
Lipid Handling Protein Gene Expression in Colorectal Cancer: CD36 and Targeting miRNAs
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Collection of Biological Specimens
2.2. Gene Expression Analysis
2.3. miRNA Selection and Expression Analysis
2.4. In Silico Validation of Gene Expression Results
2.5. Immunohistochemistry Analysis
2.6. Statistical Analysis
3. Results
3.1. Gene Expression Levels
3.2. miRNA Levels
3.3. miRNA/mRNA Correlations
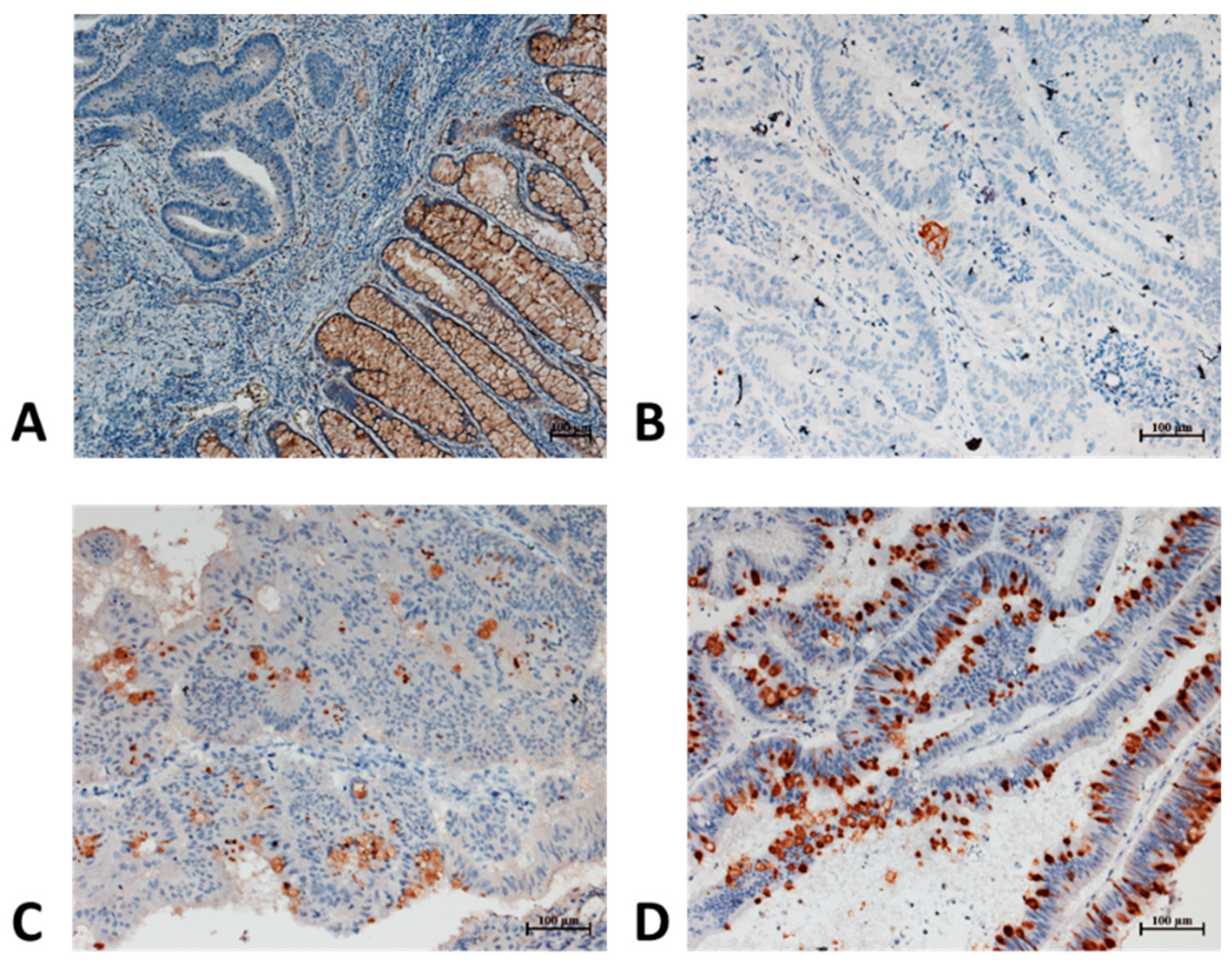
3.4. CD36 Immunostaining
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bradbury, K.E.; Murphy, N.; Key, T.J. Diet and colorectal cancer in UK Biobank: A prospective study. Int. J. Epidemiol. 2020, 49, 246–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.-H.; Wu, K.; Ng, K.; Zauber, A.G.; Nguyen, L.H.; Song, M.; He, X.; Fuchs, C.S.; Ogino, S.; Willett, W.C.; et al. Association of Obesity with Risk of Early-Onset Colorectal Cancer Among Women. JAMA Oncol. 2019, 5, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namasivayam, V.; Lim, S. Recent advances in the link between physical activity, sedentary behavior, physical fitness, and colorectal cancer. F1000Research 2017, 6, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-Y.; Wilkens, L.R.; Setiawan, V.W.; Monroe, K.R.; Haiman, C.A.; Le Marchand, L. Alcohol Intake and Colorectal Cancer Risk in the Multiethnic Cohort Study. Am. J. Epidemiol. 2019, 188, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Amitay, E.L.; Carr, P.R.; Jansen, L.; Roth, W.; Alwers, E.; Herpel, E.; Kloor, M.; Bläker, H.; Chang-Claude, J.; Brenner, H.; et al. Smoking, alcohol consumption and colorectal cancer risk by molecular pathological subtypes and pathways. Br. J. Cancer 2020, 122, 1604–1610. [Google Scholar] [CrossRef] [Green Version]
- Coleman, O.; Ecker, M.; Haller, D. Dysregulated lipid metabolism in colorectal cancer. Curr. Opin. Gastroenterol. 2022, 38, 162–167. [Google Scholar] [CrossRef]
- Fhu, C.W.; Ali, A. Fatty Acid Synthase: An Emerging Target in Cancer. Molecules 2020, 25, 3935. [Google Scholar] [CrossRef]
- Drury, J.; Young, L.E.A.; Scott, T.L.; Kelson, C.O.; He, D.; Liu, J.; Wu, Y.; Wang, C.; Weiss, H.L.; Fan, T.; et al. Tissue-Specific Downregulation of Fatty Acid Synthase Suppresses Intestinal Adenoma Formation via Coordinated Reprograming of Transcriptome and Metabolism in the Mouse Model of Apc-Driven Colorectal Cancer. Int. J. Mol. Sci. 2022, 23, 6510. [Google Scholar] [CrossRef]
