
Ethyl P-Methoxycinnamate: An Active Anti-Metastasis Agent and Chemosensitizer Targeting NFκB from Kaempferia galanga for Melanoma Cells

 , ,
, , 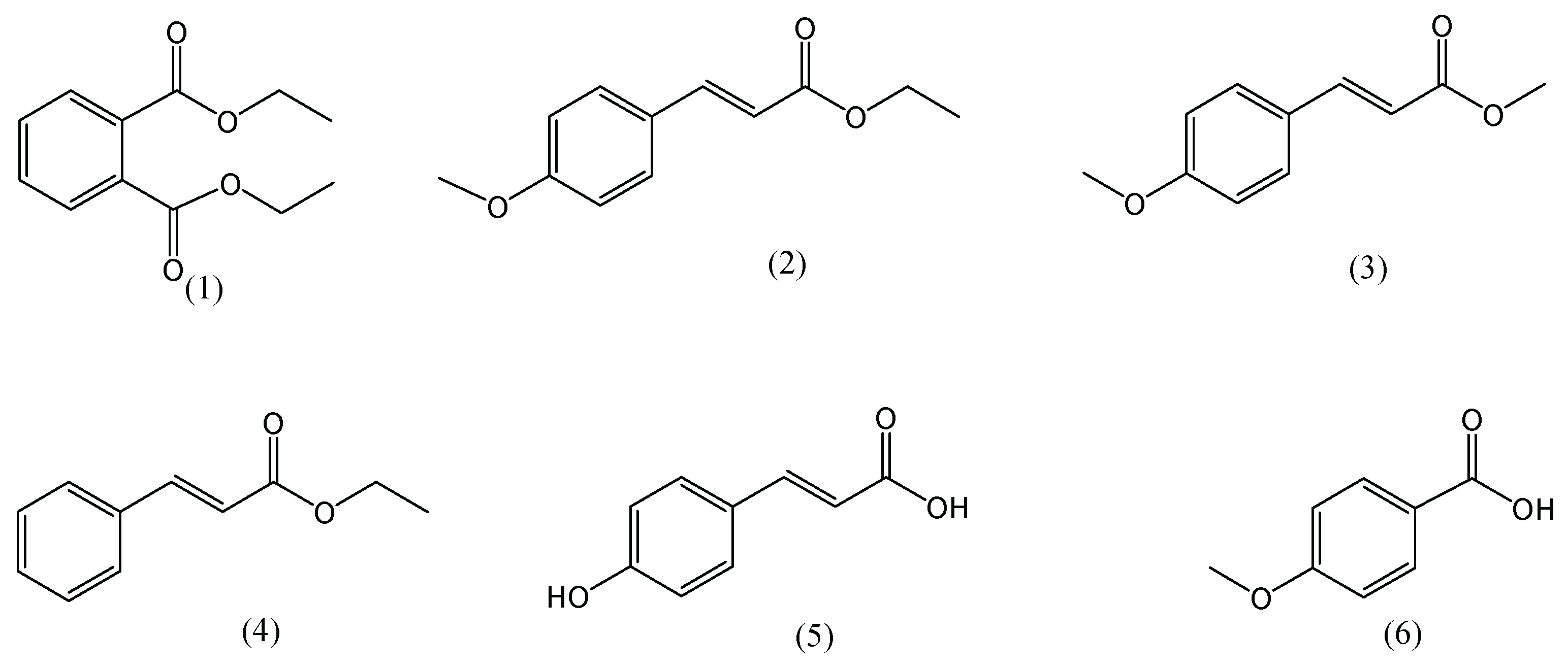{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Extracts and Compounds
2.2. Cells and Reagents
2.3. Cell Viability
2.4. NFκB Reporter Gene Assay
2.5. Wound Healing Assay
2.6. Transwell Invasion Assay
2.7. Western Blotting
3. Results
3.1. Active Compounds of Kaempferia galanga
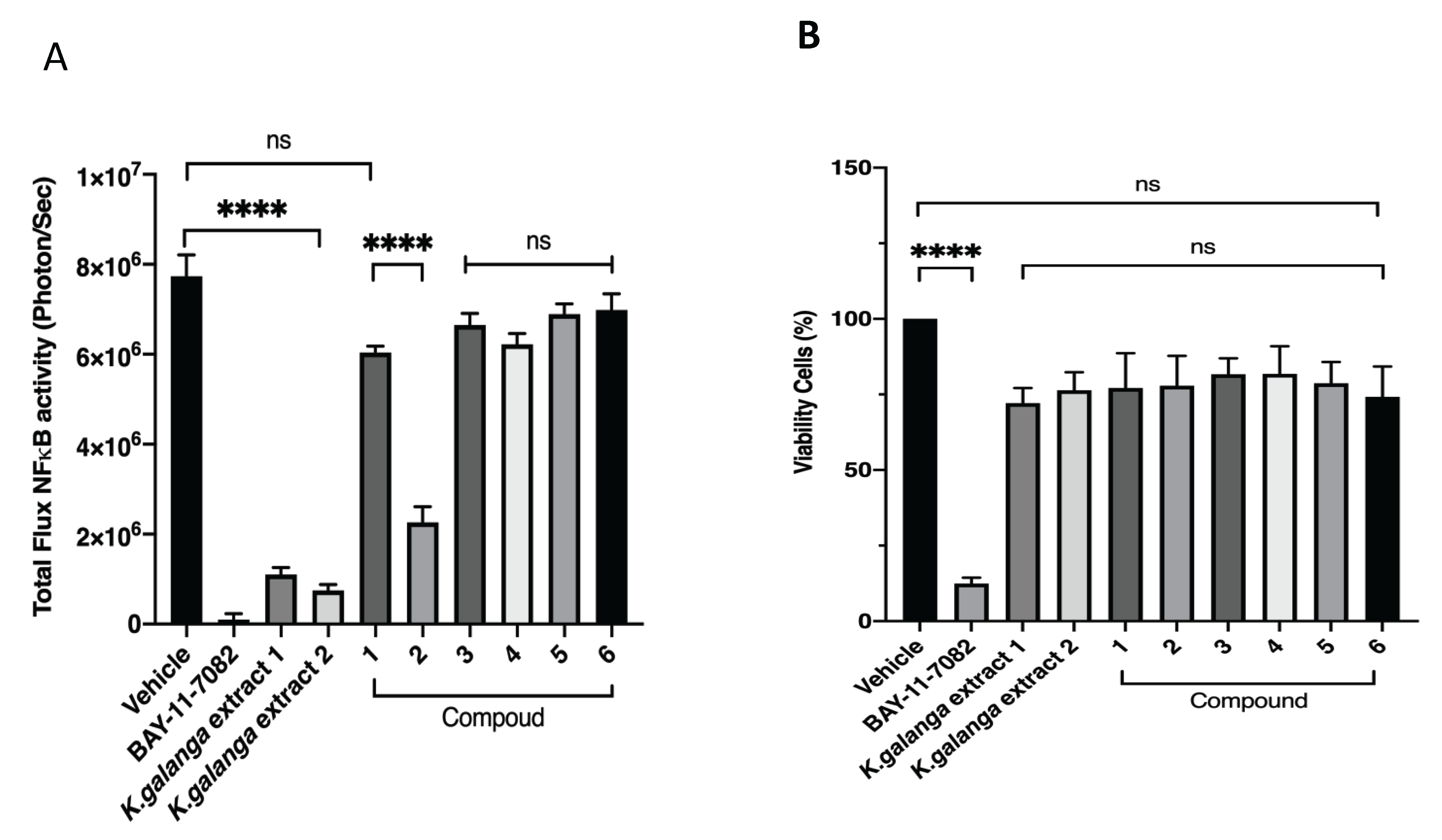
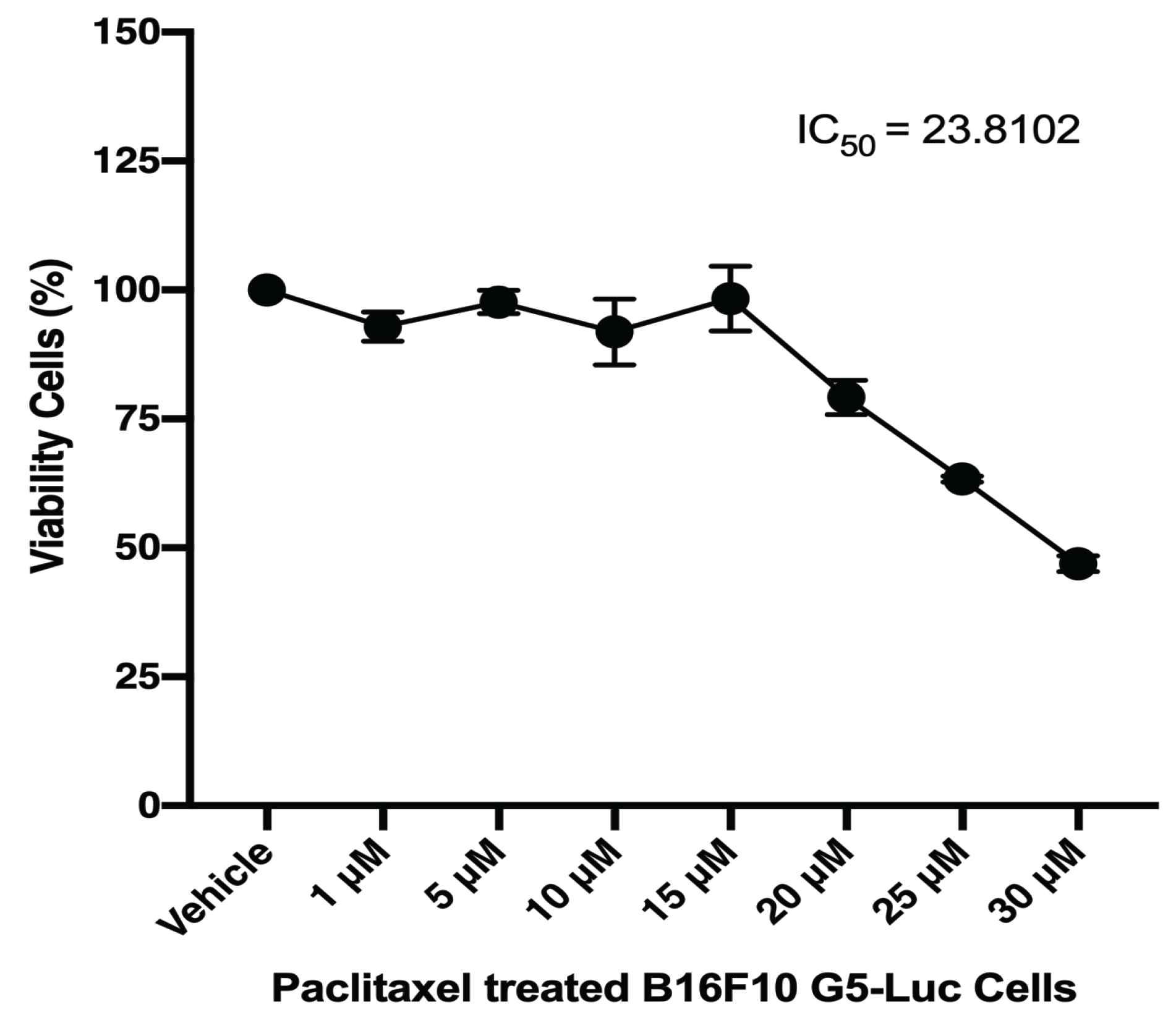
3.2. NFκB Inhibition and Viability Cell against B16F10 Melanoma Cell
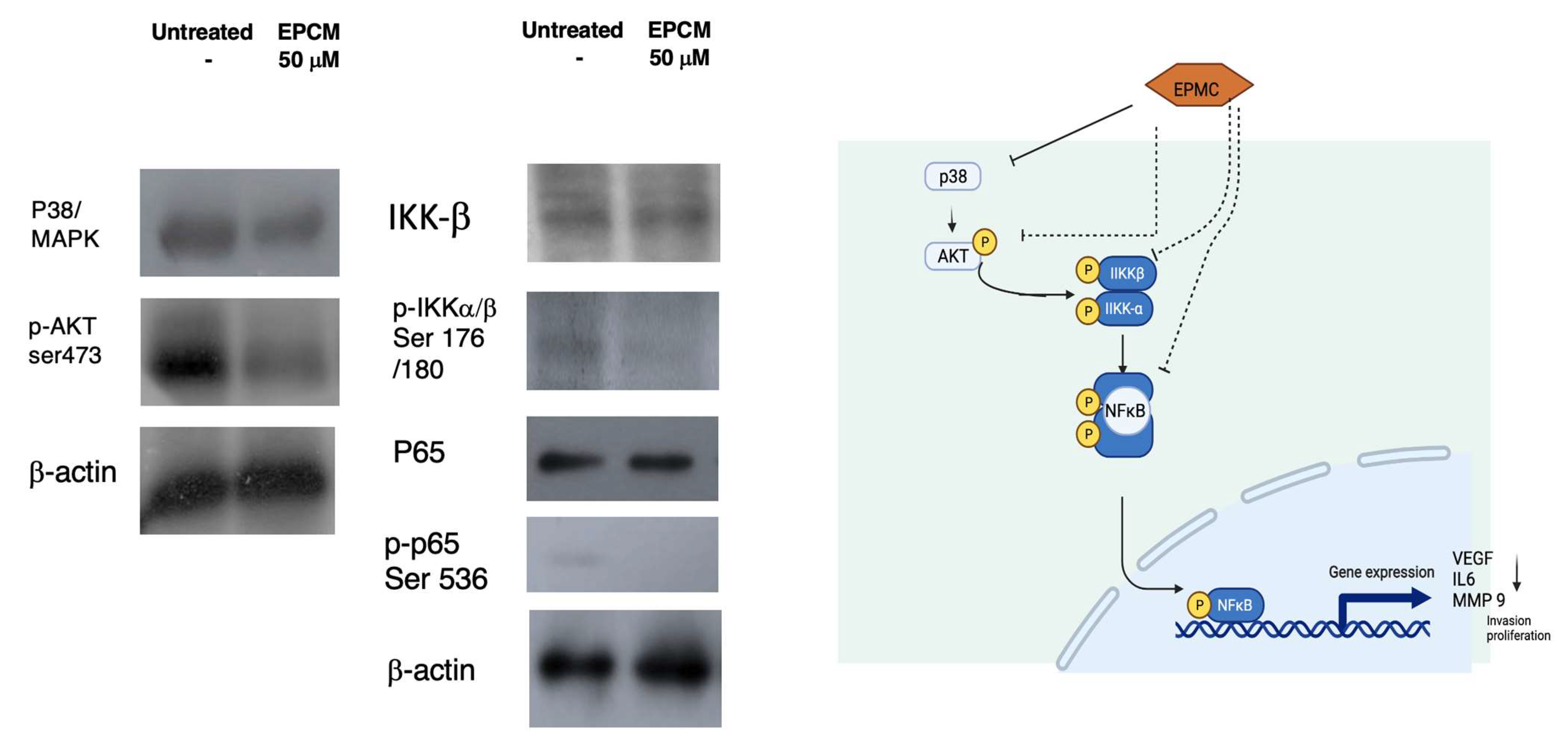
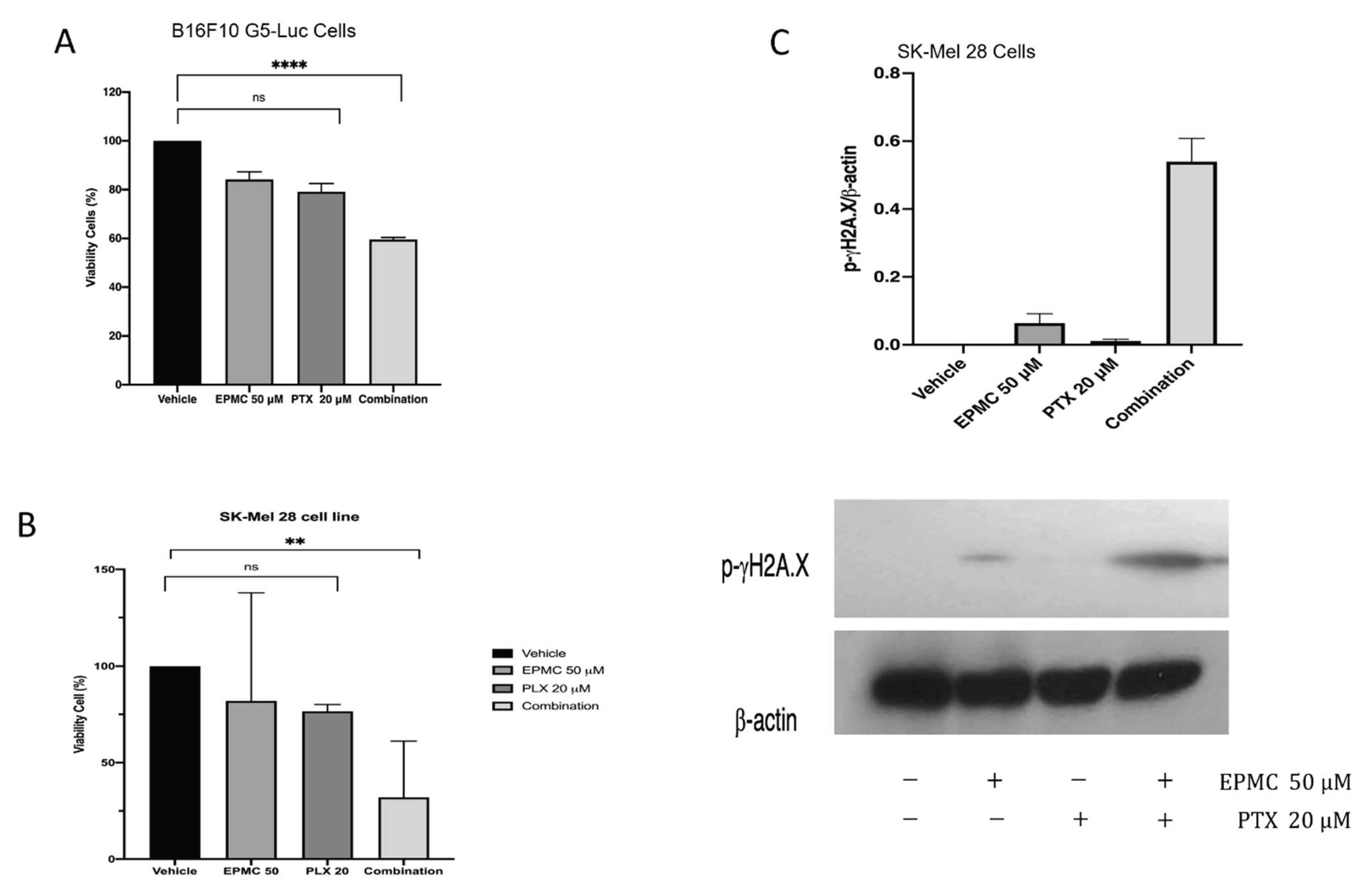
3.3. Western Blotting Analysis
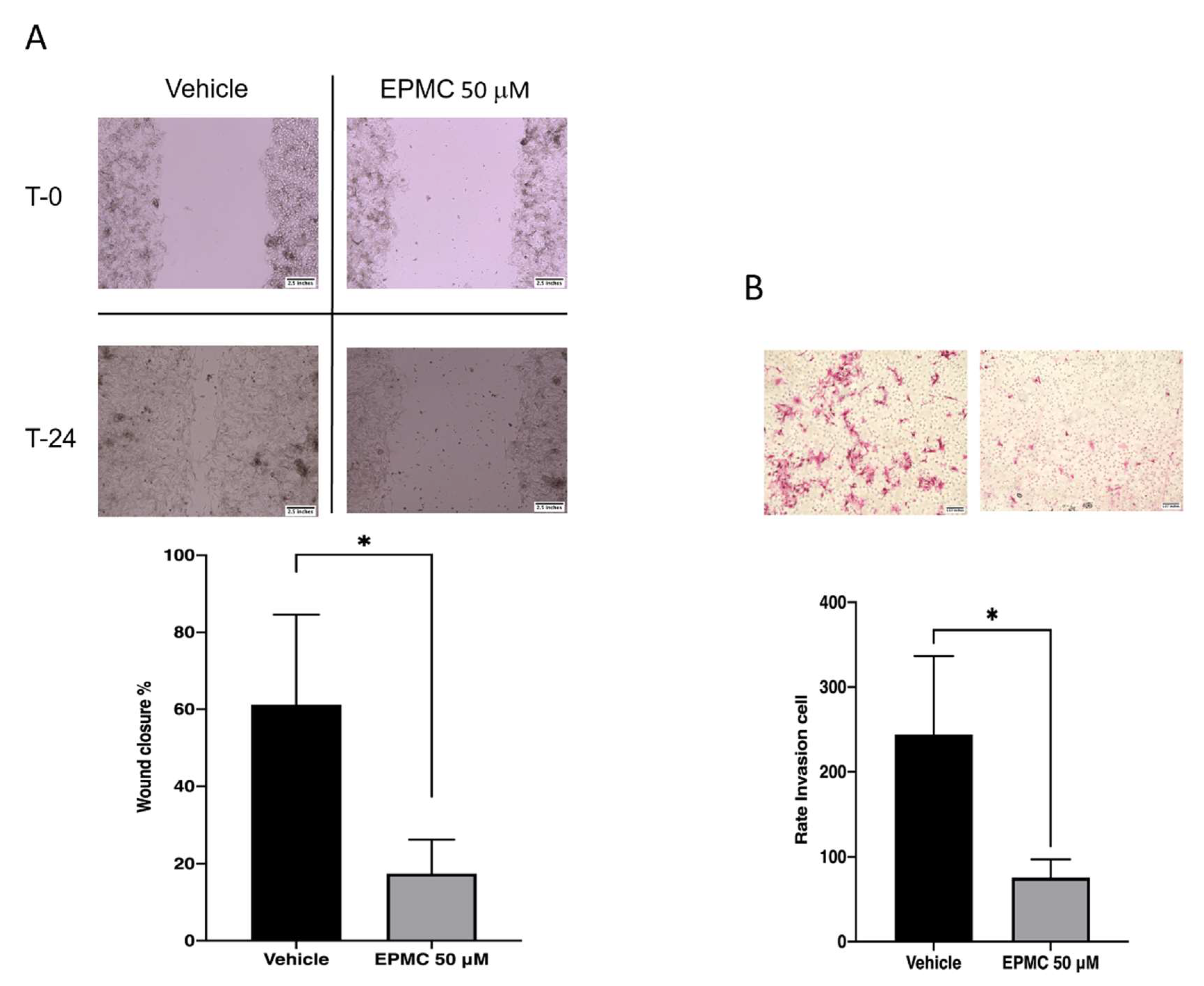
3.4. Migration Assay
3.5. Sensitivity Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chin, Y.W.; Balunas, M.J.; Chai, H.B.; Kinghorn, A.D. Drug discovery from natural sources. AAPS J. 2006, 8, E239–E253. [Google Scholar] [CrossRef] [Green Version]
