
Disentangling the Taxonomic Status of Caprella penantis sensu stricto (Amphipoda: Caprellidae) Using an Integrative Approach


Abstract
:1. Introduction
2. Materials and Methods
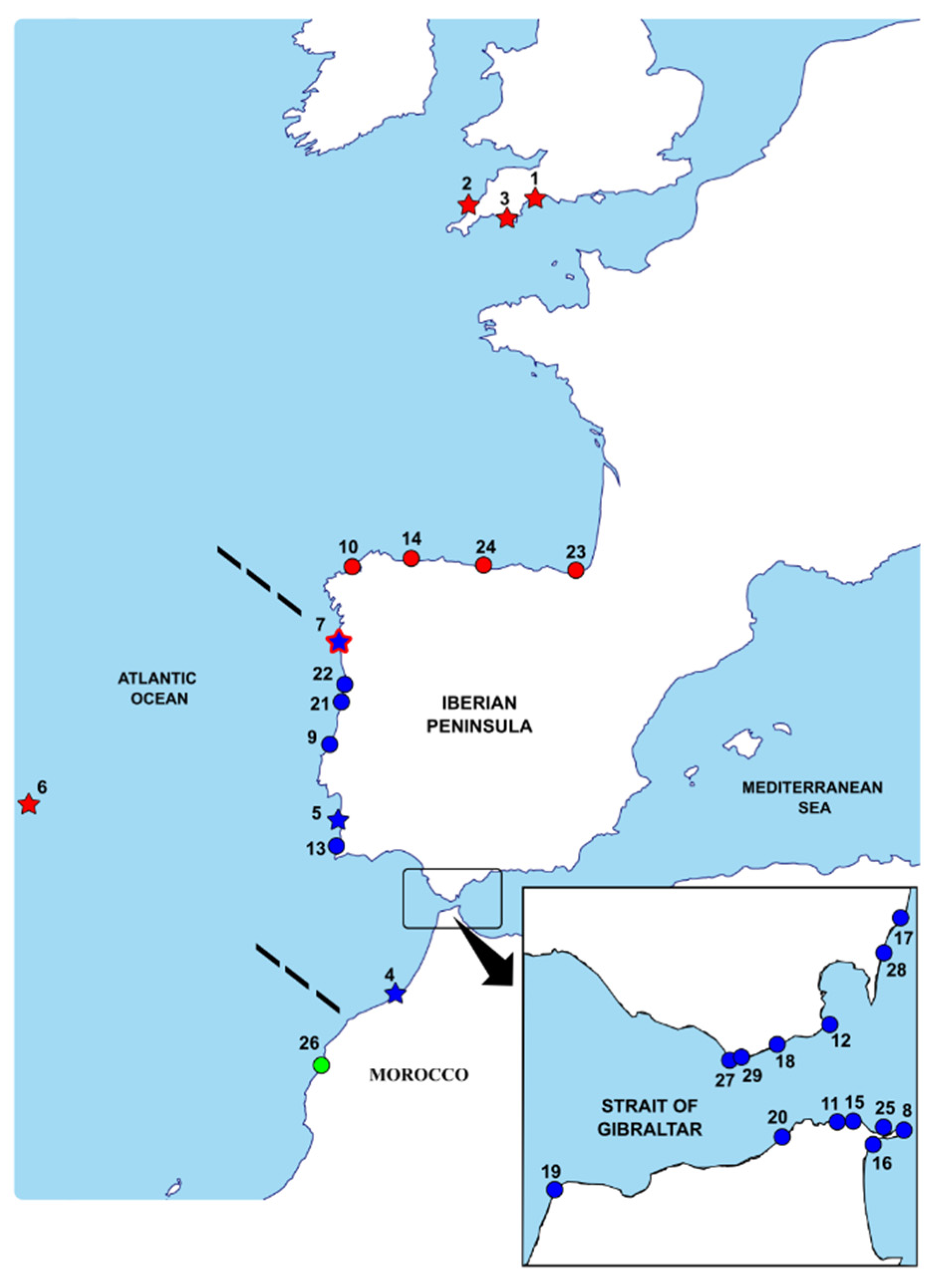
2.1. Sample Collection and Taxonomic Identification
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Sequence Analysis
2.4. Phylogenetic Reconstruction
2.5. Molecular Species Delimitation
2.6. Genetic Structure Analyses
3. Results
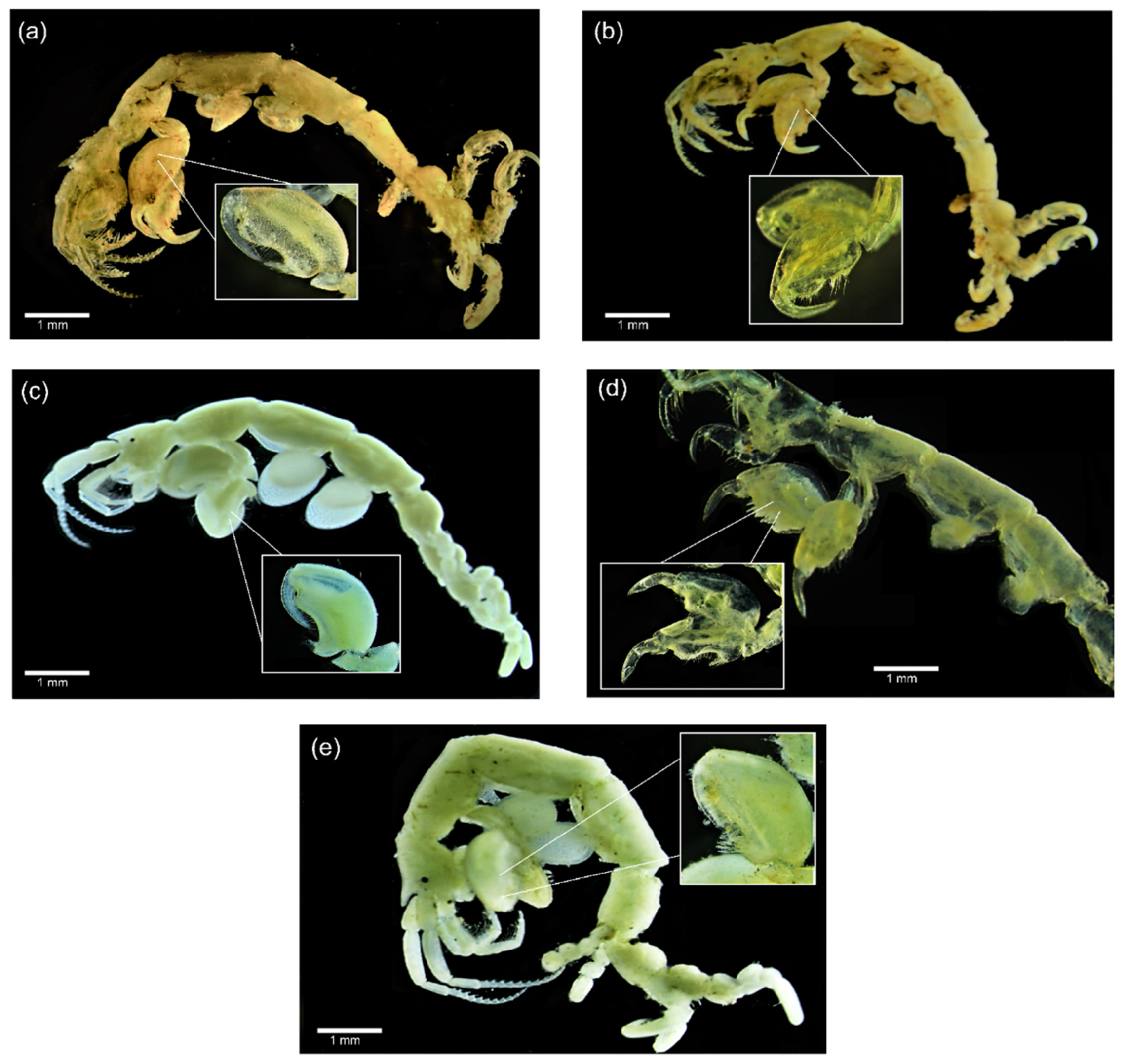
3.1. Morphological Analysis
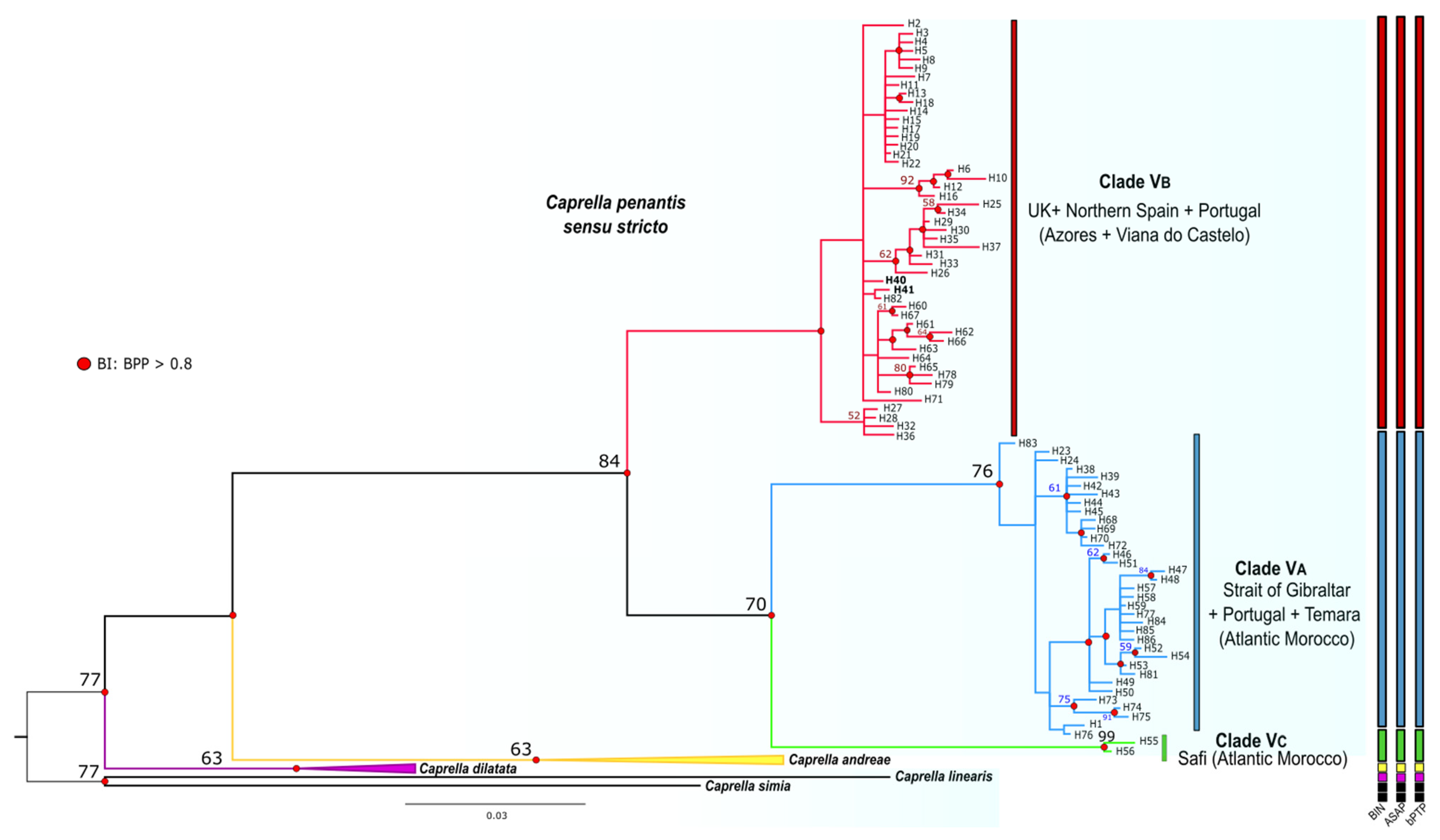
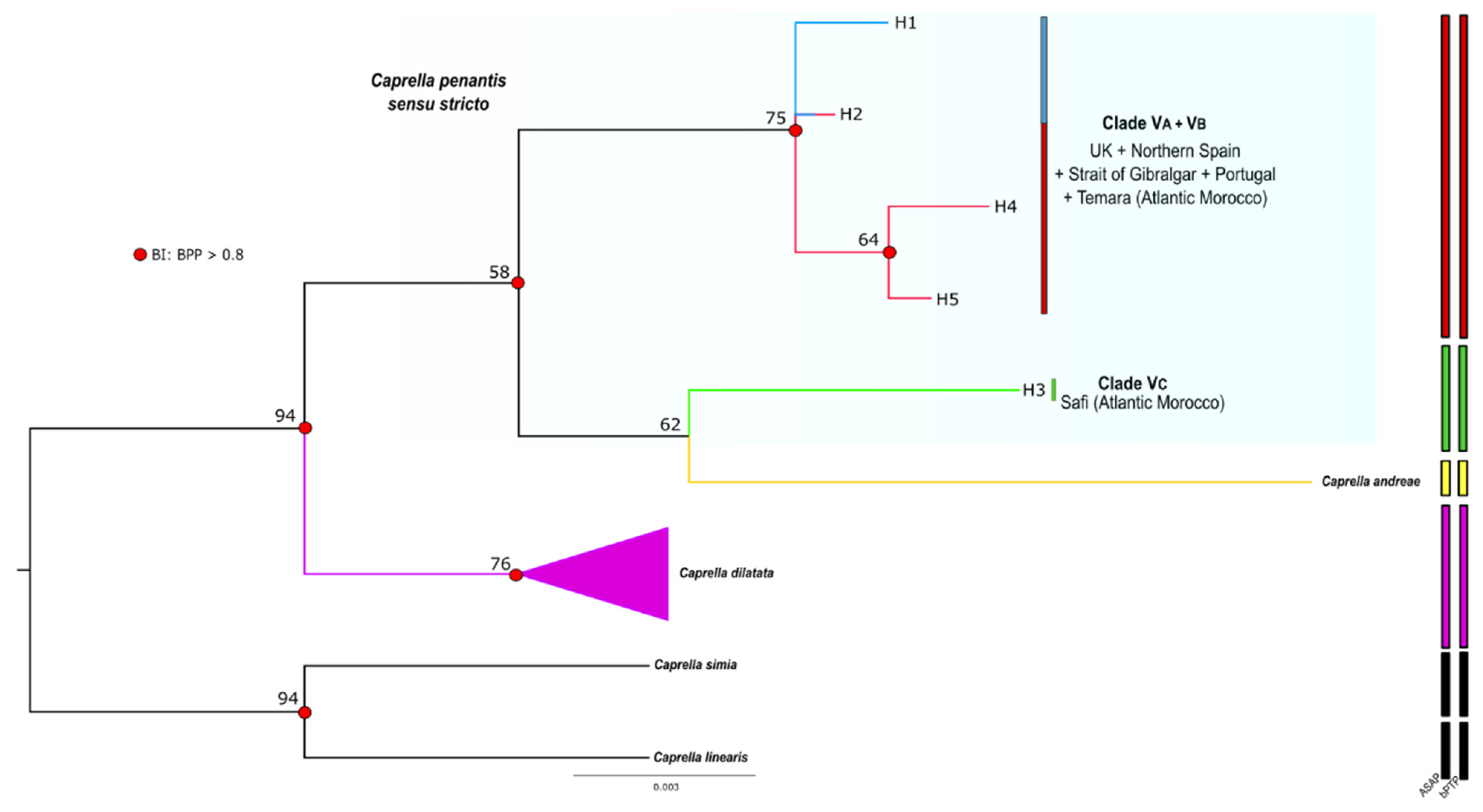
3.2. Phylogenetic Inference and Species Delimitation
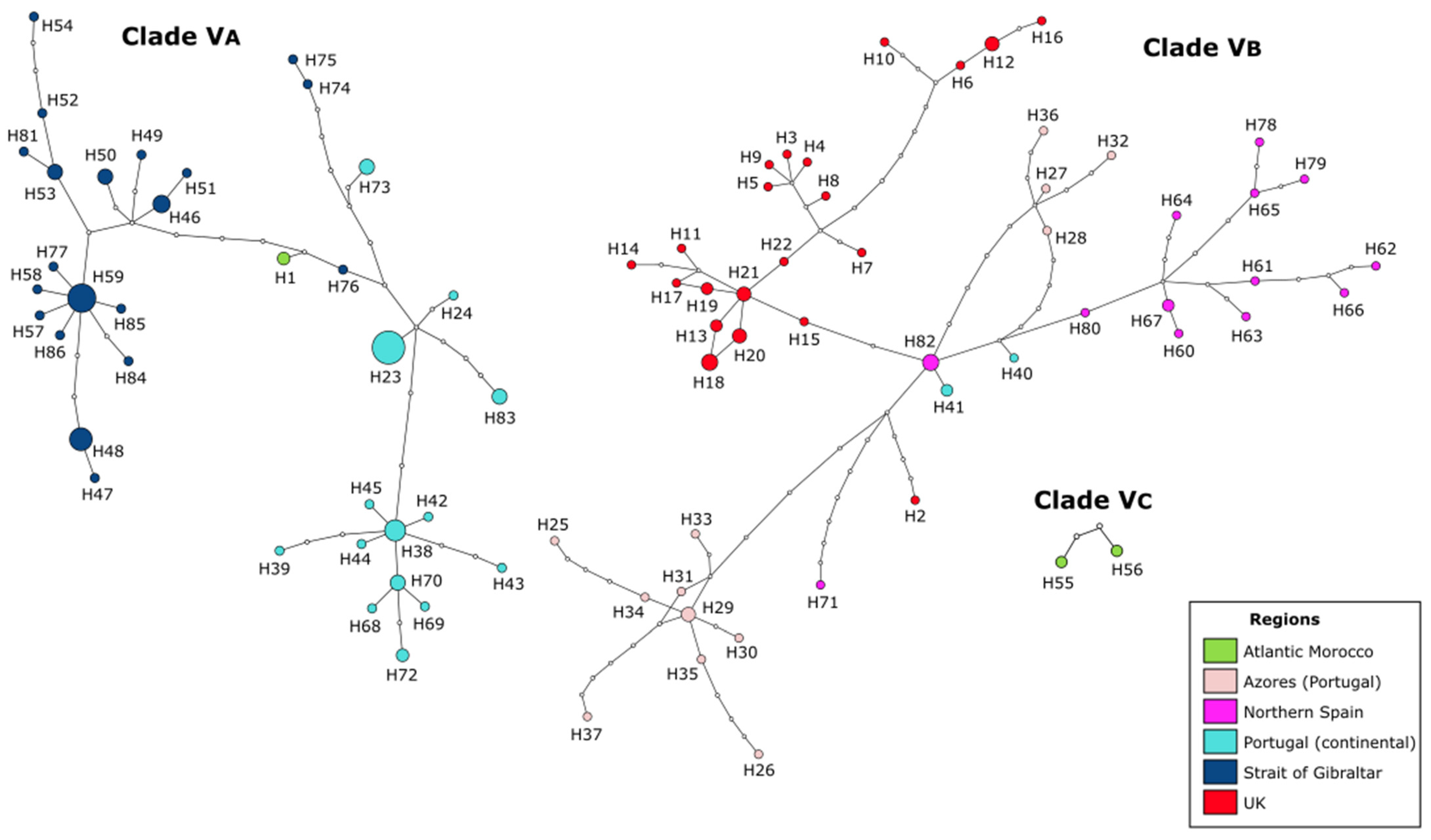
3.3. Phylogeographic Structure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guerra-García, J.M. Habitat use of the Caprellidea (Crustacea: Amphipoda) from Ceuta, North Africa. Ophelia 2001, 55, 27–38. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Baeza-Rojano, E.; Cabezas, M.P.; García-Gómez, J.C. Vertical distribution and seasonality of peracarid crustaceans associated with intertidal macroalgae. J. Sea Res. 2011, 65, 256–264. [Google Scholar] [CrossRef]
- McCain, J.C. The Capreillidae (Crustacea: Amphipoda) of the Western North Atlantic. Bull. US Natl. Mus. 1968, 278, 1–147. [Google Scholar] [CrossRef]
- Vader, W.; Tandberg, A.H.S. Gammarid amphipods (Crustacea) in Norway, with a key to the species. Fauna Nor. 2019, 39, 12–25. [Google Scholar] [CrossRef]
- Jacobucci, G.B.; Tanaka, M.O.; Leite, F.P.P. Temporal variation of amphipod assemblages associated with Sargassum filipendula (Phaeophyta) and its epiphytes in a subtropical shore. Aquat. Ecol. 2009, 43, 1031–1040. [Google Scholar] [CrossRef]
- Thiel, M.; Guerra-García, J.M.; Lancellotti, D.A.; Vásquez, N. The distribution of littoral caprellids (Crustacea: Amphipoda: Caprellidea) along the Pacific Coast of continental Chile. Rev. Chil. Hist. Nat. 2003, 76, 297–312. [Google Scholar] [CrossRef] [Green Version]
- Ros, M.; Guerra-García, J.M.; González-Macías, M.; Saavedra, Á.; López-Fe, C.M. Influence of fouling communities on the establishment success of alien caprellids (Crustacea: Amphipoda) in Southern Spain. Mar. Biol. Res. 2013, 9, 261–273. [Google Scholar] [CrossRef]
- Martínez-Laiz, G.; Guerra-García, J.M.; Ros, M.; Fenwick, D.; Bishop, J.D.; Horton, T.; Faasse, M.A.; Cabezas, M.P. Hitchhiking northwards: On the presence of the invasive skeleton shrimp Caprella scaura in the UK. Mar. Biodivers. 2021, 51, 78. [Google Scholar] [CrossRef]