- Enciu, A.-M.; Radu, E.; Popescu, I.D.; Hinescu, M.E.; Ceafalan, L.C. Targeting CD36 as Biomarker for Metastasis Prognostic: How Far from Translation into Clinical Practice? Biomed Res. Int. 2018, 2018, 7801202. [Google Scholar] [CrossRef] [Green Version]
- Lobo, M.V.; Huerta, L.; Ruiz-Velasco, N.; Teixeiro, E.; de la Cueva, P.; Celdrán, A.; Martín-Hidalgo, A.; Vega, M.A.; Bragado, R. Localization of the lipid receptors CD36 and CLA-1/SR-BI in the human gastrointestinal tract: Towards the identification of receptors mediating the intestinal absorption of dietary lipids. J. Histochem. Cytochem. 2001, 49, 1253–1260. [Google Scholar] [CrossRef]
- Ladanyi, A.; Mukherjee, A.; Kenny, H.A.; Johnson, A.; Mitra, A.K.; Sundaresan, S.; Nieman, K.M.; Pascual, G.; Benitah, S.A.; Montag, A.; et al. Adipocyte-induced CD36 expression drives ovarian cancer progression and metastasis. Oncogene 2018, 37, 2285–2301. [Google Scholar] [CrossRef] [PubMed]
- Hale, J.S.; Otvos, B.; Sinyuk, M.; Alvarado, A.G.; Hitomi, M.; Stoltz, K.; Wu, Q.; Flavahan, W.; Levison, B.; Johansen, M.L.; et al. Cancer stem cell-specific scavenger receptor CD36 drives glioblastoma progression. Stem Cells 2014, 32, 1746–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Zhi, Z.; Wang, C.; Xing, H.; Song, G.; Yu, X.; Zhu, Y.; Wang, X.; Zhang, X.; Di, Y. Exogenous lipids promote the growth of breast cancer cells via CD36. Oncol. Rep. 2017, 38, 2105–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual, G.; Avgustinova, A.; Mejetta, S.; Martín, M.; Castellanos, A.; Attolini, C.S.-O.; Berenguer, A.; Prats, N.; Toll, A.; Hueto, J.A.; et al. Targeting metastasis-initiating cells through the fatty acid receptor CD36. Nature 2017, 541, 41–45. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Liao, W.-X.; Huang, S.-Z.; Yu, Y.-F.; Wen, J.-Y.; Chen, J.; Lin, D.-G.; Wu, X.-Y.; Jiang, N.; Li, X. Prognostic and immunological role of CD36: A pan-cancer analysis. J. Cancer 2021, 12, 4762–4773. [Google Scholar] [CrossRef]
- Rachidi, S.M.; Qin, T.; Sun, S.; Zheng, W.J.; Li, Z. Molecular profiling of multiple human cancers defines an inflammatory cancer-associated molecular pattern and uncovers KPNA2 as a uniform poor prognostic cancer marker. PLoS ONE 2013, 8, e57911. [Google Scholar] [CrossRef]