- Jirovetz, L.; Buchbauer, G.; Shafi, P.M.; Abraham, G.T. Analysis of the essential oil of the roots of the medicinal plant Kaempferia galanga L. (Zingiberaceae) from South-India. Act. Pharm. Turc. 2001, 43, 107–110. [Google Scholar]
- Lallo, S.; Hasmiranti, A.; Hardianti, B. Effect of the growth environment on Patchouli (Pogostemon cablin Benth.) oil character at Southeast Sulawesi, Indonesia. Med. Plants Int. J. Phytomed. Relat. Ind. 2020, 12, 77–81. [Google Scholar] [CrossRef]
- Khairullah, A.R.; Solikhah, T.I.; Ansori, A.N.M.; Hanisia, R.H.; Puspitarani, G.A.; Fadholly, A.; Ramandinianto, S.C. Medicinal importance of Kaempferia galanga L. (Zingiberaceae): A comprehensive review. J. Herbmed Pharmacol. 2021, 10, 281–288. [Google Scholar] [CrossRef]
- Elshamy, A.I.; Mohamed, T.A.; Essa, A.F.; Gawad, A.-E.; Ahmed, M.; Alqahtani, A.S.; Shahat, A.A.; Yoneyama, T.; Farrag, A.R.H.; Noji, M. Recent advances in Kaempferia phytochemistry and biological activity: A comprehensive review. Nutrients 2019, 11, 2396. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, N.; Singh, S.; Gupta, A.C.; Shanker, K.; Bawankule, D.U.; Luqman, S. Aromatic ginger (Kaempferia galanga L.) extracts with ameliorative and protective potential as a functional food, beyond its flavor and nutritional benefits. Toxicol. Rep. 2019, 6, 521–528. [Google Scholar] [CrossRef]
- Jagadish, P.C.; Latha, K.P.; Mudgal, J.; Nampurath, G.K. Extraction, characterization and evaluation of Kaempferia galanga L. (Zingiberaceae) rhizome extracts against acute and chronic inflammation in rats. J. Ethnopharmacol. 2016, 194, 434–439. [Google Scholar] [CrossRef]
- He, Z.-H.; Yue, G.G.-L.; Lau, C.B.-S.; Ge, W.; But, P.P.-H. Antiangiogenic effects and mechanisms of trans-ethyl p-Methoxycinnamate from Kaempferia galanga L. J. Agric. Food Chem. 2012, 60, 11309–11317. [Google Scholar] [CrossRef]
- Amuamuta, A.; Plengsuriyakarn, T.; Na-Bangchang, K. Anticholangiocarcinoma activity and toxicity of the Kaempferia galanga Linn. rhizome ethanolic extract. BMC Complement. Altern. Med. 2017, 17, 213. [Google Scholar] [CrossRef]