- Ros, M.; Tierno de Figueroa, J.M.; Guerra-García, J.M.; Navarro-Barranco, C.; Lacerda, M.B.; Vázquez-Luis, M.; Masunari, S. Exploring trophic strategies of exotic caprellids (Crustacea: Amphipoda): Comparison between habitat types and native vs. introduced distribution ranges. Estuar. Coast. Shelf Sci. 2014, 139, 88–98. [Google Scholar] [CrossRef]
- Woods, C.M.C. Caprellid amphipods: An overlooked marine finfish aquaculture resource? Aquaculture 2009, 289, 199–211. [Google Scholar] [CrossRef]
- Elias-Piera, F.; Rossi, S.; Petti, M.A.V.; Campos, L.S.; Valério-Berardo, M.T.; Corbisier, T.N. Fauna Associated with Morphologically Distinct Macroalgae from Admiralty Bay, King George Island (Antarctica). Polar Biol. 2020, 43, 1535–1547. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Cabezas, M.P.; Baeza-Rojano, E.; Espinosa, F.; García-Gómez, J.C. Is the north side of the Strait of Gibraltar more diverse than the south side? A case study using the intertidal peracarids (Crustacea: Malacostraca) associated to the seaweed Corallina elongata. J. Mar. Biol. Assoc. UK 2009, 89, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Barranco, C.; Ros, M.; Tierno de Figueroa, J.M.; Guerra-García, J.M. Marine crustaceans as bioindicators: Amphipods as case study. In Fisheries and Aquaculture; Oxford University Press: New York, NY, USA, 2020; ISBN 9780190865627. [Google Scholar]
- Lourenço, R.A.; Magalhães, C.A.; Taniguchi, S.; Siqueira, S.G.L.; Jacobucci, G.B.; Leite, F.P.P.; Bícego, M.C. Evaluation of macroalgae and amphipods as bioindicators of petroleum hydrocarbons input into the marine environment. Mar. Pollut. Bull. 2019, 145, 564–568. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, I.; Takahashi, S.; Tanabe, S.; Miyazaki, N. Butyltin concentrations along the Japanese coast from 1997 to 1999 monitored by Caprella Spp. (Crustacea: Amphipoda). Mar. Environ. Res. 2004, 57, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Guerra-GarcÍa, J.M.; Koonjul, M.S. Metaprotella sandalensis (Crustacea: Amphipoda: Caprellidae): A bioindicator of nutrient enrichment on coral reefs? Environ. Monit. Assess. 2005, 104, 353. [Google Scholar] [CrossRef] [PubMed]
