
Predator-Prey Interactions between Nonnative Juvenile Largemouth Bass (Micropterus salmoides) and Local Candidate Prey Species in the Pearl River Delta: Predation Capacity, Preference and Growth Performance

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Maintenance of Predator and Prey
2.2. Predation Capacity Experiment
2.3. Predation Preference Experiment
2.4. Growth Performance Experiment
2.5. Statistical Analysis
3. Results
3.1. Predation Capacity
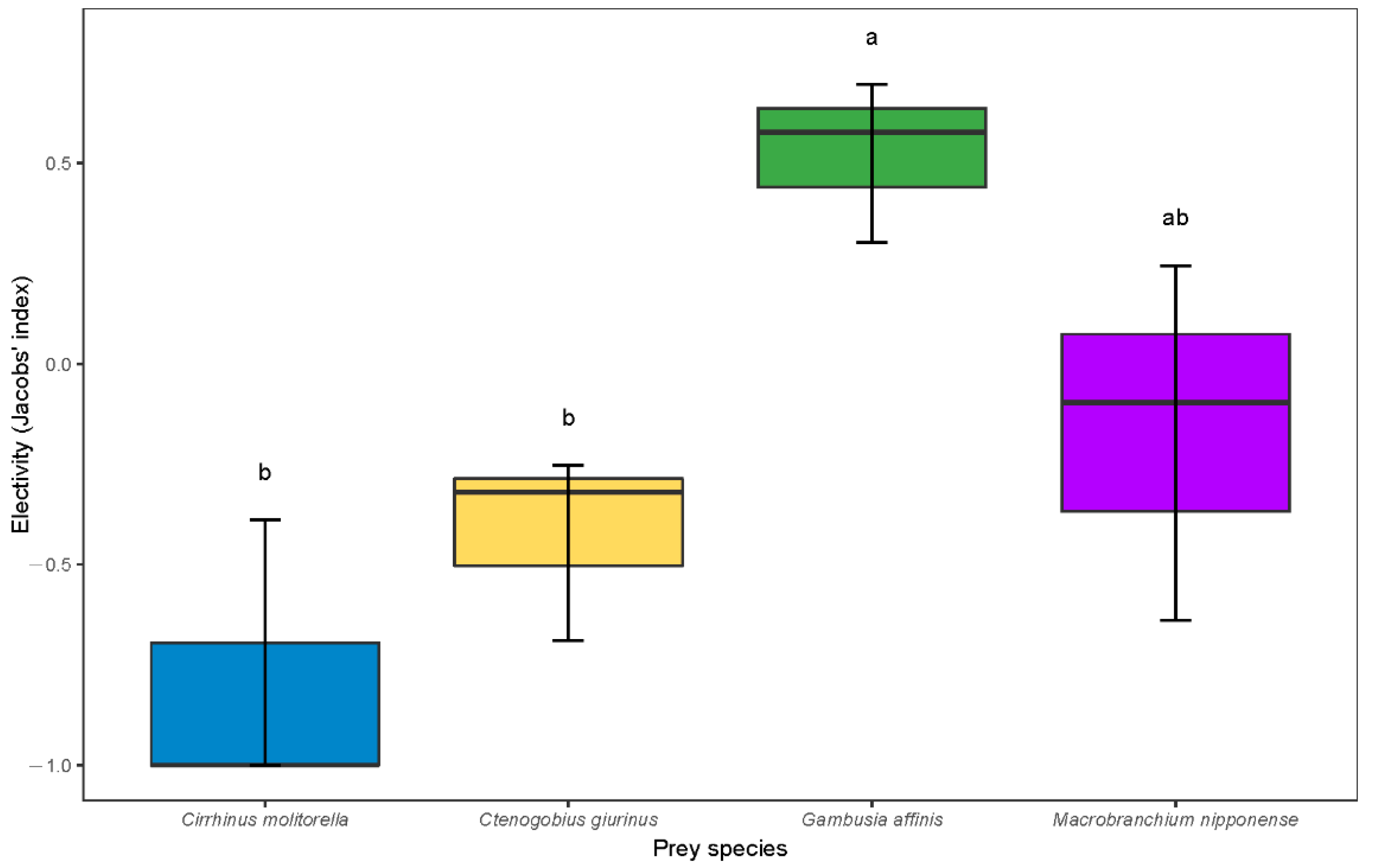
3.2. Predation Preference
3.3. Growth Performance
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nunn, A.D.; Tewson, L.H.; Cowx, I.G. The foraging ecology of larval and juvenile fishes. Rev. Fish Biol. Fish. 2012, 22, 377–408. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Persson, L. The ontogeny of piscivory and its ecological consequences. Can. J. Fish. Aquat. Sci. 1998, 55, 1454–1465. [Google Scholar] [CrossRef]
- Armstrong, J.B.; Schindler, D.E. Excess digestive capacity in predators reflects a life of feast and famine. Nature 2011, 476, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Mihalitsis, M.; Hemingson, C.R.; Goatley, C.H.R.; Bellwood, D.R. The role of fishes as food: A functional perspective on predator–prey interactions. Funct. Ecol. 2021, 35, 1109–1119. [Google Scholar] [CrossRef]
- Battini, N.; Giachetti, C.B.; Castro, K.L.; Bortolus, A.; Schwindt, E. Predator–prey interactions as key drivers for the invasion success of a potentially neurotoxic sea slug. Biol. Invasions 2021, 23, 1207–1229. [Google Scholar] [CrossRef]
- Hall, E.S.; Martin, B.E.; Brubaker, K.; Grant, C.J. Latitudinal variation in the geometric morphology of the largemouth bass, Micropterus salmoides. Mar. Freshw. Res. 2018, 69, 1480–1485. [Google Scholar] [CrossRef]
- Olson, M.H. Ontogenetic Niche Shifts in Largemouth Bass: Variability and Consequences for First-Year Growth. Ecology 1996, 77, 179–190. [Google Scholar] [CrossRef]
- Wang, D.; Yao, H.; Li, Y.H.; Xu, Y.J.; Ma, X.F.; Wang, H.P. Global diversity and genetic landscape of natural populations and hatchery stocks of largemouth bass micropterus salmoides across American and Asian regions. Sci. Rep. 2019, 9, 16697. [Google Scholar] [CrossRef]