- Kumar, A. Phytochemistry, pharmacological activities and uses of traditional medicinal plant Kaempferia galanga L.—An overview. J. Ethnopharmacol. 2020, 253, 112667. [Google Scholar] [CrossRef]
- Shetu, H.J.; Trisha, K.T.; Sikta, S.A.; Anwar, R.; Rashed, S.S.B.; Dash, P.R. Pharmacological importance of Kaempferia galanga (Zingiberaceae): A mini review. Int. J. Res. Pharm. Pharm. Sci. 2018, 3, 32–39. [Google Scholar]
- Xia, Y.; Shen, S.; Verma, I.M. NF-κB, an active player in human cancers. Cancer Immunol. Res. 2014, 2, 823–830. [Google Scholar] [CrossRef] [Green Version]
- Lallo, S.; Hardianti, B.; Sartini, S.; Ismail, I.; Hayakawa, Y. Anti-inflamatory and cytoprotective effect of Kaempferia galanga extracts by targeting NFκB activity. Asian J. Plant Sci. 2022, 21, 183–191. [Google Scholar]
- Takahashi, K.; Takeda, K.; Saiki, I.; Irimura, T.; Hayakawa, Y. Functional roles of tumor necrosis factor-related apoptosis-inducing ligand-DR5 interaction in B16F10 cells by activating the nuclear factor-κB pathway to induce metastatic potential. Cancer Sci. 2013, 104, 558–562. [Google Scholar] [CrossRef] [Green Version]