- Guerra-García, J.M.; Baeza-Rojano, E.; Cabezas, M.P.; Díaz-Pavón, J.J.; Pacios, I.; García-Gómez, J.C. The amphipods Caprella penantis and Hyale schmidtii as Biomonitors of trace metal contamination in intertidal ecosystems of Algeciras Bay, southern Spain. Mar. Pollut. Bull. 2009, 58, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Guerra-García, J.M.; Hachero-Cruzado, I.; González-Romero, P.; Jiménez-Prada, P.; Cassell, C.; Ros, M. Towards integrated multi-trophic aquaculture: Lessons from caprellids (Crustacea: Amphipoda). PLoS ONE 2016, 11, e0154776. [Google Scholar] [CrossRef] [Green Version]
- Baeza-Rojano, E.; Hachero-Cruzado, I.; Guerra-García, J.M. Nutritional analysis of freshwater and marine amphipods from the Strait of Gibraltar and potential aquaculture applications. J. Sea Res. 2014, 85, 29–36. [Google Scholar] [CrossRef]
- Baeza-Rojano, E.; Domingues, P.; Guerra-García, J.M.; Capella, S.; Noreña-Barroso, E.; Caamal-Monsreal, C.; Rosas, C. Marine gammarids (Crustacea: Amphipoda): A new live prey to culture Octopus maya hatchlings. Aquac. Res. 2013, 44, 1602–1612. [Google Scholar] [CrossRef]
- Baeza-Rojano, E.; Calero-Cano, S.; Hachero-Cruzado, I.; Guerra-García, J.M. A preliminary study of the Caprella scaura amphipod culture for potential use in aquaculture. J. Sea Res. 2013, 83, 146–151. [Google Scholar] [CrossRef]
- Suontama, J.; Kiessling, A.; Melle, W.; Waagbø, R.; Olsen, R.E. Protein from Northern krill (Thysanoessa inermis), Antarctic krill (Euphausia superba) and the Arctic amphipod (Themisto libellula) can partially replace fish meal in diets to Atlantic salmon (Salmo salar) without affecting product quality. Aquac. Nutr. 2007, 13, 50–58. [Google Scholar] [CrossRef]
- Hughes, L.E.; Ahyong, S.T. Collecting and processing amphipods. J. Crustac. Biol. 2016, 36, 584–588. [Google Scholar] [CrossRef] [Green Version]
- Arfianti, T.; Wilson, S.; Costello, M.J. Progress in the discovery of amphipod crustaceans. PeerJ 2018, 6, e5187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobo, J.; Ferreira, M.S.; Antunes, I.C.; Teixeira, M.A.L.; Borges, L.M.S.; Sousa, R.; Gomes, P.A.; Costa, M.H.; Cunha, M.R.; Costa, F.O. Contrasting morphological and DNA barcode-suggested species boundaries among shallow-water amphipod fauna from the southern European Atlantic coast. Genome 2017, 60, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Coleman, C.O. Taxonomy in times of the taxonomic impediment—Examples from the community of experts on amphipod crustaceans. J. Crustac. Biol. 2015, 35, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Beheregaray, L.B.; Caccone, A. Cryptic