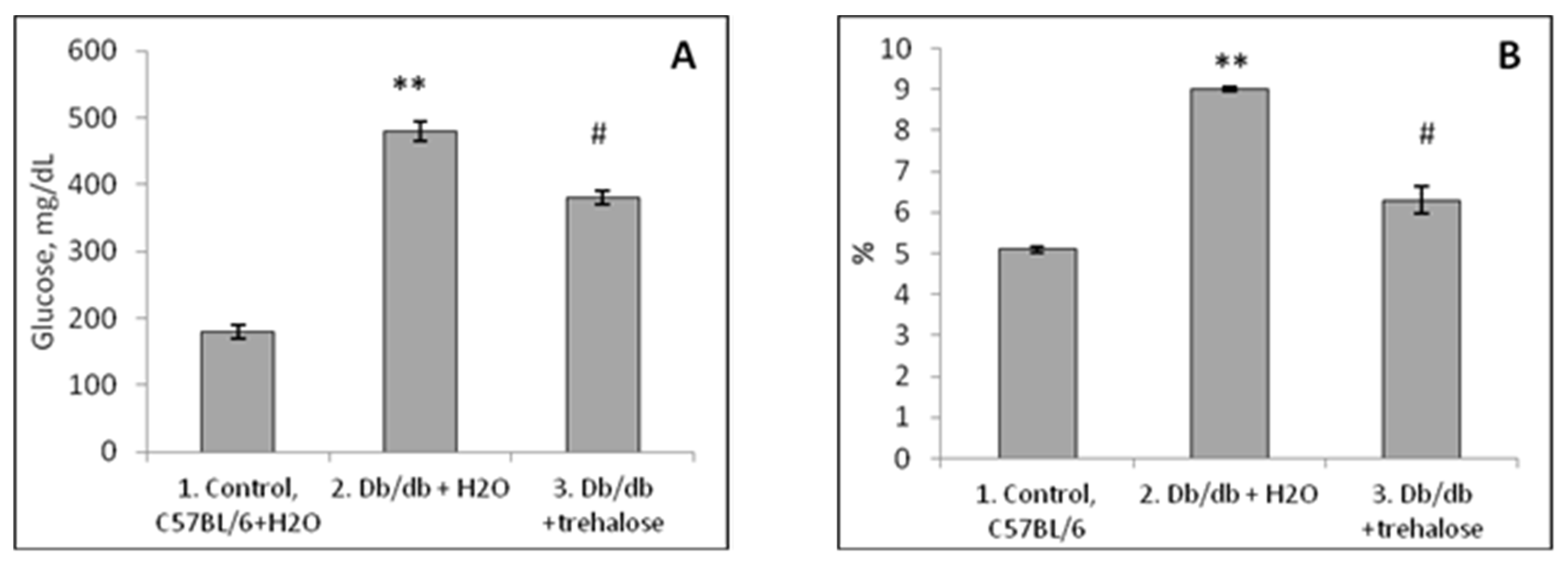
3.1. Blood Leukocyte Characteristics in Db/db Mice
The Impact of Trehalose Treatment on Peripheral-Blood Leukocytes in Db/db Mice
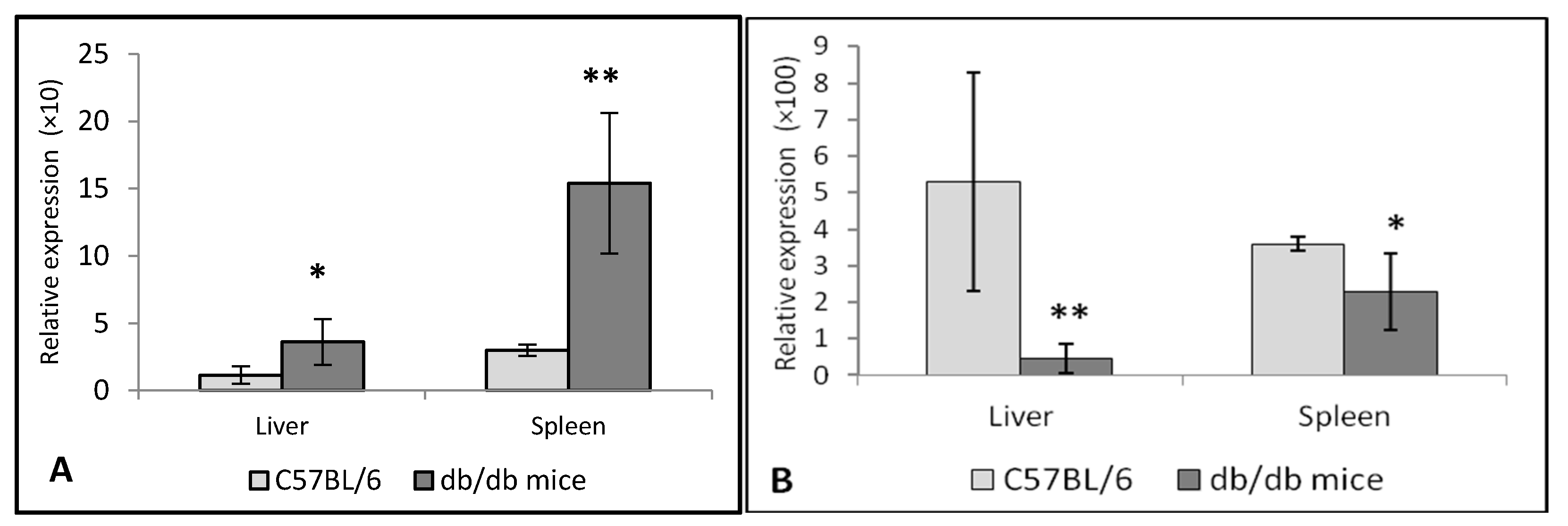
3.2. Expression of TNF-α and IL-10 in the Liver and Spleen of Db/db Mice
3.3. Expression of Chitinases in the Liver and Spleen of Db/db Mice
3.3.1. Chitotriosidase and Acid AMCase in Diabetes
3.3.2. Expression of Chitinases in the Liver and Spleen of Db/db Mice
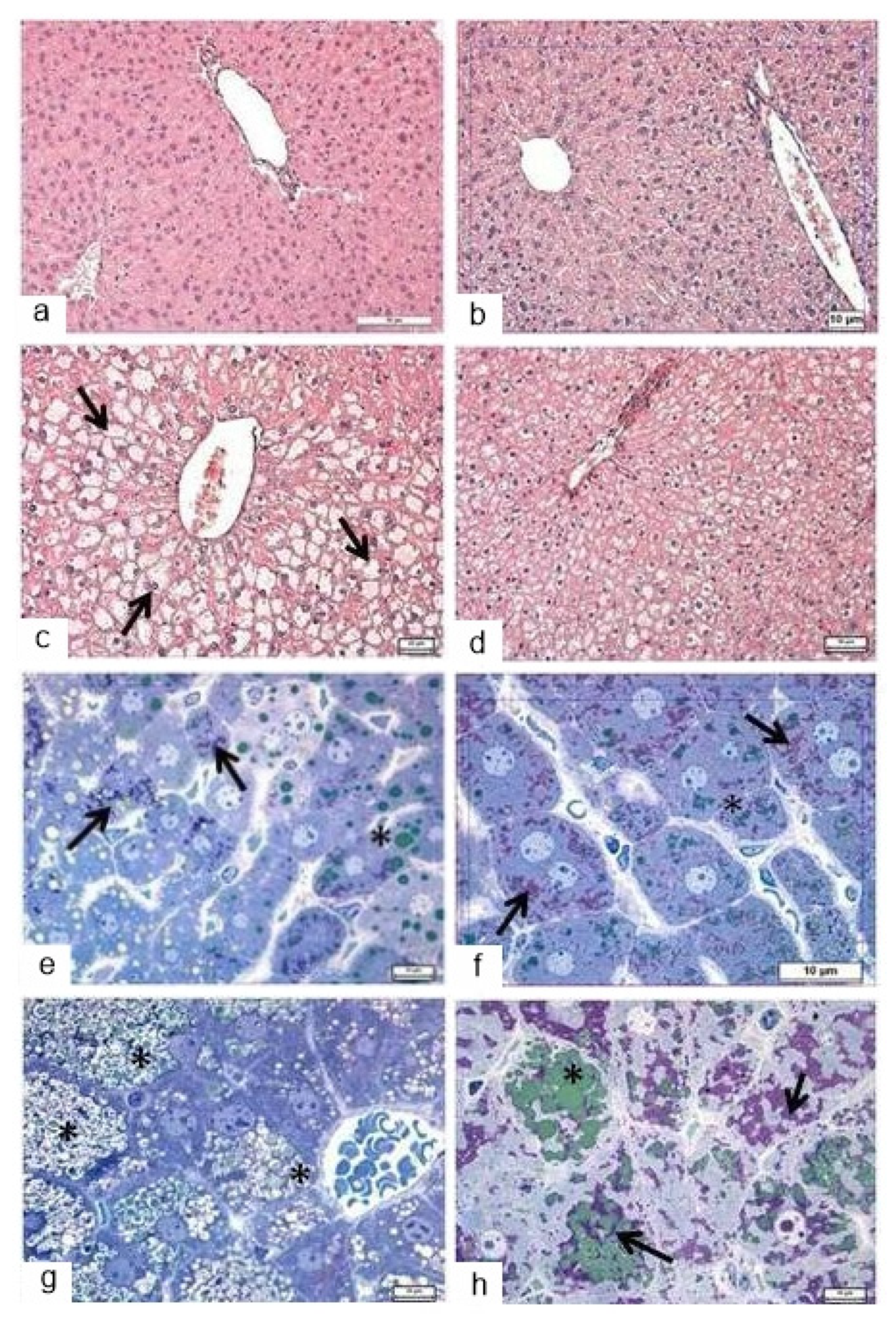
3.4. Morphological Analysis of the Liver in Db/db Mice
3.4.1. Light Microscopy
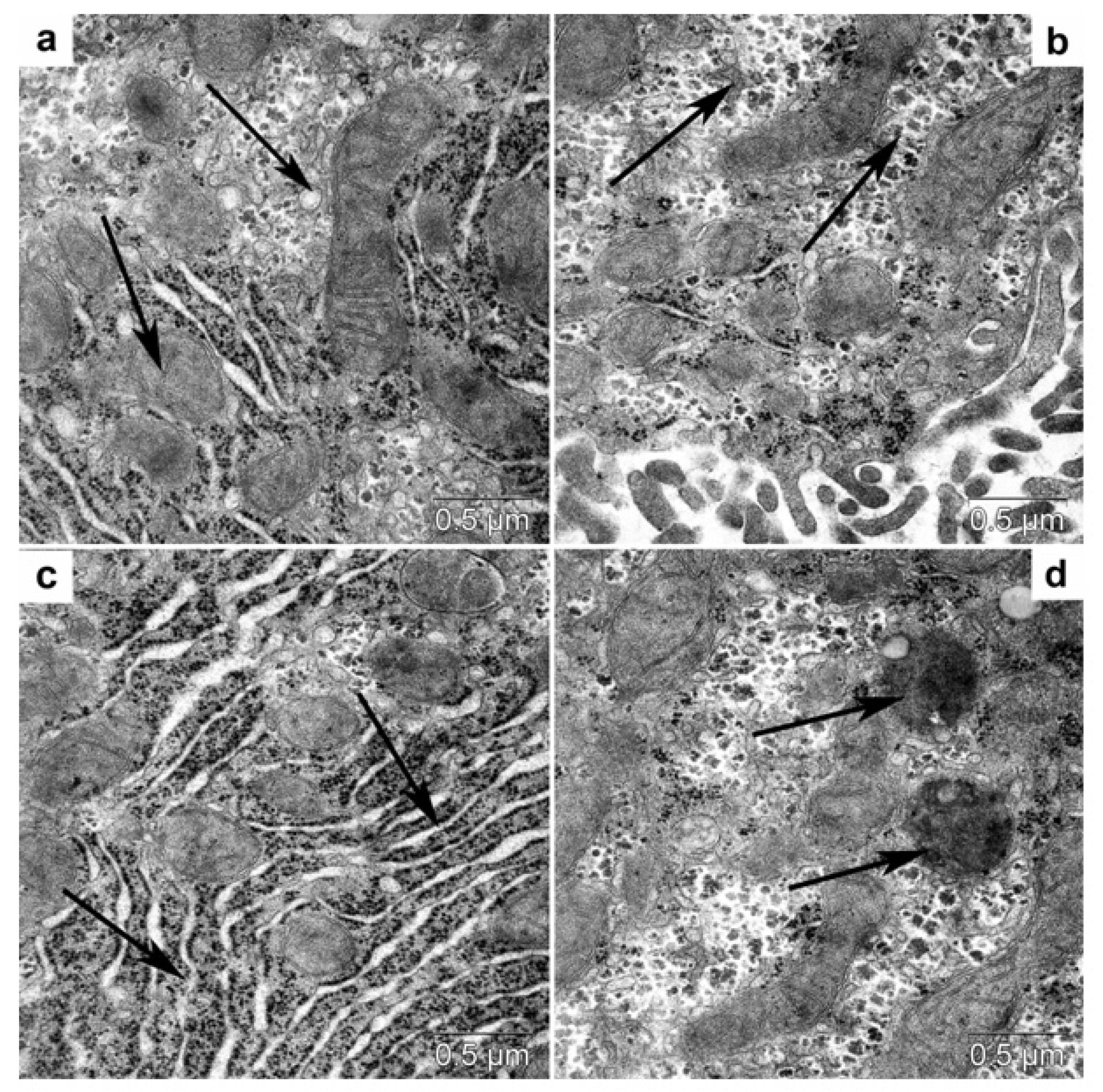
3.4.2. Transmission Electron Microscopy
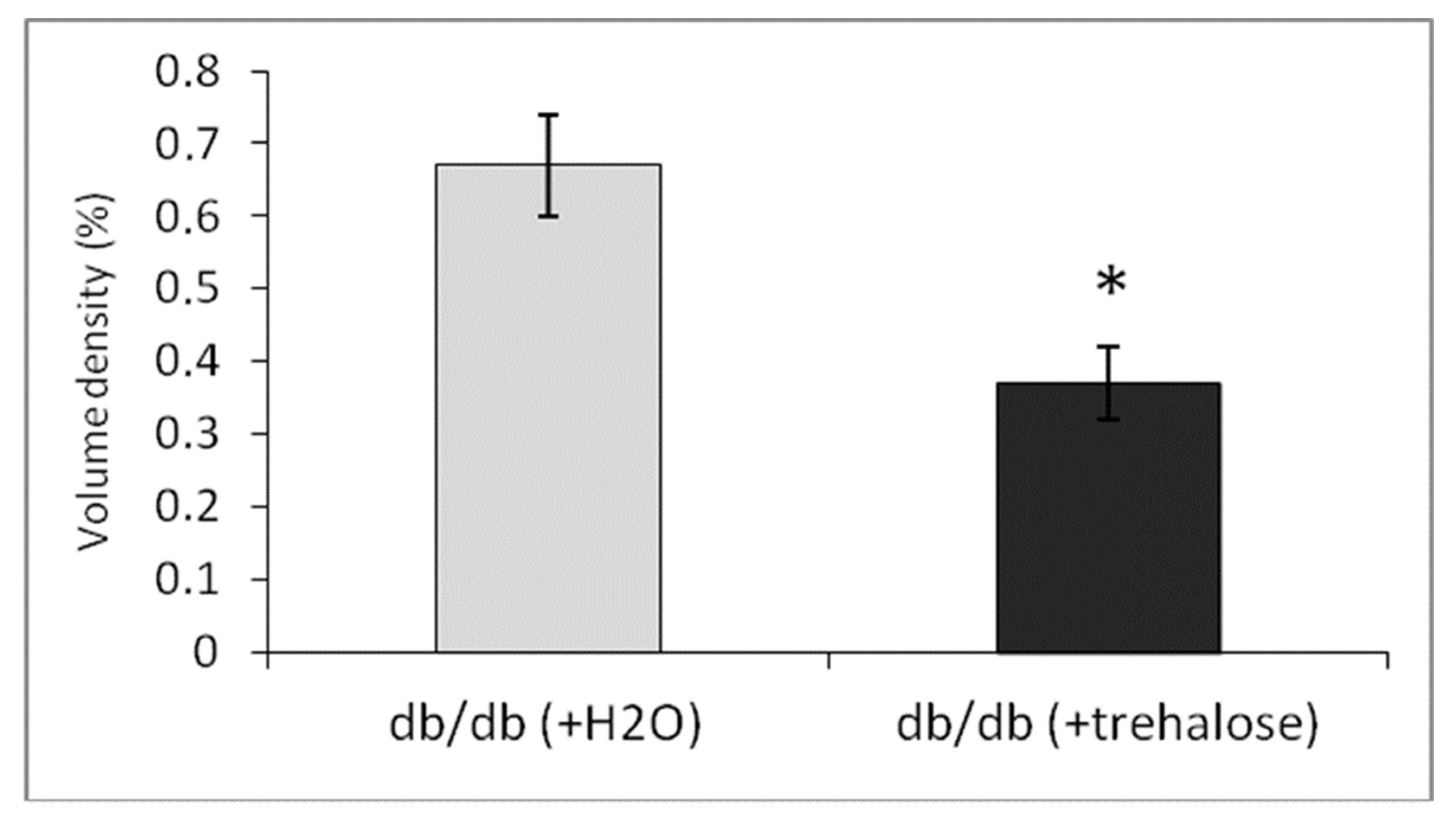
3.4.3. Morphometric Electron-Microscopic Analysis of Hepatocytes
3.5. Morphological Analysis of the Heart
3.5.1. Light Microscopy
3.5.2. Transmission Electron Microscopy
3.5.3. Morphometric Electron-Microscopic Analysis of the Heart
4. Discussion
4.1. The Db/db Mouse Model and Inflammation
4.2. Expression of Chitinases in Db/db Mice
4.3. Expression of Chitinases in Db/db Mice, and Chitinases as Possible Markers of Diabetes
4.4. Possible Mechanisms behind the Protective Effects of Trehalose in Db/db Mice
4.5. The Therapeutic Effect of Trehalose on the Liver and Heart of Db/db Mice
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Pan, X.F.; Chen, J.; Xia, L.; Cao, A.; Zhang, Y.; Wang, J.; Li, H.; Yang, K.; Guo, K.; et al. 2Combined lifestyle factors and risk of incident type 2 diabetes and prognosis among individuals with type 2 diabetes: A systematic review and meta-analysis of prospective cohort studies. Diabetologia 2020, 63, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.X.; Wang, S.H.; Xie, Y.; Chen, Z.T.; Guo, Q.; Yuan, W.L.; Guan, C.; Xu, C.Z.; Huang, Y.N.; Wang, J.F.; et al. Interleukin-33 alleviates diabetic cardiomyopathy through regulation of endoplasmic reticulum stress and autophagy via insulin-like growth factor-binding protein 3. J. Cell. Physiol. 2021, 236, 4403–4419. [Google Scholar] [CrossRef] [PubMed]
- Shan, R.; Liu, N.; Yan, Y.; Liu, B. Apoptosis, autophagy and atherosclerosis: Relationships and the role of Hsp27. Pharmacol. Res. 2021, 166, 105169. [Google Scholar] [CrossRef]
- Khneizer, G.; Rizvi, S.; Gawrieh, S. Non-alcoholic Fatty Liver Disease and Diabetes Mellitus. Adv. Exp. Med. Biol. 2021, 1307, 417–440. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, R.M.; Akpovi, C.D.; Chen, L.; Vitale, M.L. Cholesterol metabolism and Cx43, Cx46, and Cx50 gap junction protein expression and localization in normal and diabetic and obese ob/ob and db/db mouse testes. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E21–E38. [Google Scholar] [CrossRef]