- Yarrow, J.C.; Perlman, Z.E.; Westwood, N.J.; Mitchison, T.J. A high-throughput cell migration assay using scratch wound healing, a comparison of image-based readout methods. BMC Biotechnol. 2004, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Jonkman, J.E.; Cathcart, J.A.; Xu, F.; Bartolini, M.E.; Amon, J.E.; Stevens, K.M.; Colarusso, P. An introduction to the wound healing assay using live-cell microscopy. Cell Adh. Migr. 2014, 8, 440–451. [Google Scholar] [CrossRef] [Green Version]
- Shin, M.K.; Sasaki, F.; Ki, D.W.; Win, N.N.; Morita, H.; Hayakawa, Y. Anti-metastatic effects of ergosterol peroxide from the entomophatogenic fungus Ophiocordyceps gracilioides on 4T1 breast cancer cells. J. Nat. Med. 2021, 75, 824–832. [Google Scholar] [CrossRef]
- Keire, D.A.; Anto, P.; Faull, K.F.; Ruth, E.; Walsh, J.H.; Chew, P.; Quisimoro, D.; Territo, M.; Reeve, J.R., Jr. Diethyl phatalate, a chemotactic factor secreted by Helicobacter pylori. J. Biol. Chem. 2001, 276, 48847–48853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasali, N.H.M.; Umar, N.M.; Zuberdi, A.M.; Alfarra, H.Y. Biotransformation of ethyl p-methoxycinnamate from Kaempferia galanga L. using Aspergillus niger. Int. J. Biosci. 2013, 3, 148–155. [Google Scholar]
- Swislocka, R.; Kwoczuk-Sadowi, M.; Kalinowska, M.; Lewandowski, W. Spectroscopic (FT-IR, FT-Raman, 1H and 13C NMR) and theoretical studies of p-coumaric acid and alkali metal p-coumarates. Spectroscopy 2012, 27, 35–48. [Google Scholar] [CrossRef]
- Hasegawa, T.; Hashimoto, M.; Fujihara, T.; Yamada, H. Aroma profile of galangal composed of cinnamic acid derivatives and their structure-odor relationships. Nat. Prod. Commun. 2016, 11, 1463–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Präbst, K.; Engelhardt, H.; Ringgeler, S.; Hübner, H. Basic colorimetric proliferation assays: MTT, WST, and resazurin. In Cell Viability Assays; Humana Press: New York, NY, USA, 2017; pp. 1–17. [Google Scholar]