- Hussein, G.H.G.; Chen, M.; Qi, P.P.; Cui, Q.K.; Yu, Y.; Hu, W.H.; Tian, Y.; Fan, Q.X.; Gao, Z.X.; Feng, M.W.; et al. Aquaculture industry development, annual price analysis and out-of-season spawning in largemouth bass Micropterus salmoides. Aquaculture 2020, 519, 734901. [Google Scholar] [CrossRef]
- Junjie, B.; Shengjie, L. Chapter 1—Germplasm Resources of Largemouth Bass. In Genetic Breeding and Molecular Marker-Assisted Selective Breeding of Largemouth Bass; Junjie, B., Shengjie, L., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 1–41. [Google Scholar]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species. A Selection from the Global Invasive Species Database (IUCN); The Invasive Species Specialist Group (ISSG): Auckland, New Zealand, 2000; 12p. [Google Scholar]
- Takamura, K. Performance as a fish predator of largemouth bass [Micropterus salmoides (Lacepède)] invading Japanese freshwaters: A review. Ecol. Res. 2007, 22, 940–946. [Google Scholar] [CrossRef]
- Kirjasniemi, M.; Valtonen, T. Size-dependent over-winter mortality of young-of-the-year roach, Rutilus rutilus. Environ. Biol. Fishes 1997, 50, 451–456. [Google Scholar] [CrossRef]
- Hoxmeier, R.J.H.; Wahl, D.H.; Brooks, R.C.; Heidinger, R.C. Growth and survival of age-0 walleye (Sander vitreus): Interactions among walleye size, prey availability, predation, and abiotic factors. Can. J. Fish. Aquat. Sci. 2006, 63, 2173–2182. [Google Scholar] [CrossRef]
- Wheeler, A.P.; Allen, M.S. Habitat and Diet Partitioning between Shoal Bass and Largemouth Bass in the Chipola River, Florida. Trans. Am. Fish. Soc. 2003, 132, 438–449. [Google Scholar] [CrossRef]
- Diana, M.; Wahl, D. Growth and Survival of Four Sizes of Stocked Largemouth Bass. N. Am. J. Fish. Manag. 2009, 29, 1653–1663. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. Editors. FishBase. World Wide Web Electronic Publication. Version (08/2021) 2021. Available online: https://www.fishbase.org (accessed on 11 June 2021).