biodiversity in a changing world. J. Biol. 2007, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Pilgrim, E.M.; Darling, J.A. Biodiversity Research: Genetic diversity in two introduced biofouling amphipods (Ampithoe valida & Jassa marmorata) along the Pacific North American coast: Investigation into molecular identification and cryptic diversity. Divers. Distrib. 2010, 16, 827–839. [Google Scholar] [CrossRef]
- Cabezas, M.P.; Cabezas, P.; Machordom, A.; Guerra-García, J.M. Hidden diversity and cryptic speciation refute cosmopolitan distribution in Caprella penantis (Crustacea: Amphipoda: Caprellidae). J. Zool. Syst. Evol. Res. 2013, 51, 85–99. [Google Scholar] [CrossRef]
- Cabezas, M.P.; Navarro-Barranco, C.; Ros, M.; Guerra-García, J.M. Long-Distance Dispersal, Low connectivity and molecular evidence of a new cryptic species in the obligate rafter Caprella andreae Mayer, 1890 (Crustacea: Amphipoda: Caprellidae). Helgol. Mar. Res. 2013, 67, 483–497. [Google Scholar] [CrossRef]
- Costa, F.O.; Cunha, M.R.; Neuparth, T.; Theodorakis, C.W.; Costa, M.H.; Shugart, L.R. Application of RAPD DNA fingerprinting in taxonomic identification of amphipods: A case-study with Gammarus species (Crustacea: Amphipoda). J. Mar. Biol. Assoc. UK 2004, 84, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Ros, M.; Guerra-García, J.M.; Navarro-Barranco, C.; Cabezas, M.P.; Vázquez-Luis, M. The spreading of the non-native caprellid (Crustacea: Amphipoda) Caprella scaura Templeton, 1836 into southern Europe and northern Africa: A complicated taxonomic history. Mediterr. Mar. Sci. 2014, 15, 145–155. [Google Scholar] [CrossRef]
- Martínez-Laiz, G.; Ros, M.; Guerra-García, J.M.; Marchini, A.; Fernández-González, V.; Vazquez-Luis, M.; Lionello, M.; Scribano, G.; Sconfietti, R.; Ferrario, J.; et al. Scientific collaboration for early detection of invaders results in a significant update on estimated range: Lessons from Stenothoe georgiana Bynum & Fox 1977. Mediterr. Mar. Sci. 2020, 21, 464–481. [Google Scholar] [CrossRef]
- Marchini, A.; Cardeccia, A. Alien amphipods in a sea of troubles: Cryptogenic species, unresolved taxonomy and overlooked introductions. Mar. Biol. 2017, 164, 69. [Google Scholar] [CrossRef]