- Lim, H.; Lim, Y.M.; Kim, K.H.; Jeon, Y.E.; Park, K.; Kim, J.; Hwang, H.Y.; Lee, D.J.; Pagire, H.; Kwon, H.J.; et al. A novel autophagy enhancer as a therapeutic agent against metabolic syndrome and diabetes. Nat. Commun. 2018, 9, 1438. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Yaribeygi, A.; Sathyapalan, T.; Sahebkar, A. Molecular mechanisms of trehalose in modulating glucose homeostasis in diabetes. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 2214–2218. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Farrokhi, F.R.; Butler, A.E.; Sahebkar, A. Insulin resistance: Review of the underlining molecular mechanisms. J. Cell. Physiol. 2019, 234, 8152–8161. [Google Scholar] [CrossRef]
- Honma, Y.; Sato-Morita, M.; Katsuki, Y.; Mihara, H.; Baba, R.; Hino, K.; Kawashima, A.; Ariyasu, T.; Harada, M. Trehalose alleviates oxidative stress-mediated liver injury and Mallor-Denk body formation via activating autophagy in mice. Med. Mol. Morphol. 2021, 54, 41–51. [Google Scholar] [CrossRef]
- Korolenko, T.A.; Dubrovina, N.I.; Ovsyukova, M.V.; Bgatova, N.P.; Tenditnik, M.V.; Pupyshev, A.B.; Akopyan, A.S.; Goncharova, N.V.; Lin, C.-L.; Zavjalov, E.L.; et al. Treatment with Autophagy Inducer Trehalose Alleviates Memory and Behavioral Impairments and Neuroinflammatory Brain Processes in db/db Mice. Cells 2021, 10, 2557. [Google Scholar] [CrossRef]
- Forouzanfar, F.; Guest, P.C.; Jamialahmadi, T.; Sahebkar, A. Hepatoprotective Effect of Trehalose: Insight into Its Mechanisms of Action. Adv. Exp. Med. Biol. 2021, 1328, 489–500. [Google Scholar] [CrossRef]
- Ahmed, A.; Khan, T.A.; Dan Ramdath, D.; Kendall, C.W.C.; Sievenpiper, J.L. Rare sugars and their health effects in humans: A systematic review and narrative synthesis of the evidence from human trials. Nutr. Rev. 2022, 80, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Sokołowska, E.; Sadowska, A.; Sawicka, D.; Kotulska-Bąblińska, I.; Car, H. A head-to-head comparison review of biological and toxicological studies of isomaltulose, d-tagatose, and trehalose on glycemic control. Crit. Rev. Food Sci. Nutr. 2021; 1–26, Online ahead of print. [Google Scholar] [CrossRef]
- Khalifeh, M.; Barreto, G.E.; Sahebkar, A. Therapeutic potential of trehalose in neurodegenerative diseases: The knowns and unknowns. Neural Regen. Res. 2021, 16, 2026–2027. [Google Scholar] [CrossRef]
- Jeong, S.J.; Stitham, J.; Evans, T.D.; Zhang, X.; Rodriguez-Velez, A.; Yeh, Y.S.; Tao, J.; Takabatake, K.; Epelman, S.; Lodhi, I.J.; et al. Trehalose causes low-grade lysosomal stress to activate TFEB and the autophagy-lysosome biogenesis response. Autophagy 2021, 17, 3740–3752. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Bettermann, K.; Willing, L.; Sinha, K.; Simpson, I.A. The role of neutrophils in mediating stroke injury in the diabetic db/db mouse brain following hypoxia-ischemia. Neurochem. Int. 2020, 139, 104790. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Su, Z.; Li, Y.; Zhang, X.; You, Q. Chitinase-3 like-protein-1 function and its role in diseases. Signal Transduct. Target. Ther. 2020, 5, 201. [Google Scholar] [CrossRef] [PubMed]