- Drury, J.; Rychahou, P.G.; He, D.; Jafari, N.; Wang, C.; Lee, E.Y.; Weiss, H.L.; Evers, B.M.; Zaytseva, Y.Y. Inhibition of Fatty Acid Synthase Upregulates Expression of CD36 to Sustain Proliferation of Colorectal Cancer Cells. Front. Oncol. 2020, 10, 1185. [Google Scholar] [CrossRef]
- Delaunay, S.; Pascual, G.; Feng, B.; Klann, K.; Behm, M.; Hotz-Wagenblatt, A.; Richter, K.; Zaoui, K.; Herpel, E.; Münch, C.; et al. Mitochondrial RNA modifications shape metabolic plasticity in metastasis. Nature 2022, 607, 593–603. [Google Scholar] [CrossRef]
- Ceafalan, L.C.; Niculae, A.M.; Ioghen, O.; Gherghiceanu, M.; Hinescu, M.E. Metastatic potential. In Unraveling the Complexities of Metastasis; Elsevier: Amsterdam, The Netherlands, 2022; pp. 153–173. [Google Scholar]
- Fang, Y.; Shen, Z.-Y.; Zhan, Y.-Z.; Feng, X.-C.; Chen, K.-L.; Li, Y.-S.; Deng, H.-J.; Pan, S.-M.; Wu, D.-H.; Ding, Y. CD36 inhibits β-catenin/c-myc-mediated glycolysis through ubiquitination of GPC4 to repress colorectal tumorigenesis. Nat. Commun. 2019, 10, 3981. [Google Scholar] [CrossRef] [Green Version]
- Pei, Z.; Fraisl, P.; Berger, J.; Jia, Z.; Forss-Petter, S.; Watkins, P.A. Mouse very long-chain Acyl-CoA synthetase 3/fatty acid transport protein 3 catalyzes fatty acid activation but not fatty acid transport in MA-10 cells. J. Biol. Chem. 2004, 279, 54454–54462. [Google Scholar] [CrossRef]
- Sun, P.; Xia, S.; Lal, B.; Shi, X.; Yang, K.S.; Watkins, P.A.; Laterra, J. Lipid metabolism enzyme ACSVL3 supports glioblastoma stem cell maintenance and tumorigenicity. BMC Cancer 2014, 14, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, Z.; Fraisl, P.; Shi, X.; Gabrielson, E.; Forss-Petter, S.; Berger, J.; Watkins, P.A. Very long-chain acyl-CoA synthetase 3: Overexpression and growth dependence in lung cancer. PLoS ONE 2013, 8, e69392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milger, K.; Herrmann, T.; Becker, C.; Gotthardt, D.; Zickwolf, J.; Ehehalt, R.; Watkins, P.A.; Stremmel, W.; Füllekrug, J. Cellular uptake of fatty acids driven by the ER-localized acyl-CoA synthetase FATP4. J. Cell Sci. 2006, 119, 4678–4688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Physiology of the Gastrointestinal Tract; Elsevier: Amsterdam, The Netherlands, 2018; ISBN 9780128099544.