- Jacobs, J. Quantitative measurement of food selection: A modification of the forage ratio and Ivlev’s electivity index. Oecologia 1974, 14, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Love, J.W.; Newhard, J.J. Using Published Information to Predict Consumption by Northern Snakehead in Maryland. Trans. Am. Fish. Soc. 2021, 150, 425–434. [Google Scholar] [CrossRef]
- Shao, X.N.; Lu, Q.; Xiong, M.Y.; Bu, H.L.; Shi, X.Y.; Wang, D.J.; Zhao, J.D.; Li, S.; Yao, M. Prey partitioning and livestock consumption in the world’s richest large carnivore assemblage. Curr. Biol. 2021, 31, 4887–4897.e4885. [Google Scholar] [CrossRef]
- Crane, D.P.; Ogle, D.H.; Shoup, D.E. Use and misuse of a common growth metric: Guidance for appropriately calculating and reporting specific growth rate. Rev. Aquac. 2020, 12, 1542–1547. [Google Scholar] [CrossRef]
- Hopkins, K.D. Reporting fish growth: A review of the basics. J. World Aquac. Soc. 1992, 23, 173–179. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Pettorelli, N.; Hilborn, A.; Duncan, C.; Durant, S.M. Chapter Two—Individual Variability: The Missing Component to Our Understanding of Predator–Prey Interactions. In Advances in Ecological Research; Pawar, S., Woodward, G., Dell, A.I., Eds.; Academic Press: Cambridge, MA, USA, 2015; Volume 52, pp. 19–44. [Google Scholar]
- Post, D.M. Individual variation in the timing of ontogenetic niche shifts in largemouth bass. Ecology 2003, 84, 1298–1310. [Google Scholar] [CrossRef]
- Richard, B.A.; Wainwright, P.C. Scaling the feeding mechanism of largemouth bass (Micropterus salmoides): Kinematics of prey capture. J. Exp. Biol. 1995, 198, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Brose, U.; Ehnes, R.B.; Rall, B.C.; Vucic-Pestic, O.; Berlow, E.L.; Scheu, S. Foraging theory predicts predator–prey energy fluxes. J. Anim. Ecol. 2008, 77, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Sanna, D.; Azzena, I.; Scarpa, F.; Cossu, P.; Pira, A.; Gagliardi, F.; Casu, M. First Record of the Alien Species Procambarus virginalis Lyko, 2017 in Fresh Waters of Sardinia and Insight into Its Genetic Variability. Life 2021, 11, 606. [Google Scholar] [CrossRef] [PubMed]
- Freedman, J.A.; Hoxmeier, R.J.H.; Einfalt, L.M.; Brooks, R.C.; Wahl, D.H. Largemouth Bass Predation Effect on Stocked Walleye Survival in Illinois Impoundments. N. Am. J. Fish. Manag. 2012, 32, 1039–1045. [Google Scholar] [CrossRef]
- Michaelson, D.P.; Ney, J.J.; Sutton, T.M. Largemouth Bass Predation on Stocked Striped Bass in Smith Mountain Lake, Virginia. N. Am. J. Fish. Manag. 2001, 21, 326–332. [Google Scholar] [CrossRef]
- Godinho, F.N.; Ferreira, M.T.; Cortes, R.V. The environmental basis of diet variation in pumpkinseed sunfish, Lepomis gibbosus, and largemouth bass, Micropterus salmoides, along an Iberian river basin. Environ. Biol. Fishes 1997, 50, 105–115. [Google Scholar] [CrossRef]