- Guerra-García, J.M.; Espinosa, F.; García-Gómez, J.C. Trends in taxonomy today: An overview about the main topics in taxonomy. Zool. Baetica 2008, 19, 15–49. [Google Scholar]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identification through DNA barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheth, B.P.; Thaker, V.S. DNA barcoding and traditional taxonomy: An integrated approach for biodiversity conservation. Genome 2017, 60, 618–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayrat, B. Towards integrative taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–415. [Google Scholar] [CrossRef]
- Radulovici, A.E.; Sainte-Marie, B.; Dufresne, F. DNA barcoding of marine crustaceans from the estuary and Gulf of St Lawrence: A regional-scale approach. Mol. Ecol. Resour. 2009, 9, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.O.; Henzler, C.M.; Lunt, D.H.; Whiteley, N.M.; Rock, J. Probing marine Gammarus (Amphipoda) taxonomy with DNA barcodes. Syst. Biodivers. 2009, 7, 365–379. [Google Scholar] [CrossRef]
- Jażdżewska, A.M.; Mamos, T. High species richness of Northwest Pacific deep-sea amphipods revealed through DNA barcoding. Prog. Oceanogr. 2019, 178, 102184. [Google Scholar] [CrossRef]
- McCain, J.C.; Steinberg, J.E. Amphipoda-I, Caprellidea-I, Family Caprellidae. In Crustaceorum Catalogus 2; Junk: Berlin, Germany, 1970; pp. 1–78. [Google Scholar]
- Laubitz, D.R. The Caprellidae (Crustacea, Amphipoda) of Atlantic and Arctic Canada; National Museums of Canada: Ottawa, ON, Canada, 1972. [Google Scholar]
- Mayer, P. Die Caprelliden Des Golfes von Neapel Und Der Angrenzenden Meeres—Abschnitte; R. Friedländer & Sohn: Berlin, Germany, 1890; pp. 1–198.
- Mayer, P. Die Caprellidae der Siboga-Expedition; Siboga Expeditie; Buchhandlung und Druckerei Vormals E. J. Brill: Leiden, The Netherlands, 1903. [Google Scholar]
- Utinomi, H. The Fauna of Akkeshi Bay XIII. Caprellidae. J. Fac. Sci. Hokkaido Univ. 1943, 8, 283–300. [Google Scholar]
- Dougherty, E.C.; Steinberg, J. Notes on the skeleton shrimps (Crustacea: Caprellidae) of California. Proc. Biol. Soc. Wash. 1954, 66, 39–49. [Google Scholar]
- Vassilenko, S. Fauna of Caprellidae (Amphipoda) of the Possjet Bay (the Sea of Japan) and some data on their ecology. Explor. Fauna Seas 1967, 58, 196–229. [Google Scholar]
- McCain, J.C. The Capreillidae (Crustacea: Amphipoda) of the Western North Atlantic; Smithsonian Institution Press: Washington, DC, USA, 1968. [Google Scholar]
- Krapp-Schickel, T. Suborder Caprellidea. In The Amphipoda of the Mediterranean: Part 3; Ruffo, S., Ed.; Mémoires de l’Institute Océanographique: Monaco City, Monaco, 1993; Volume 13, pp. 577–813. [Google Scholar]