- An, J.; Oh, H.E.; Kim, H.; Lee, J.-H.; Lee, E.S.; Kim, Y.-S.; Choi, J.-W. Significance of Altered Fatty Acid Transporter Expressions in Uterine Cervical Cancer and Its Precursor Lesions. Anticancer Res. 2022, 42, 2131–2137. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Salathé, E.P.; Schmid-Schönbein, G.W. Fluid exchange in skeletal muscle with viscoelastic blood vessels. Am. J. Physiol. 1987, 253, H1548–H1556. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Jung, J.; Jeong, H.; Lee, J.-H.; Oh, H.E.; Lee, E.S.; Choi, J.-W. High Membranous Expression of Fatty Acid Transport Protein 4 Is Associated with Tumorigenesis and Tumor Progression in Clear Cell Renal Cell Carcinoma. Dis. Markers 2019, 2019, 5702026. [Google Scholar] [CrossRef] [Green Version]
- Yen, M.-C.; Chou, S.-K.; Kan, J.-Y.; Kuo, P.-L.; Hou, M.-F.; Hsu, Y.-L. Solute Carrier Family 27 Member 4 (SLC27A4) Enhances Cell Growth, Migration, and Invasion in Breast Cancer Cells. Int. J. Mol. Sci. 2018, 19, 434. [Google Scholar] [CrossRef] [Green Version]
- To, K.K.; Tong, C.W.; Wu, M.; Cho, W.C. MicroRNAs in the prognosis and therapy of colorectal cancer: From bench to bedside. World J. Gastroenterol. 2018, 24, 2949–2973. [Google Scholar] [CrossRef]
- Zhang, N.; Hu, X.; Du, Y.; Du, J. The role of miRNAs in colorectal cancer progression and chemoradiotherapy. Biomed. Pharmacother. 2021, 134, 111099. [Google Scholar] [CrossRef]
- Milanesi, E.; Dobre, M.; Bucuroiu, A.I.; Herlea, V.; Manuc, T.E.; Salvi, A.; De Petro, G.; Manuc, M.; Becheanu, G. miRNAs-Based Molecular Signature for KRAS Mutated and Wild Type Colorectal Cancer: An Explorative Study. J. Immunol. Res. 2020, 2020, 4927120. [Google Scholar] [CrossRef]
- Gómez de Cedrón, M.; Ramírez de Molina, A. Microtargeting cancer metabolism: Opening new therapeutic windows based on lipid metabolism. J. Lipid Res. 2016, 57, 193–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobre, M.; Milanesi, E.; Mănuc, T.E.; Arsene, D.E.; Ţieranu, C.G.; Maj, C.; Becheanu, G.; Mănuc, M. Differential Intestinal Mucosa Transcriptomic Biomarkers for Crohn’s Disease and Ulcerative Colitis. J. Immunol. Res. 2018, 2018, 9208274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niculite, C.-M.; Enciu, A.-M.; Hinescu, M.E. CD 36: Focus on Epigenetic and Post-Transcriptional Regulation. Front. Genet. 2019, 10, 680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yao, J.; Shi, H.; Gao, B.; Zhang, L. LncRNA TINCR/microRNA-107/CD36 regulates cell proliferation and apoptosis in colorectal cancer via PPAR signaling pathway based on bioinformatics analysis. Biol. Chem. 2019, 400, 663–675. [Google Scholar] [CrossRef]
- Guo, J.; Fang, W.; Sun, L.; Lu, Y.; Dou, L.; Huang, X.; Tang, W.; Yu, L.; Li, J. Ultraconserved element uc.372 drives hepatic lipid accumulation by suppressing miR-195/miR4668 maturation. Nat. Commun. 2018, 9, 612. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Spetz, M.R.; Ho, M. The Role of Glypicans in Cancer Progression and Therapy. J. Histochem. Cytochem. 2020, 68, 841–862. [Google Scholar] [CrossRef]
- Yu, W.; Ling, J.; Yu, H.; Du, J.; Liu, T. AZGP1 suppresses the process of colorectal cancer after upregulating FASN expression via mTOR signal pathway. Gen. Physiol. Biophys. 2020, 39, 239–248. [Google Scholar] [CrossRef]
- Jafari, N.; Drury, J.; Morris, A.J.; Onono, F.O.; Stevens, P.D.; Gao, T.; Liu, J.; Wang, C.; Lee, E.Y.; Weiss, H.L.; et al. De Novo Fatty Acid Synthesis-Driven Sphingolipid Metabolism Promotes Metastatic Potential of Colorectal Cancer. Mol. Cancer Res. 2019, 17, 140–152. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.; Sun, L.; Wang, Z.; Zhang, Y.; He, Z.; Xu, C. Fatty acid synthase enhances colorectal cancer cell proliferation and metastasis via regulating AMPK/mTOR pathway. Onco Targets Ther. 2019, 12, 3339–3347. [Google Scholar] [CrossRef] [Green Version]
