
A “Mammalian-like” Pycnodont Fish: Independent Acquisition of Thecodont Implantation, True Vertical Replacement, and Carnassial Dentitions in Carnivorous Mammals and a Peculiar Group of Pycnodont Fish


{kind=link}
{kind=link}
Abstract
:1. Introduction
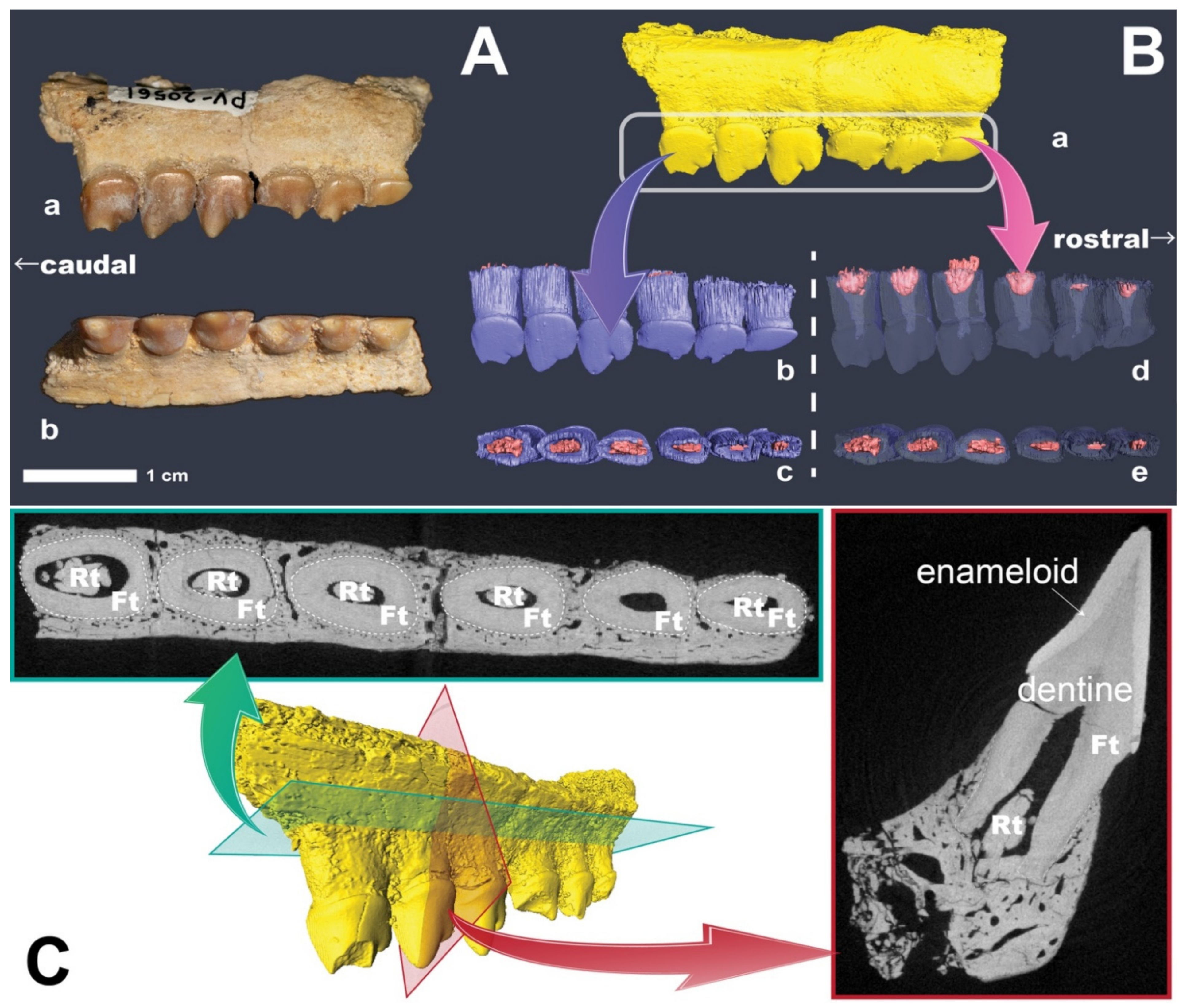
2. Methods
Computed Tomography
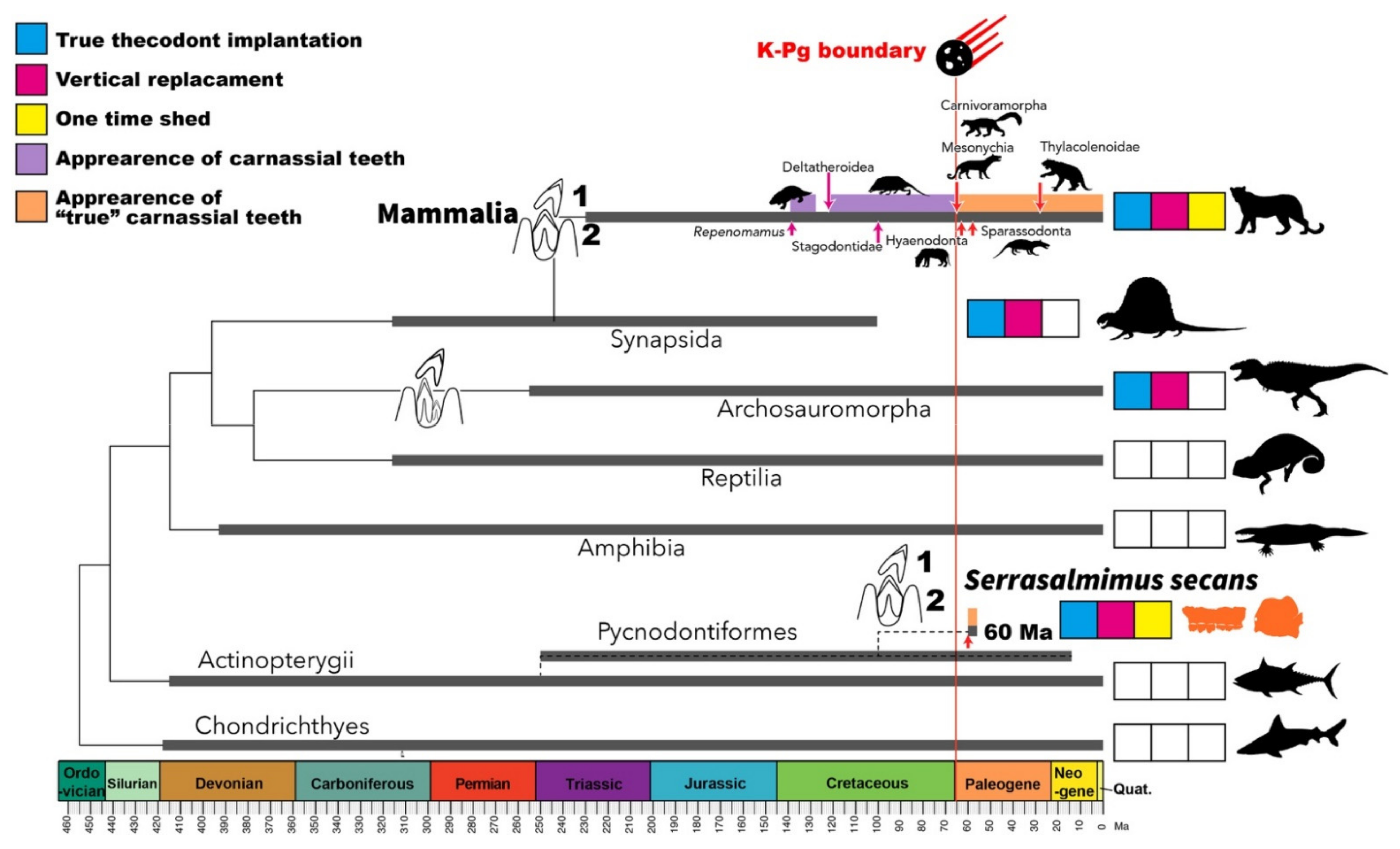
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NMNS | National Museum of Natural History, Ibaraki, Japan |
References
- Chen, D.; Blom, H.; Sanchez, S.; Tafforeau, P.; Ahlberg, P.E. The stem osteichthyan Andreolepis and the origin of tooth replacement. Nature 2016, 539, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Bertin, T.J.C.; Thivichon-Prince, B.; LeBlanc, A.R.H.; Caldwell, M.W.; Viriot, L. Current perspectives on tooth implantation, attachment, and replacement in amniota. Front. Physiol. 2018, 9, 1630. [Google Scholar] [CrossRef] [PubMed]
- Whitlock, J.A.; Mantilla, J.A.W. The late jurassic sauropod dinosaur ‘Morosaurus’ agilis Marsh, 1889 reexamined and reinterpreted as a dicraeosaurid. J. Vertebr. Paleontol. 2020, 40, e1780600. [Google Scholar] [CrossRef]
- Salomies, L.; Eymann, J.; Khan, I.; Di-Poï, N. The alternative regenerative strategy of bearded dragon unveils the key processes underlying vertebrate tooth renewal. Elife 2019, 8, e47702. [Google Scholar] [CrossRef]
- Cooper, S.L.A.; Martill, D.M. A diverse assemblage of pycnodont fishes (Actinopterygii, Pycnodontiformes) from the mid-Cretaceous, continental kem kem group of south-east Morocco. Cretac. Res. 2020, 112, 104456. [Google Scholar] [CrossRef]
- Vullo, R.; Cavin, L.; Khalloufi, B.; Amaghzaz, M.; Bardet, N.; Jalil, N.-E.; Jourani, E.; Khaldoune, F.; Gheerbrant, E. A unique Cretaceous–Paleogene lineage of piranha-jawed pycnodont fishes. Sci. Rep. 2017, 7, 6802. [Google Scholar] [CrossRef]
- Marcus, M.A.; Amini, S.; Stifler, C.A.; Sun, C.-Y.; Tamura, N.; Bechtel, H.A.; Parkinson, D.Y.; Barnard, H.S.; Zhang, X.X.X.; Chua, J.Q.I.; et al. Parrotfish teeth: Stiff biominerals whose microstructure makes them tough and abrasion-resistant to bite stony corals. ACS Nano 2017, 11, 11856–11865. [Google Scholar] [CrossRef] [Green Version]
- Bemis, K.E.; Burke, S.M.; St. John, C.A.; Hilton, E.J.; Bemis, W.E. Tooth development and replacement in the Atlantic cutlassfish, Trichiurus lepturus, with comparisons to other scombroidei. J. Morphol. 2019, 280, 78–94. [Google Scholar] [CrossRef] [Green Version]