- Guerra-García, J.M. Identification Guide to British Caprellidae v.3.3; NMBAQC 2012 taxonomic workshop; Dove Marine Laboratory: North Shields, UK, 2014. [Google Scholar]
- Geller, J.; Meyer, C.; Parker, M.; Hawk, H. Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application in all-taxa biotic surveys. Mol. Ecol. Resour. 2013, 13, 851–861. [Google Scholar] [CrossRef]
- Cabezas, M.P.; Ros, M.; Santos, A.M.; Martínez-Laiz, G.; Xavier, R.; Montelli, L.; Hoffman, R.; Fersi, A.; Dauvin, J.C.; Guerra-García, J.M. Unravelling the origin and introduction pattern of the tropical species Paracaprella pusilla Mayer, 1890 (Crustacea, Amphipoda, Caprellidae) in temperate European waters: First molecular insights from a spatial and temporal perspective. NeoBiota 2019, 47, 43–80. [Google Scholar] [CrossRef]
- Whiting, M.F. Mecoptera Is paraphyletic: Multiple genes and phylogeny of Mecoptera and Siphonaptera. Zool. Scr. 2002, 31, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strimmer, K.; von Haeseler, A.; Salemi, M. Genetic distances and nucleotide substitution models. In The Phylogenetic Handbook: A Practical Approach to Phylogenetic Analysis and Hypothesis Testing; Vandamme, A.-M., Salemi, M., Lemey, P., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 111–141. ISBN 9780521730716. [Google Scholar]
- Pond, S.L.K.; Poon, A.F.Y.; Frost, S.D.W. Estimating selection pressures on alignments of coding sequences. In The Phylogenetic Handbook: A Practical Approach to Phylogenetic Analysis and Hypothesis Testing; Vandamme, A.-M., Salemi, M., Lemey, P., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 419–490. ISBN 9780-521730716. [Google Scholar]
- Zwickl, D.J. Genetic Algorithm Approaches for the Phylogenetic Analysis of Large Biological Sequence Datasets under the Maximum Likelihood Criterion. Ph.D. Thesis, The University of Texas at Austin, Austin, TX, USA, May 2006. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. A New Look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Sukumaran, J.; Holder, M.T. DendroPy: A Python library for phylogenetic computing. Bioinformatics 2010, 26, 1569–1571. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree-Version 1.4.4, a Graphical Viewer of Phylogenetic Trees. 2018. Available online: https://github.com/rambaut/figtree/releases (accessed on 20 November 2021).