- Zaytseva, Y.Y.; Rychahou, P.G.; Gulhati, P.; Elliott, V.A.; Mustain, W.C.; O’Connor, K.; Morris, A.J.; Sunkara, M.; Weiss, H.L.; Lee, E.Y.; et al. Inhibition of fatty acid synthase attenuates CD44-associated signaling and reduces metastasis in colorectal cancer. Cancer Res. 2012, 72, 1504–1517. [Google Scholar] [CrossRef]
- Ventura, R.; Mordec, K.; Waszczuk, J.; Wang, Z.; Lai, J.; Fridlib, M.; Buckley, D.; Kemble, G.; Heuer, T.S. Inhibition of de novo Palmitate Synthesis by Fatty Acid Synthase Induces Apoptosis in Tumor Cells by Remodeling Cell Membranes, Inhibiting Signaling Pathways, and Reprogramming Gene Expression. EBioMedicine 2015, 2, 808–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.H.; Lee, M.S.; Cha, E.Y.; Sul, J.Y.; Lee, J.S.; Kim, J.S.; Park, J.B.; Kim, J.Y. Inhibitory effect of emodin on fatty acid synthase, colon cancer proliferation and apoptosis. Mol. Med. Rep. 2017, 15, 2163–2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaytseva, Y.Y.; Rychahou, P.G.; Le, A.-T.; Scott, T.L.; Flight, R.M.; Kim, J.T.; Harris, J.; Liu, J.; Wang, C.; Morris, A.J.; et al. Preclinical evaluation of novel fatty acid synthase inhibitors in primary colorectal cancer cells and a patient-derived xenograft model of colorectal cancer. Oncotarget 2018, 9, 24787–24800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Guo, D.; He, J.; Song, L.; Chen, H.; Zhang, Z.; Tan, N. Inhibition of fatty acid synthesis arrests colorectal neoplasm growth and metastasis: Anti-cancer therapeutical effects of natural cyclopeptide RA-XII. Biochem. Biophys. Res. Commun. 2019, 512, 819–824. [Google Scholar] [CrossRef]
- Czumaj, A.; Zabielska, J.; Pakiet, A.; Mika, A.; Rostkowska, O.; Makarewicz, W.; Kobiela, J.; Sledzinski, T.; Stelmanska, E. In Vivo Effectiveness of Orlistat in the Suppression of Human Colorectal Cancer Cell Proliferation. Anticancer Res. 2019, 39, 3815–3822. [Google Scholar] [CrossRef]
- Rapuano, R.; Ziccardi, P.; Cioffi, V.; Dallavalle, S.; Moricca, S.; Lupo, A. Cladosporols A and B, two natural peroxisome proliferator-activated receptor gamma (PPARγ) agonists, inhibit adipogenesis in 3T3-L1 preadipocytes and cause a conditioned-culture-medium-dependent arrest of HT-29 cell proliferation. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129973. [Google Scholar] [CrossRef]
- Qian, J.; Jiang, B.; Li, M.; Chen, J.; Fang, M. Prognostic significance of microRNA-16 expression in human colorectal cancer. World J. Surg. 2013, 37, 2944–2949. [Google Scholar] [CrossRef]
- Xiao, G.; Tang, H.; Wei, W.; Li, J.; Ji, L.; Ge, J. Aberrant Expression of MicroRNA-15a and MicroRNA-16 Synergistically Associates with Tumor Progression and Prognosis in Patients with Colorectal Cancer. Gastroenterol. Res. Pract. 2014, 2014, 364549. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Shen, L.; Li, F.; Yang, J.; Wan, X.; Ouyang, M. microRNA-16-5p-containing exosomes derived from bone marrow-derived mesenchymal stem cells inhibit proliferation, migration, and invasion, while promoting apoptosis of colorectal cancer cells by downregulating ITGA2. J. Cell. Physiol. 2019, 234, 21380–21394. [Google Scholar] [CrossRef]
- Huang, X.; Xu, X.; Ke, H.; Pan, X.; Ai, J.; Xie, R.; Lan, G.; Hu, Y.; Wu, Y. microRNA-16-5p suppresses cell proliferation and angiogenesis in colorectal cancer by negatively regulating forkhead box K1 to block the PI3K/Akt/mTOR pathway. Eur. J. Histochem. 2022, 66, 3333. [Google Scholar] [CrossRef]