- Bemis, W.E.; Giuliano, A.; McGuire, B. Structure, attachment, replacement and growth of teeth in bluefish, Pomatomus saltatrix ((Linnaeus, 1766), a teleost with deeply socketed teeth. Zoology 2005, 108, 317–327. [Google Scholar] [CrossRef]
- Sattler, F.; Schwarz, D. Tooth replacement in a specimen of Tyrannosaurus rex (Dinosauria, Theropoda) from the Hell Creek Formation (Maastrichtian), Montana. Hist. Biol. 2021, 33, 949–972. [Google Scholar] [CrossRef]
- Butler, P.M. The evolution of Carnassial dentitions in the Mammalia. Proc. Zool. Soc. Lond. 1946, 116, 198–220. [Google Scholar] [CrossRef]
- Muizon, C.; Lange-Badré, B. Carnivorous dental adaptations in tribosphenic mammals and phylogenetic reconstruction. Lethaia 2007, 30, 353–366. [Google Scholar] [CrossRef]
- Van Valkenburgh, B. Déjà vu: The evolution of feeding morphologies in the Carnivora. Integr. Comp. Biol. 2007, 47, 147–163. [Google Scholar] [CrossRef] [Green Version]
- Lang, A.J.; Engler, T.; Martin, T. Dental topographic and three-dimensional geometric morphometric analysis of carnassialization in different clades of carnivorous mammals (Dasyuromorphia, Carnivora, Hyaenodonta). J. Morphol. 2021, 283, 91–108. [Google Scholar] [CrossRef]
- Hu, Y.; Meng, J.; Wang, Y.; Li, C. Large Mesozoic mammals fed on young dinosaurs. Nature 2005, 433, 149–152. [Google Scholar] [CrossRef]
- Lyson, T.R.; Miller, I.M.; Bercovici, A.D.; Weissenburger, K.; Fuentes, A.J.; Clyde, W.C.; Hagadorn, J.W.; Butrim, M.J.; Johnson, K.R.; Fleming, R.F.; et al. Exceptional continental record of biotic recovery after the Cretaceous−Paleogene mass extinction. Science 2019, 366, 977–983. [Google Scholar] [CrossRef]
- Jablonski, D. Extinctions in the fossil record. Philos. Trans. R. Soc. B Biol. Sci. 1994, 344, 11–17. [Google Scholar] [CrossRef]
- Schulte, P.; Alegret, L.; Arenillas, I.; Arz, J.A.; Barton, P.J.; Bown, P.R.; Bralower, T.J.; Christeson, G.L.; Claeys, P.; Cockell, C.S.; et al. The chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary. Science 2010, 327, 1214–1218. [Google Scholar] [CrossRef] [Green Version]
- Bourque, R.D.; Douglas, P.M.J.; Larsson, H.C.E. Changes in terrestrial ecosystems across the Cretaceous-Paleogene boundary in western Canada inferred from plant wax lipid distributions and isotopic measurements. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 562, 110081. [Google Scholar] [CrossRef]