- Wijesekara, N.; Ahrens, R.; Sabale, M.; Wu, L.; Ha, K.; Verdile, G.; Fraser, P.E. Amyloid-β and islet amyloid pathologies link Alzheimer’s disease and type 2 diabetes in a transgenic model. FASEB J. 2017, 31, 5409–5418. [Google Scholar] [CrossRef] [Green Version]
- Lontchi-Yimagou, E.; Sobngwi, E.; Matsha, T.E.; Kengne, A.P. Diabetes mellitus and inflammation. Curr. Diabetes Rep. 2013, 13, 435–444. [Google Scholar] [CrossRef]
- Halim, M.; Halim, A. The effects of inflammation, aging and oxidative stress on the pathogenesis of diabetes mellitus (type 2 diabetes). Diabetes Metab. Syndr. 2019, 13, 1165–1172. [Google Scholar] [CrossRef]
- Klopf, J.; Brostjan, C.; Eilenberg, W.; Neumayer, C. Neutrophil Extracellular Traps and Their Implications in Cardiovascular and Inflammatory Disease. Int. J. Mol. Sci. 2021, 22, 559. [Google Scholar] [CrossRef]
- Tans, R.; van Diepen, J.A.; Bijlsma, S.; Verschuren, L.; Suppers, A.; Stienstra, R.; Wevers, R.A.; Tack, C.J.; Gloerich, J.; van Gool, A.J. Evaluation of chitotriosidase as a biomarker for adipose tissue inflammation in overweight individuals and type 2 diabetic patients. Int. J. Obes. 2019, 43, 1712–1723. [Google Scholar] [CrossRef] [PubMed]
- Tans, R.; Verschuren, L.; Wessels, H.J.C.T.; Bakker, S.J.L.; Tack, C.J.; Gloerich, J.; van Gool, A.J. The future of protein biomarker research in type 2 diabetes mellitus. Expert Rev. Proteom. 2019, 16, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Safina, A.F.; Korolenko, T.A.; Mynkina, G.I.; Dushkin, M.I.; Krasnoselskaya, G.A. Liver and serum lysosomal enzymes activity during zymosan-induced inflammation in mice. Agents Actions Suppl. 1992, 38, 370–375. [Google Scholar] [PubMed]
- Sakitani, K.; Enooku, K.; Kubo, H.; Tanaka, A.; Arai, H.; Kawazu, S.; Koike, K. Clinical characteristics of patients with diabetes mellitus and fatty liver diagnosed by liver/spleen Hounsfield units on CT scan. J. Int. Med. Res. 2017, 45, 1208–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchan, L.; St Aubin, C.R.; Fisher, A.L.; Hellings, A.; Castro, M.; Al-Nakkash, L.; Broderick, T.L.; Plochocki, J.H. High-fat, high-sugar diet induces splenomegaly that is ameliorated with exercise and genistein treatment. BMC Res. Notes 2018, 11, 752. [Google Scholar] [CrossRef] [PubMed]
- Sciarretta, S.; Yee, D.; Nagarajan, N.; Bianchi, F.; Saito, T.; Valenti, V.; Tong, M.; Del Re, D.P.; Vecchione, C.; Schirone, L.; et al. Trehalose-Induced Activation of Autophagy Improves Cardiac Remodeling After Myocardial Infarction. J. Am. Coll. Cardiol. 2018, 71, 1999–2010. [Google Scholar] [CrossRef]
- Elmonem, M.A.; Amin, H.S.; El-Essawy, R.A.; Mehaney, D.A.; Nabil, M.; Kamel, L.N.; Farid, I.M. Association of chitotriosidase enzyme activity and genotype with the risk of nephropathy in type 2 diabetes. Clin. Biochem. 2016, 49, 444–448. [Google Scholar] [CrossRef]