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Templeton, A.R.; Crandall, K.A.; Sing, C.F. A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genetics 1992, 132, 619–633. [Google Scholar] [CrossRef]
- Santos, A.M.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. TcsBU: A Tool to extend TCS network layout and visualization. Bioinformatics 2016, 32, 627–628. [Google Scholar] [CrossRef] [Green Version]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-based registry for all animal species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble Species by Automatic Partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System (Http://Www.Barcodinglife.Org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin Suite Ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of Molecular Variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Bohonak, A.J. IBD (Isolation by Distance): A program for analyses of isolation by distance. J. Hered. 2002, 93, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Horton, T.; Lowry, J.; De Broyer, C.; Bellan-Santini, D.; Coleman, C.O.; Corbari, L.; Costello, M.J.; Daneliya, M.; Dauvin, J.-C.; Fišer, C.; et al. WoRMS—World Register of Marine Species. Caprella Lamarck 1801. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=101430 (accessed on 11 December 2021).
- Guerra-García, J.M.; Tierno de Figueroa, J.M. What do caprellids (Crustacea: Amphipoda) feed on? Mar. Biol. 2009, 156, 1881–1890. [Google Scholar] [CrossRef]
- Cabezas, M.P. Taxonomía Molecular y Biogeografía de los Caprélidos (Crustacea: Amphipoda) del Estrecho de Gibraltar. Ph.D. Thesis, University of Seville, Seville, Spain, October 2013. [Google Scholar]
- Cabezas, M.P.; Xavier, R.; Branco, M.; Santos, A.M.; Guerra-García, J.M. Invasion history of Caprella scaura Templeton, 1836 (Amphipoda: Caprellidae) in the Iberian Peninsula: Multiple introductions revealed by mitochondrial sequence data. Biol. Invasions 2014, 16, 2221–2245. [Google Scholar] [CrossRef]
- Baird, H.P.; Miller, K.J.; Stark, J.S. Evidence of hidden biodiversity, ongoing speciation and diverse patterns of genetic structure in giant Antarctic amphipods. Mol. Ecol. 2011, 20, 3439–3454. [Google Scholar] [CrossRef]
- Witt, J.D.S.; Threloff, D.L.; Hebert, P.D.N. DNA barcoding reveals extraordinary cryptic diversity in an amphipod genus: Implications for desert spring conservation. Mol. Ecol. 2006, 15, 3073–3082. [Google Scholar] [CrossRef]
- Mohrbeck, I.; Horton, T.; Jażdżewska, A.M.; Martínez Arbizu, P. DNA barcoding and cryptic diversity of deep-sea scavenging amphipods in the Clarion-Clipperton zone (Eastern Equatorial Pacific). Mar. Biodivers. 2021, 51, 26. [Google Scholar] [CrossRef]
- Vieira, P.E.; Desiderato, A.; Holdich, D.M.; Soares, P.; Creer, S.; Carvalho, G.R.; Costa, F.O.; Queiroga, H. Deep segregation in the open ocean: Macaronesia as an evolutionary hotspot for low dispersal marine invertebrates. Mol. Ecol. 2019, 28, 1784–1800. [Google Scholar] [CrossRef] [PubMed]
- Barrowclough, G.F.; Zink, R.M. Funds enough, and time: mtDNA, nuDNA and the discovery of divergence. Mol. Ecol. 2009, 18, 2934–2936. [Google Scholar] [CrossRef]
- Maggs, C.A.; Castilho, R.; Foltz, D.; Henzler, C.; Jolly, M.T.; Kelly, J.; Olsen, J.; Perez, K.E.; Stam, W.; Väinölä, R.; et al. Evaluating signatures of glacial refugia for North Atlantic benthic marine taxa. Ecology 2008, 89, S108–S122. [Google Scholar] [CrossRef] [Green Version]
- Palumbi, S.R. Marine speciation on a small planet. Trends Ecol. Evol. 1992, 7, 114–118. [Google Scholar] [CrossRef]
- Patarnello, T.; Volckaert, F.A.M.J.; Castilho, R. Pillars of Hercules: Is the Atlantic-Mediterranean transition a phylogeographical break? Mol. Ecol. 2007, 16, 4426–4444. [Google Scholar] [CrossRef]
- Xavier, R.; Zenboudji, S.; Lima, F.P.; Harris, D.J.; Santos, A.M.; Branco, M. Phylogeography of the marine isopod Stenosoma nadejda (Rezig, 1989) in north African Atlantic and western Mediterranean coasts reveals complex differentiation patterns and a new Species. Biol. J. Linn. Soc. 2011, 104, 419–431. [Google Scholar] [CrossRef] [Green Version]