- Brusatte, S.L.; Butler, R.J.; Barrett, P.M.; Carrano, M.T.; Evans, D.C.; Lloyd, G.T.; Mannion, P.D.; Norell, M.A.; Peppe, D.J.; Upchurch, P.; et al. The extinction of the dinosaurs. Biol. Rev. 2015, 90, 628–642. [Google Scholar] [CrossRef] [Green Version]
- Sibert, E.C.; Norris, R.D. New age of fishes initiated by the Cretaceous−Paleogene mass extinction. Proc. Natl. Acad. Sci. USA 2015, 112, 8537–8542. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Tanabe, K. Cretaceous plesiosaurs ate ammonites. Nature 1998, 394, 629–630. [Google Scholar] [CrossRef]
- Schulp, A.S.; Vonhof, H.B.; Van Der Lubbe, J.H.J.L.; Janssen, R.; Van Baal, R.R. On diving and diet: Resource partitioning in type-maastrichtian mosasaurs. Neth. J. Geosci. 2013, 92, 165–170. [Google Scholar] [CrossRef]
- Everhart, M.J.; Hageman, S.A.; Hoffman, B.L. Another sternberg “Fish-within-a-Fish” discovery: First report of Ichthyodectes ctenodon (Teleostei; Ichthyodectiformes) with stomach contents. Trans. Kans. Acad. Sci. 2010, 113, 197–205. [Google Scholar] [CrossRef]
- Buffetaut, E.; Koeberl, C. (Eds.) Geological and Biological Effects of Impact Events; Impact Studies; Springer: Berlin/Heidelberg, Germany, 2002; ISBN 978-3-642-63960-9. [Google Scholar]
- Nursall, J.R. Distribution and ecology of pycnodont fishes. In Mesozoic Fishes—Systematics and Paleoecology; Arratia, G., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 1996; pp. 115–124. ISBN 3-923871–90-2. [Google Scholar]
- Poyato-Ariza, F.J. Pycnodont fishes: Morphologic variation, ecomorphologic plasticity, and a new interpretation of their evolutionary history. Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist. Ser. A 2005, 3, 169–184. [Google Scholar]
- Bajpai, S.; Gingerich, P.D. A new Eocene archaeocete (Mammalia, Cetacea) from India and the time of origin of whales. Proc. Natl. Acad. Sci. USA 1998, 95, 15464–15468. [Google Scholar] [CrossRef] [Green Version]
- Sire, J.Y.; Donoghue, P.C.J.; Vickaryous, M.K. Origin and evolution of the integumentary skeleton in non-tetrapod vertebrates. J. Anat. 2009, 214, 409–440. [Google Scholar] [CrossRef]
- Sasagawa, I.; Ishiyama, M.; Yokosuka, H.; Mikami, M.; Uchida, T. Tooth enamel and enameloid in actinopterygian fish. Front. Mater. Sci. China 2009, 3, 174–182. [Google Scholar] [CrossRef]
- Lacruz, R.S.; Habelitz, S.; Wright, J.T.; Paine, M.L. Dental enamel formation and implications for oral health and disease. Physiol. Rev. 2017, 97, 939–993. [Google Scholar] [CrossRef]