- Rathcke, C.N.; Vestergaard, H. YKL-40—An emerging biomarker in cardiovascular disease and diabetes. Cardiovasc. Diabetol. 2009, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Di Rosa, M.; Malaguarnera, L. Chitotriosidase: A New Inflammatory Marker in Diabetic Complications. Pathobiology 2016, 83, 211–219. [Google Scholar] [CrossRef]
- Kimura, M.; Watanabe, T.; Sekine, K.; Ishizuka, H.; Ikejiri, A.; Sakaguchi, M.; Kamaya, M.; Yamanaka, D.; Matoska, V.; Bauer, P.O.; et al. Comparative functional analysis between human and mouse chitotriosidase: Substitution at amino acid 218 modulates the chitinolytic and transglycosylation activity. Int. J. Biol. Macromol. 2020, 164, 2895–2902. [Google Scholar] [CrossRef]
- Ohno, M.; Tsuda, K.; Sakaguchi, M.; Sugahara, Y.; Oyama, F. Chitinase mRNA levels by quantitative PCR using the single standard DNA: Acidic mammalian chitinase is a major transcript in the mouse stomach. PLoS ONE 2012, 7, e50381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.G.; Da Silva, C.A.; Dela Cruz, C.S.; Ahangari, F.; Ma, B.; Kang, M.J.; He, C.H.; Takyar, S.; Elias, J.A. Role of chitin and chitinase/chitinase-like proteins in inflammation, tissue remodeling, and injury. Annu. Rev. Physiol. 2011, 73, 479–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penttilä, I.; Penttilä, K.; Holm, P.; Laitinen, H.; Ranta, P.; Törrönen, J.; Rauramaa, R. Methods, units and quality requirements for the analysis of haemoglobin A1c in diabetes mellitus. World J. Methodol. 2016, 6, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Korolenko, T.; Johnston, T.P.; Lykov, A.P.; Shintyapina, A.B.; Khrapova, M.V.; Goncharova, N.V.; Korolenko, E.; Bgatova, N.P.; Machova, E.; Nescakova, Z.; et al. A comparative study of the hypolipidaemic effects of a new polysaccharide, mannan Candida albicans serotype A, and atorvastatin in mice with poloxamer 407-induced hyperlipidaemia. J. Pharm. Pharmacol. 2016, 68, 1516–1526. [Google Scholar] [CrossRef]
- Korolenko, T.A.; Johnston, T.P.; Machova, E.; Bgatova, N.P.; Lykov, A.P.; Goncharova, N.V.; Nescakova, Z.; Shintyapina, A.B.; Maiborodin, I.V.; Karmatskikh, O.L. Hypolipidemic effect of mannans from C. albicans serotypes a and B in acute hyperlipidemia in mice. Int. J. Biol. Macromol. 2018, 107, 2385–2394. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Abdelmohsen, K.; Abe, A.; Abedin, M.J.; Abeliovich, H.; Acevedo Arozena, A.; Adachi, H.; Adams, C.M.; Adams, P.D.; Adeli, K.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 2016, 12, 1–222. [Google Scholar] [CrossRef] [Green Version]
- Klinnikova, M.G.; Lushnikova, E.L.; Koldysheva, E.V.; Tolstikova, T.G.; Sorokina, I.V.; Yuzhik, E.I.; Mzhelskaya, M.M. Cardiotoxic and Dyslipidemic Effects of Doxorubicin and Betulinic Acid Amide. Bull. Exp. Biol. Med. 2016, 162, 277–282. [Google Scholar] [CrossRef]
- Żurawska-Płaksej, E.; Ługowska, A.; Hetmańczyk, K.; Knapik-Kordecka, M.; Piwowar, A. Neutrophils as a Source of Chitinases and Chitinase-Like Proteins in Type 2 Diabetes. PLoS ONE 2015, 10, e0141730. [Google Scholar] [CrossRef]