- Martincuks, A.; Andryka, K.; Küster, A.; Schmitz-Van de Leur, H.; Komorowski, M.; Müller-Newen, G. Nuclear translocation of STAT3 and NF-κB are independent of each other but NF-κB supports expression and activation of STAT3. Cell. Signal. 2017, 32, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, D.; Ghosh, S. Regulation of the NF-κB-mediated transcription of inflammatory genes. Front. Immunol. 2014, 5, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Zhang, J.; Arfuso, F.; Chinnathambi, A.; Zayed, M.; Alharbi, S.A.; Kumar, A.P.; Ahn, K.S.; Sethi, G. NF-κB in cancer therapy. Arch. Toxicol. 2015, 89, 711–731. [Google Scholar] [CrossRef] [PubMed]
- Rauert-Wunderlich, H.; Siegmund, D.; Maier, E.; Giner, T.; Bargou, R.C.; Wajant, H.; Stühmer, T. The IKK inhibitor Bay 11-7082 induces cell death independent from inhibition of activation of NFκB transcription factors. PLoS ONE 2013, 8, e59292. [Google Scholar] [CrossRef]
- Quest Graph™ IC50 Calculator. AAT Bioquest, Inc. Available online: https://www.aatbio.com/tools/ic50-calculator (accessed on 9 December 2021).