- Goclowski, M.R.; Kaeser, A.J.; Sammons, S.M. Movement and Habitat Differentiation among Adult Shoal Bass, Largemouth Bass, and Spotted Bass in the Upper Flint River, Georgia. N. Am. J. Fish. Manag. 2013, 33, 56–70. [Google Scholar] [CrossRef]
- Laegdsgaard, P.; Johnson, C. Why do juvenile fish utilise mangrove habitats? J. Exp. Mar. Biol. Ecol. 2001, 257, 229–253. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.A.; Anderson, R.L.; Wang, J.; Gillespie, N.; Lampo, E.G.; McClelland, N.N.; Solomon, L.E.; Pendleton, R.; Lamer, J.T. Juvenile Silver Carp and Bighead Carp as Forage for Predatory Fish in the LaGrange Reach of the Illinois River. N. Am. J. Fish. Manag. 2021. [Google Scholar] [CrossRef]
- Sanft, E.; Parkos, J.J., III; Collins, S.F.; Porreca, A.P.; Wahl, D.H. Vulnerability of Juvenile Bighead and Silver Carps to Predation by Largemouth Bass. Trans. Am. Fish. Soc. 2018, 147, 1207–1214. [Google Scholar] [CrossRef]
- Weber, M.J.; Dettmers, J.M.; Wahl, D.H.; Czesny, S.J. Effects of Predator–Prey Interactions and Benthic Habitat Complexity on Selectivity of a Foraging Generalist. Trans. Am. Fish. Soc. 2010, 139, 1004–1013. [Google Scholar] [CrossRef]
- Belgrad, B.A.; Griffen, B.D. Predator-prey interactions mediated by prey personality and predator hunting mode. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, L.C.; Hutton, S.A.; Harman, L.; Russell Poole, W.; Gargan, P.; McGinnity, P.; Reed, T.E. Associations between metabolic traits and growth rate in brown trout (Salmo trutta) depend on thermal regime. Proc. R. Soc. B Biol. Sci. 2021, 288, 20211509. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Xiong, M.; Yu, J.; Li, W.; Li, B.; Liu, J.; Zhang, T. Effects of artificial submersed vegetation on consumption and growth of mandarin fish Siniperca chuatsi (Basilewsky) foraging on live prey. J. Freshw. Ecol. 2019, 34, 433–444. [Google Scholar] [CrossRef]
- Malinovskyi, O.; Rahimnejad, S.; Stejskal, V.; Boňko, D.; Stará, A.; Velíšek, J.; Policar, T. Effects of different photoperiods on growth performance and health status of largemouth bass (Micropterus salmoides) juveniles. Aquaculture 2022, 548, 737631. [Google Scholar] [CrossRef]
- Wang, L.; Cui, Z.H.; Ren, X.; Li, P.; Wang, Y. Growth performance, feed cost and environmental impact of largemouth bass Micropterus salmoides fed low fish meal diets. Aquac. Rep. 2021, 20, 100757. [Google Scholar] [CrossRef]
- Min, Z.Z.; Mei, M.D.; Jie, B.J.; Jia, F.J.; Li, L.G.; Fang, L.X. Effects of formulated diets and frozen trash fish on growth and expression of LPL gene mRNA in largemouth bass Micropterus salmoides. J. Dalian Ocean Univ. 2014, 29, 360–363. [Google Scholar]


{kind=link}
| No. | Species | Common Name | Family | Origin | MXL (cm) | Habitat and Food |
|---|---|---|---|---|---|---|