- Kolmann, M.A.; Cohen, K.E.; Bemis, K.E.; Summers, A.P.; Irish, F.J.; Hernandez, L.P. Tooth and consequences: Heterodonty and dental replacement in piranhas and pacus (Serrasalmidae). Evol. Dev. 2019, 21, 278–293. [Google Scholar] [CrossRef]
- Rücklin, M.; King, B.; Cunningham, J.A.; Johanson, Z.; Marone, F.; Donoghue, P.C.J. Acanthodian dental development and the origin of gnathostome dentitions. Nat. Ecol. Evol. 2021, 5, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.; Tibbetts, I.R.; Kemp, A.; Truss, R.; Drennan, J. Inferring parrotfish (Teleostei: Scaridae) pharyngeal mill function from dental morphology, wear, and microstructure. J. Morphol. 2006, 267, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Sire, J.-Y.; Davit-Beal, T.; Delgado, S.; Van Der Heyden, C.; Huysseune, A. First-generation teeth in non-mammaliannonmammalian lineages: Evidence for a conserved ancestral character? Microsc. Res. Tech. 2002, 59, 408–434. [Google Scholar] [CrossRef]
- Miller, R.F.; Cloutier, R.; Turner, S. The oldest articulated chondrichthyan from the early Devonian period. Nature 2003, 425, 501–504. [Google Scholar] [CrossRef]
- Lu, J.; Giles, S.; Friedman, M.; den Blaauwen, J.L.; Zhu, M. The oldest actinopterygian highlights the cryptic early history of the hyperdiverse ray-finned fishes. Curr. Biol. 2016, 26, 1602–1608. [Google Scholar] [CrossRef] [Green Version]
- Niedwiedzki, G.; Szrek, P.; Narkiewicz, K.; Narkiewicz, M.; Ahlberg, P.E. Tetrapod trackways from the early middle Devonian period of Poland. Nature 2010, 463, 43–48. [Google Scholar] [CrossRef]
- Falcon-Lang, H.J.; Benton, M.J.; Stimson, M. Ecology of earliest reptiles inferred from basal Pennsylvanian trackways. J. Geol. Soc. 2007, 164, 1113–1118. [Google Scholar] [CrossRef] [Green Version]
- Sennikov, A.G.; Golubev, V.K. Vyazniki biotic assemblage of the terminal Permian. Paleontol. J. 2006, 40, S475–S481. [Google Scholar] [CrossRef]
- Mann, A.; Gee, B.M.; Pardo, J.D.; Marjanović, D.; Adams, G.R.; Calthorpe, A.S.; Maddin, H.C.; Anderson, J.S. Reassessment of historic ‘microsaurs’ from Joggins, Nova Scotia, reveals hidden diversity in the earliest amniote ecosystem. Pap. Palaeontol. 2020, 6, 605–625. [Google Scholar] [CrossRef]
- Bhat, M.S.; Ray, S.; Datta, P.M. New cynodonts (Therapsida, Eucynodontia) from the late Triassic of India and their significances. J. Paleontol. 2021, 95, 376–393. [Google Scholar] [CrossRef]