- Dolcet, X.; Llobet, D.; Pallares, J.; Matias-Guiu, X. NF-κB in development and progression of human cancer. Virchows Arch. 2005, 446, 475–482. [Google Scholar] [CrossRef]
- Guy, J.B.; Espenel, S.; Vallard, A.; Batiston-Montagne, P.; Wozny, A.S.; Ardail, D.; Alphonse, G.; Rancoule, C.; Rodriguez-Lafrasse, C.; Magne, N. Evaluation of the cell invasion and migration process: A comparison of the video microscope-based scratch wound assay and the boyden chamber assay. J. Viz. Exp. 2017, 129, e56337. [Google Scholar] [CrossRef]
- Ma, H.; Yokoyama, S.; Saiki, I.; Hayakawa, Y. Chemosensitizing effect of saikosaponin B on B16F10 melanoma cells. Nutr. Cancer 2017, 69, 505–511. [Google Scholar] [CrossRef]
- An, J.; Huang, Y.-C.; Xu, Q.-Z.; Zhou, L.-J.; Shang, Z.-F.; Huang, B.; Wang, Y.; Liu, X.-D.; Wu, D.-C.; Zhou, P.-K. DNA-PKcs plays a dominant role in the regulation of H2AX phosphorylation in response to DNA damage and cell cycle progression. BMC Mol. Biol. 2010, 11, 18. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.-F.; Kong, F.-L. Akt regulates RSK2 to alter phosphorylation level of H2A. X in breast cancer. Oncol. Lett. 2021, 21, 187. [Google Scholar] [CrossRef]
- Brown, K.K.; Montaser-Kouhsari, L.; Beck, A.H.; Toker, A. MERIT40 Is an Akt substrate that promotes resolution of DNA damage induced by chemotherapy. Cell Rep. 2015, 11, 1358–1366. [Google Scholar] [CrossRef] [Green Version]
- Habibie, H.; Yokoyama, S.; Abdelhamed, S.; Awale, S.; Sakurai, H.; Hayakawa, Y.; Saiki, I. Survivin suppression through STAT3/β-catenin is essential for resveratrol-induced melanoma apoptosis. Int. J. Oncol. 2014, 45, 895–901. [Google Scholar] [CrossRef] [Green Version]
- Winingsih, W.; Husein, S.G.; Ramdhani, R.P.N. Analysis of ethyl p-methoxycinnamate from Kaempferia galanga L. extract by high performance liquid chromatography. J. Trop. Pharm. Chem. 2021, 5, 353–358. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lallo, S.; Hardianti, B.; Sartini, S.; Ismail, I.; Laela, D.; Hayakawa, Y. Ethyl P-Methoxycinnamate: An Active Anti-Metastasis Agent and Chemosensitizer Targeting NFκB from Kaempferia galanga for Melanoma Cells. Life 2022, 12, 337. https://doi.org/10.3390/life12030337
Lallo S, Hardianti B, Sartini S, Ismail I, Laela D, Hayakawa Y. Ethyl P-Methoxycinnamate: An Active Anti-Metastasis Agent and Chemosensitizer Targeting NFκB from Kaempferia galanga for Melanoma Cells. Life. 2022; 12(3):337. https://doi.org/10.3390/life12030337
Chicago/Turabian StyleLallo, Subehan, Besse Hardianti, Sartini Sartini, Ismail Ismail, Dewi Laela, and Yoshihiro Hayakawa. 2022. "Ethyl P-Methoxycinnamate: An Active Anti-Metastasis Agent and Chemosensitizer Targeting NFκB from Kaempferia galanga for Melanoma Cells" Life 12, no. 3: 337. https://doi.org/10.3390/life12030337