| 1 | Macrobranchium nipponense | Oriental river prawn | Palaemonidae | China | 10.0 | freshwater, brackish; phytoplankton, zoobenthos, detritus |
| 2 | Ctenogobius giurinus | Barcheek goby | Gobionellinae | China | 12.1 | marine, freshwater, brackish; zoobenthos, plants, zooplankton |
| 3 | Gambusia affinis | Western Mosquitofish | Poeciliinae | North and Central America | 7.0 | freshwater, brackish; zooplankton, zoobenthos, others |
| 4 | Hypophthalmichthys molitrix | Silver carp | Cyprinidae | China | 120.0 | freshwater, brackish; plants, zooplankton |
| 5 | Cirrhinus molitorella | Mud carp | Cyprinidae | China | 55.0 | freshwater; plants, detritus, zoobenthos |
| No. | Prey Species | TL (cm) | BM (g/ind.) | APN (ind.) | PCN (ind.) | RPCN (%) | PCW (g) |
|---|---|---|---|---|---|---|---|
| 1 | Macrobranchium nipponense | 3.6 ± 0.8 | 0.40 ± 0.07 | 20 | 6.0 ± 3.0 bc | 55.6 ± 19.2 b | 2.5 ± 1.0 c |
| 2 | Ctenogobius giurinus | 3.7 ± 1.4 | 1.32 ± 0.15 | 20 | 12.0 ± 1.4 b | 83.2 ± 2.0 a | 12.6 ± 1.6 a |
| 3 | Gambusia affinis | 3.4 ± 1.2 | 0.29 ± 0.01 | 50 | 22.5 ± 3.5 a | 89.5 ± 7.8 a | 8.0 ± 0.1 ab |
| 4 | Hypophthalmichthys molitrix | 4.6 ± 0.4 | 1.70 ± 0.30 | 15 | 4.0 ± 0.0 c | 100.0 ± 0.0 a | 5.0 ± 0.3 bc |
| 5 | Cirrhinus molitorella | 3.9 ± 0.3 | 1.06 ± 0.09 | 20 | 8.0 ± 1.7 bc | 100.0 ± 0.0 a | 7.9 ± 2.3 ab |
| No. | Prey Species | TL (cm) | BM (g) | SGR | SGR2 | ADG (g/d) | FCR |
|---|---|---|---|---|---|---|---|
| 1 | Macrobranchium nipponense | 3.6 ± 0.5 | 0.40 ± 0.03 | 2.78 ± 0.41 a | 2.74 ± 0.40 a | 1.46 ± 0.48 a | 4.33 ± 0.71 a |
| 2 | Gambusia affinis | 3.4 ± 0.4 | 0.22 ± 0.02 | 2.64 ± 0.70 a | 2.61 ± 0.37 a | 1.64 ± 0.63 a | 3.37 ± 0.65 a |
| 3 | Hypophthalmichthys molitrix | 4.0 ± 0.2 | 1.24 ± 0.40 | 3.35 ± 0.44 a | 3.30 ± 0.43 a | 1.16 ± 0.32 a | 3.48 ± 1.11 a |
| 4 | Cirrhinus molitorella | 4.8 ± 0.4 | 1.20 ± 0.25 | 3.06 ± 0.65 a | 3.02 ± 0.63 a | 2.08 ± 0.57 a | 3.71 ± 0.15 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, D.; Ye, M.; Yang, D. Predator-Prey Interactions between Nonnative Juvenile Largemouth Bass (Micropterus salmoides) and Local Candidate Prey Species in the Pearl River Delta: Predation Capacity, Preference and Growth Performance. Life 2022, 12, 295. https://doi.org/10.3390/life12020295
Luo D, Ye M, Yang D. Predator-Prey Interactions between Nonnative Juvenile Largemouth Bass (Micropterus salmoides) and Local Candidate Prey Species in the Pearl River Delta: Predation Capacity, Preference and Growth Performance. Life. 2022; 12(2):295. https://doi.org/10.3390/life12020295
Chicago/Turabian StyleLuo, Du, Minghao Ye, and Dingtian Yang. 2022. "Predator-Prey Interactions between Nonnative Juvenile Largemouth Bass (Micropterus salmoides) and Local Candidate Prey Species in the Pearl River Delta: Predation Capacity, Preference and Growth Performance" Life 12, no. 2: 295. https://doi.org/10.3390/life12020295