- Żurawska-Płaksej, E.; Kratz, E.M.; Ferens-Sieczkowska, M.; Knapik-Kordecka, M.; Piwowar, A. Changes in glycosylation of human blood plasma chitotriosidase in patients with type 2 diabetes. Glycoconj. J. 2016, 33, 29–39. [Google Scholar] [CrossRef]
- Barad, B.A.; Liu, L.; Diaz, R.E.; Basilio, R.; Van Dyken, S.J.; Locksley, R.M.; Fraser, J.S. Differences in the chitinolytic activity of mammalian chitinases on soluble and insoluble substrates. Protein Sci. 2020, 29, 966–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Chen, H.; Gao, J.; Liu, Y.; Li, J.; Wang, J. Molecular machinery and interplay of apoptosis and autophagy in coronary heart disease. J. Mol. Cell. Cardiol. 2019, 136, 27–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillon, N.J.; Loos, R.J.F.; Marshall, S.M.; Zierath, J.R. Metabolic consequences of obesity and type 2 diabetes: Balancing genes and environment for personalized care. Cell 2021, 184, 1530–1544. [Google Scholar] [CrossRef] [PubMed]
- Bowker, N.; Shah, R.L.; Sharp, S.J.; Luan, J.; Stewart, I.D.; Wheeler, E.; Ferreira, M.A.R.; Baras, A.; Wareham, N.J.; Langenberg, C.; et al. Meta-analysis investigating the role of interleukin-6 mediated inflammation in type 2 diabetes. EBioMedicine 2020, 61, 103062. [Google Scholar] [CrossRef] [PubMed]
- Akash, M.S.H.; Rehman, K.; Liaqat, A. Tumor Necrosis Factor-Alpha: Role in Development of Insulin Resistance and Pathogenesis of Type 2 Diabetes Mellitus. J. Cell. Biochem. 2018, 119, 105–110. [Google Scholar] [CrossRef]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Chen, X.; Chen, H. Association of Single Nucleotide Polymorphisms of the IL-6, IL-10, and TNF-α Genes with Susceptibility to Gestational Diabetes Mellitus. Genet. Test. Mol. Biomark. 2020, 24, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, L.; Qiao, L.; Hu, S.; Ge, J.; Hu, C.; Li, T. Combination therapy with anti-CD20 mAb and IL-10 gene to reverse type 1 diabetes by attenuating pancreatitis and inhibiting apoptosis in NOD mice. Life Sci. 2020, 256, 117985. [Google Scholar] [CrossRef]
- Lehrskov, L.L.; Christensen, R.H. The role of interleukin-6 in glucose homeostasis and lipid metabolism. Semin. Immunopathol. 2019, 41, 491–499. [Google Scholar] [CrossRef]
- Kristiansen, O.P.; Mandrup-Poulsen, T. Interleukin-6 and diabetes: The good, the bad, or the indifferent? Diabetes 2005, 54 (Suppl. 2), S114–S124. [Google Scholar] [CrossRef] [Green Version]
- Mazur, M.; Zielińska, A.; Grzybowski, M.M.; Olczak, J.; Fichna, J. Chitinases and Chitinase-Like Proteins as Therapeutic Targets in Inflammatory Diseases, with a Special Focus on Inflammatory Bowel Diseases. Int. J. Mol. Sci. 2021, 22, 6966. [Google Scholar] [CrossRef] [PubMed]