- Desiderato, A.; Costa, F.O.; Serejo, C.S.; Abbiati, M.; Queiroga, H.; Vieira, P.E. Macaronesian islands as promoters of diversification in amphipods: The remarkable case of the Family Hyalidae (Crustacea, Amphipoda). Zool. Scr. 2019, 48, 359–375. [Google Scholar] [CrossRef]
- Vieira, P.E.; Desiderato, A.; Azevedo, S.L.; Esquete, P.; Costa, F.O.; Queiroga, H. Molecular evidence for sweeping discontinuity between peracarid (Crustacea) fauna of Macaronesian islands and nearby continental coasts: Over fifty candidate endemic species. bioRxiv 2021. [Google Scholar] [CrossRef]
- Virgilio, M.; Fauvelot, C.; Costantini, F.; Abbiati, M.; Backeljau, T. Phylogeography of the common ragworm Hediste diversicolor (Polychaeta: Nereididae) reveals cryptic diversity and multiple colonization events across its distribution. Mol. Ecol. 2009, 18, 1980–1994. [Google Scholar] [CrossRef] [PubMed]
- Delić, T.; Stoch, F.; Borko, Š.; Flot, J.-F.; Fišer, C. How did subterranean amphipods cross the Adriatic Sea? Phylogenetic evidence for dispersal–vicariance interplay mediated by marine regression–transgression cycles. J. Biogeogr. 2020, 47, 1875–1887. [Google Scholar] [CrossRef]
- Gómez-Gesteira, M.; deCastro, M.; Alvarez, I.; Gómez-Gesteira, J.L. Coastal sea surface temperature warming trend along the continental part of the Atlantic Arc (1985–2005). J. Geophys. Res. Ocean. 2008, 113, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Savolainen, O.; Lascoux, M.; Merilä, J. Ecological genomics of local adaptation. Nat. Rev. Genet. 2013, 14, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Gittenberger, E. What about non-adaptive radiation? Biol. J. Linn. Soc. 1991, 43, 263–272. [Google Scholar] [CrossRef]
- Mathews, L.M. Cryptic biodiversity and phylogeographical patterns in a snapping shrimp species complex. Mol. Ecol. 2006, 15, 4049–4063. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Locality | Country | Coordinates | Collection Data | N | H | Haplotype Codes | Phylogenetic Clade (COI) | GenBank acc. nos. COI/18S |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Sidmouth, Devonshire coast | UK | 50°40′29.881″ N, 3°14′44.992″ W | 14/08/2018 | 14 /3 | 6/1 | H17-H22/H2 | VB | OM057907−OM057920/OM112220−OM112222 |
| 2 | St. Ives | UK | 50°13′9.685″ N, 5°28′42.211″ W | 13/08/2018 | 17/2 | 14/1 | H3-H16/H2 | VB | OM057890−OM057906/OM112218−OM112219 |
| 3 | Wembury, Devonshire coast | UK | 50°18′57.287″ N, 4°4′59.124″ W | 20/06/2010 | 1 | 1 | H2 | VB | OM057889 |
| 4 | Temara | Morocco | 33°54′50.904″ N, 6°58′49.577″ W | 31/01/2010 | 2/1 | 1/1 | H1/H1 | VA | OM057887−OM057888/OM112217 |
| 5 | Alteirinhos | Portugal | 37°31′9.89″ N, 8°47′18.949″ W | 05/06/2012 | 12 | 2 | H23-H24 | VA | OM057921−OM057932 |
| 6 | Monte da Guia, Azores | Portugal | 38°31′9.102″ N, 28°37′40.778″ W | 02/12/2014 | 15/5 | 13/2 | H25-H37/H2 | VB | OM057933−OM057947/OM112223−OM112227 |
| 7 | Viana do Castelo | Portugal | 41°41′59.59″ N, 8°51′24.559″ W | 14/07/2014 | 14/13 | 8/1 | H38-H45/H2 | VA, VB | OM057948−OM057961/OM112228−OM112240 |
| Clade VA | Clade VB | Clade VC | C. dilatata | C. andreae | C. simia | C. linearis | |
|---|---|---|---|---|---|---|---|
| Clade VA | -- | -- | -- | -- | -- | -- | |
| Clade VB | 10.2 | 0.4 | 0.9 | 1.1 | 1.6 | 1.3 | |
| Clade VC | 10.3 | 10.6 | 0.8 | 1.0 | 1.5 | 1.2 | |
| C. dilatata | 13.4 | 13.4 | 13.2 | 1.3 | 1.5 | 1.4 | |
| C. andreae | 15.0 | 14.3 | 14.2 | 11.1 | 2.3 | 1.6 | |
| C. simia | 19.2 | 19.4 | 19.6 | 17.5 | 17.0 | 0.9 | |
| C. linearis | 22.6 | 21.3 | 21.1 | 19.8 | 20.9 | 19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabezas, M.P.; Guerra-García, J.M.; Santos, A.M. Disentangling the Taxonomic Status of Caprella penantis sensu stricto (Amphipoda: Caprellidae) Using an Integrative Approach. Life 2022, 12, 155. https://doi.org/10.3390/life12020155
Cabezas MP, Guerra-García JM, Santos AM. Disentangling the Taxonomic Status of Caprella penantis sensu stricto (Amphipoda: Caprellidae) Using an Integrative Approach. Life. 2022; 12(2):155. https://doi.org/10.3390/life12020155
Chicago/Turabian StyleCabezas, M. Pilar, José M. Guerra-García, and António M. Santos. 2022. "Disentangling the Taxonomic Status of Caprella penantis sensu stricto (Amphipoda: Caprellidae) Using an Integrative Approach" Life 12, no. 2: 155. https://doi.org/10.3390/life12020155