- Bogdanov, D. Dimetrodon/Dimetrodon milleri. Available online: http://phylopic.org/image/1f3e74df-30e6-422b-9ce2-db7df199f11d/ (accessed on 24 January 2022).
- Karala, M. Tyrannosaurus rex. Available online: http://phylopic.org/image/eb78548d-4f61-43c7-a087-a69074bc8bea/ (accessed on 24 January 2022).
- Zica, M. Repenomamus. Available online: http://phylopic.org/image/ad9a4ed1-2292-4fef-80c7-4f1bc5cb724a/ (accessed on 24 January 2022).
- Zimices Mesonychia/Mesonychidae/Mesonyx/Mesonyx obtusidens. Available online: http://phylopic.org/image/73624419-f6f1-41a7-becf-7f1c90d5b203/ (accessed on 24 January 2022).
- Zimices Cladosictis/Cladostictis patagonica/Hathliacynidae/Sparassodonta. Available online: http://phylopic.org/image/327edeee-e34c-4573-be88-66ad6b14ffc4/ (accessed on 24 January 2022).
- Bell, M.A.; Lloyd, G.T. Strap: Stratigraphic Tree Analysis for Palaeontology. 2014. Available online: https://cran.r-project.org/web/packages/strap/index.html (accessed on 24 January 2022).
- R Core Team. R: A Language and Environment for Statistical Computing. 2021. Available online: https://www.r-project.org/ (accessed on 24 January 2022).
- Huxley, T.H. On the Application of the Laws of Evolution to the Arrangement of the Vertebrata and, more particularly, of the Mammalia. In Proceedings of the Zoological Society of London; Academic Press: London, UK, 1880; pp. 649–662. [Google Scholar]
- Cope, E.D. Geology and paleontology. Am. Nat. 1887, 21, 1014–1019. [Google Scholar]
- Berg, L.S. A classification of fish-like vertebrates. Bulletin de l’Académie Des Sciences de l’URSS 1937, 4, 1277–1280. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsui, K.; Kimura, Y. A “Mammalian-like” Pycnodont Fish: Independent Acquisition of Thecodont Implantation, True Vertical Replacement, and Carnassial Dentitions in Carnivorous Mammals and a Peculiar Group of Pycnodont Fish. Life 2022, 12, 250. https://doi.org/10.3390/life12020250
Matsui K, Kimura Y. A “Mammalian-like” Pycnodont Fish: Independent Acquisition of Thecodont Implantation, True Vertical Replacement, and Carnassial Dentitions in Carnivorous Mammals and a Peculiar Group of Pycnodont Fish. Life. 2022; 12(2):250. https://doi.org/10.3390/life12020250
Chicago/Turabian StyleMatsui, Kumiko, and Yuri Kimura. 2022. "A “Mammalian-like” Pycnodont Fish: Independent Acquisition of Thecodont Implantation, True Vertical Replacement, and Carnassial Dentitions in Carnivorous Mammals and a Peculiar Group of Pycnodont Fish" Life 12, no. 2: 250. https://doi.org/10.3390/life12020250