- Park, B.J.; Yoon, Y.B.; Lee, D.H.; Shin, C.; Juakali, L.; Cho, S.J.; Park, S.C. Transcriptional upregulation of multiple earthworm chitinase genes following bacterial challenge suggests their implications in innate immunity. Genes Genom. 2021, 43, 1497–1502. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Rabach, M.; Chen, N.Y.; Rabach, L.; Hu, X.; Elias, J.A.; Zhu, Z. Molecular cloning and functional characterization of mouse chitotriosidase. Gene 2005, 357, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Fujimoto, W.; Goto, M.; Morimatsu, M.; Syuto, B.; Iwanaga, T. Cellular expression of gut chitinase mRNA in the gastrointestinal tract of mice and chickens. J. Histochem. Cytochem. 2002, 50, 1081–1089. [Google Scholar] [CrossRef] [Green Version]
- Malaguarnera, L.; Di Rosa, M.; Zambito, A.M.; dell’Ombra, N.; Nicoletti, F.; Malaguarnera, M. Chitotriosidase gene expression in Kupffer cells from patients with non-alcoholic fatty liver disease. Gut 2006, 55, 1313–1320. [Google Scholar] [CrossRef]
- Hu, C.; Ma, Z.; Zhu, J.; Fan, Y.; Tuo, B.; Li, T.; Liu, X. Physiological and pathophysiological roles of acidic mammalian chitinase (CHIA) in multiple organs. Biomed. Pharmacother. 2021, 138, 111465. [Google Scholar] [CrossRef]
- Yap, J.; McCurdy, S.; Alcala, M.; Irei, J.; Garo, J.; Regan, W.; Lee, B.H.; Kitamoto, S.; Boisvert, W.A. Expression of Chitotriosidase in Macrophages Modulates Atherosclerotic Plaque Formation in Hyperlipidemic Mice. Front. Physiol. 2020, 23, 714. [Google Scholar] [CrossRef]
- Pinteac, R.; Montalban, X.; Comabella, M. Chitinases and chitinase-like proteins as biomarkers in neurologic disorders. Neurol. Neuroimmunol. Neuroinflamm. 2020, 8, e921. [Google Scholar] [CrossRef]
- Kitamoto, S.; Egashira, K.; Ichiki, T.; Han, X.; McCurdy, S.; Sakuda, S.; Sunagawa, K.; Boisvert, W.A. Chitinase inhibition promotes atherosclerosis in hyperlipidemic mice. Am. J. Pathol. 2013, 183, 313–325. [Google Scholar] [CrossRef] [Green Version]
- Artieda, M.; Cenarro, A.; Gañán, A.; Jericó, I.; Gonzalvo, C.; Casado, J.M.; Vitoria, I.; Puzo, J.; Pocoví, M.; Civeira, F. Serum chitotriosidase activity is increased in subjects with atherosclerosis disease. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1645–1652. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Li, X.; Wang, F.; Weng, H.; Yang, P. Trehalose prevents neural tube defects by correcting maternal diabetes-suppressed autophagy and neurogenesis. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E667–E678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Lim, Y.M.; Lee, M.S. The role of autophagy in systemic metabolism and human-type diabetes. Mol. Cells 2018, 41, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Cutaș, A.; Drugan, C.; Roman, G.; Rusu, A.; Cătană, C.S.; Achimaș-Cadariu, A.; Drugan, T. Evaluation of Chitotriosidase and Neopterin as Biomarkers of Microvascular Complications in Patients with Type 1 Diabetes Mellitus. Diagnostics 2021, 11, 263. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Raymond, K. Beta-glucans in the treatment of diabetes and associated cardiovascular risks. Vasc. Health Risk Manag. 2008, 4, 1265–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pupyshev, A.B.; Tikhonova, M.A.; Akopyan, A.A.; Tenditnik, M.V.; Dubrovina, N.I.; Korolenko, T.A. Therapeutic activation of autophagy by combined treatment with rapamycin and trehalose in a mouse MPTP-induced model of Parkinson’s disease. Pharmacol. Biochem. Behav. 2019, 177, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Korolenko, T.A.; Johnston, T.P.; Vetvicka, V. Lysosomotropic features and autophagy modulators among medical drugs: Evaluation of their role in pathologies. Molecules 2020, 25, 5052. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.K.; Kwon, Y.; Byeon, S.; Lee, Y.H. Stimulation of autophagy improves vascular function in the mesenteric arteries of type 2 diabetic mice. Exp. Physiol. 2020, 105, 192–200. [Google Scholar] [CrossRef]
- Abdellatif, M.; Ljubojevic-Holzer, S.; Madeo, F.; Sedej, S. Autophagy in cardiovascular health and disease. Prog. Mol. Biol. Transl. Sci. 2020, 172, 87–106. [Google Scholar] [CrossRef] [PubMed]
- Mack, C.I.; Ferrario, P.G.; Weinert, C.H.; Egert, B.; Hoefle, A.S.; Lee, Y.M.; Skurk, T.; Kulling, S.E.; Daniel, H. Exploring the Diversity of Sugar Compounds in Healthy, Prediabetic, and Diabetic Volunteers. Mol. Nutr. Food Res. 2020, 64, e1901190. [Google Scholar] [CrossRef]
- Gao, G.; Chen, W.; Yan, M.; Liu, J.; Luo, H.; Wang, C.; Yang, P. Rapamycin regulates the balance between cardiomyocyte apoptosis and autophagy in chronic heart failure by inhibiting mTOR signaling. Int. J. Mol. Med. 2020, 45, 195–209. [Google Scholar] [CrossRef] [Green Version]
- Stachowicz, A.; Wiśniewska, A.; Kuś, K.; Kiepura, A.; Gębska, A.; Gajda, M.; Białas, M.; Totoń-Żurańska, J.; Stachyra, K.; Suski, M.; et al. The Influence of Trehalose on Atherosclerosis and Hepatic Steatosis in Apolipoprotein E Knockout Mice. Int. J. Mol. Sci. 2019, 20, 1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Zhang, K.; Wang, H.; Wu, Y.; Chen, N.; Chen, J.; Qiu, C.; Cai, P.; Li, M.; Liang, X.; et al. Hrd1-mediated ACLY ubiquitination alleviate NAFLD in db/db mice. Metabolism 2021, 114, 154349. [Google Scholar] [CrossRef] [PubMed]
- Noratto, G.D.; Murphy, K.; Chew, B.P. Quinoa intake reduces plasma and liver cholesterol, lessens obesity-associated inflammation, and helps to prevent hepatic steatosis in obese db/db mouse. Food Chem. 2019, 287, 107–114. [Google Scholar] [CrossRef]
- Omori, K.; Nakamura, A.; Miyoshi, H.; Takahashi, K.; Kitao, N.; Nomoto, H.; Kameda, H.; Cho, K.Y.; Takagi, R.; Hatanaka, K.C.; et al. Effects of dapagliflozin and/or insulin glargine on beta cell mass and hepatic steatosis in db/db mice. Metabolism 2019, 98, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.G.; Wang, X.Y.; Wang, J.H.; Fan, R.F.; Wang, L. Trehalose prevents cadmium-induced hepatotoxicity by blocking Nrf2 pathway, restoring autophagy and inhibiting apoptosis. J. Inorg. Biochem. 2019, 192, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Schulze, R.J.; McNiven, M.A. Lipid Droplet Formation and Lipophagy in Fatty Liver Disease. Semin. Liver Dis. 2019, 39, 283–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kounakis, K.; Chaniotakis, M.; Markaki, M.; Tavernarakis, N. Emerging Roles of Lipophagy in Health and Disease. Front. Cell Dev. Biol. 2019, 7, 185. [Google Scholar] [CrossRef] [PubMed]
- Chao, X.; Wang, S.; Yang, L.; Ni, H.M.; Ding, W.X. Trehalose activates hepatic transcription factor EB (TFEB) but fails to ameliorate alcohol-impaired TFEB and liver injury in mice. Alcohol Clin. Exp. Res. 2021, 45, 1950–1964. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Oligonucleotide Sequence |
|---|---|
| Mus musculus EEF2 | forward 5′-GGAGACCGGTACTTTGATCC-3′ |
| reverse 5′-TTTGTCCTCGCTGTCCAGCT-3′ | |
| Mus musculus TBP | forward 5′-AGGAGCCAAGAGTGAAGAACAA-3′ |
| reverse 5′-ATAATTCTGGCTCATAGCTACTGA-3′ | |
| Mus musculus CHIA | forward 5′-TCCTGGTGAAGGAAATGCGT-3′ |
| reverse 5′-AAATCCCACCAGCTACAGCA-3′ | |
| Mus musculus CHIT1 | forward 5′-TCAGACAATGGAGTTGGGGC-3′ |
| reverse 5′-TTCCAGGAGCAGGCCTCATA-3′ | |
| Mus musculus IL-10 | forward 5′-TGGGTTGCCAAGCCTTATCG-3′ |
| reverse 5′-CTCTTCACCTGCTCCACTGC-3′ | |
| Mus musculus TNF-α | forward 5′-TCAGTTCTATGGCCCAGACC-3′ |
| reverse 5′-ACCACTAGTTGGTTGTCTTTGAG-3′ |
| Groups of Mice | Autophagosome, Vv | Autolysosome, Vv | Lysosome, Vv |
|---|---|---|---|
| db/db + H2O | 0.67 ± 0.18 | 0.4 ± 0.21 | 1.31 ± 0.97 |
| db/db + trehalose | 0.91 ± 0.32 | 1.26 ± 0.99 * | 1.56 ± 0.26 |
| Groups of Mice | Autophagosomes, Vv | Autolysosomes, Vv | Lysosomes, Vv |
|---|---|---|---|
| db/db + H2O | 2.58 ± 0.41 | 0.99 ± 0.32 | 0.01 ± 0.008 |
| db/db + trehalose | 2.28 ± 0.30 | 2.07 ± 0.34 * | 0.02 ± 0.009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korolenko, T.A.; Ovsyukova, M.V.; Bgatova, N.P.; Ivanov, I.D.; Makarova, S.I.; Vavilin, V.A.; Popov, A.V.; Yuzhik, E.I.; Koldysheva, E.V.; Korolenko, E.C.; et al. Trehalose Activates Hepatic and Myocardial Autophagy and Has Anti-Inflammatory Effects in db/db Diabetic Mice. Life 2022, 12, 442. https://doi.org/10.3390/life12030442
Korolenko TA, Ovsyukova MV, Bgatova NP, Ivanov ID, Makarova SI, Vavilin VA, Popov AV, Yuzhik EI, Koldysheva EV, Korolenko EC, et al. Trehalose Activates Hepatic and Myocardial Autophagy and Has Anti-Inflammatory Effects in db/db Diabetic Mice. Life. 2022; 12(3):442. https://doi.org/10.3390/life12030442
Chicago/Turabian StyleKorolenko, Tatiana A., Marina V. Ovsyukova, Nataliya P. Bgatova, Igor D. Ivanov, Svetlana I. Makarova, Valentin A. Vavilin, Alexey V. Popov, Ekaterina I. Yuzhik, Elena V. Koldysheva, Erik C. Korolenko, and et al. 2022. "Trehalose Activates Hepatic and Myocardial Autophagy and Has Anti-Inflammatory Effects in db/db Diabetic Mice" Life 12, no. 3: 442. https://doi.org/10.3390/life12030442