- Diamantopoulos, M.A.; Kontos, C.K.; Kerimis, D.; Papadopoulos, I.N.; Scorilas, A. Upregulated miR-16 expression is an independent indicator of relapse and poor overall survival of colorectal adenocarcinoma patients. Clin. Chem. Lab. Med. 2017, 55, 737–747. [Google Scholar] [CrossRef]
- Hasáková, K.; Bezakova, J.; Vician, M.; Reis, R.; Zeman, M.; Herichova, I. Gender-dependent expression of leading and passenger strand of miR-21 and miR-16 in human colorectal cancer and adjacent colonic tissues. Physiol. Res. 2017, 66, S575–S582. [Google Scholar] [CrossRef] [PubMed]
- Earle, J.S.L.; Luthra, R.; Romans, A.; Abraham, R.; Ensor, J.; Yao, H.; Hamilton, S.R. Association of microRNA expression with microsatellite instability status in colorectal adenocarcinoma. J. Mol. Diagn. 2010, 12, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Tong, J.; Huang, G. Nicotinamide phosphoribosyl transferase (Nampt) is a target of microRNA-26b in colorectal cancer cells. PLoS ONE 2013, 8, e69963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Li, J.; Yan, X.; Jin, K.; Li, W.; Liu, X.; Zhao, J.; Shang, W.; Liu, Y. Long Noncoding RNA Plasmacytoma Variant Translocation 1 (PVT1) Promotes Colon Cancer Progression via Endogenous Sponging miR-26b. Med. Sci. Monit. 2018, 24, 8685–8692. [Google Scholar] [CrossRef]
- Ma, Y.-L.; Zhang, P.; Wang, F.; Moyer, M.P.; Yang, J.-J.; Liu, Z.-H.; Peng, J.-Y.; Chen, H.-Q.; Zhou, Y.-K.; Liu, W.-J.; et al. Human embryonic stem cells and metastatic colorectal cancer cells shared the common endogenous human microRNA-26b. J. Cell. Mol. Med. 2011, 15, 1941–1954. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sun, Z.; Liu, B.; Shan, Y.; Zhao, L.; Jia, L. Tumor-suppressive miR-26a and miR-26b inhibit cell aggressiveness by regulating FUT4 in colorectal cancer. Cell Death Dis. 2017, 8, e2892. [Google Scholar] [CrossRef] [Green Version]
- Cristóbal, I.; Manso, R.; Gónzález-Alonso, P.; Madoz-Gúrpide, J.; Rojo, F.; García-Foncillas, J. Clinical Value of miR-26b Discriminating Ulcerative Colitis-associated Colorectal Cancer in the Subgroup of Patients with Metastatic Disease. Inflamm. Bowel Dis. 2015, 21, E24–E25. [Google Scholar] [CrossRef] [Green Version]
- Fan, D.; Lin, X.; Zhang, F.; Zhong, W.; Hu, J.; Chen, Y.; Cai, Z.; Zou, Y.; He, X.; Chen, X.; et al. MicroRNA 26b promotes colorectal cancer metastasis by downregulating phosphatase and tensin homolog and wingless-type MMTV integration site family member 5A. Cancer Sci. 2018, 109, 354–362. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Lin, L.; Xia, L. MiR-107 function as a tumor suppressor gene in colorectal cancer by targeting transferrin receptor 1. Cell. Mol. Biol. Lett. 2019, 24, 31. [Google Scholar] [CrossRef]
- Liu, F.; Liu, S.; Ai, F.; Zhang, D.; Xiao, Z.; Nie, X.; Fu, Y. miR-107 Promotes Proliferation and Inhibits Apoptosis of Colon Cancer Cells by Targeting Prostate Apoptosis Response-4 (Par4). Oncol. Res. 2017, 25, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Li, H.; Zhou, J. Long non-coding RNA MIR503HG inhibits the proliferation, migration and invasion of colon cancer cells via miR-107/Par4 axis. Exp. Cell Res. 2020, 395, 112205. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Ma, J.; Zhang, L. Integrin Subunit Alpha 5 (ITGA5) Gene Circular RNA Sponges microRNA-107 in Colorectal Carcinoma Cells and Tissues and Regulates the Expression of the Forkhead Box J3 (FOXJ3) Gene. Med. Sci. Monit. 2020, 26, e920623. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Liang, X.; Li, X.; Zhang, Y.; Sun, Z.; Liu, Y.; Wang, J. MicroRNA-195: A review of its role in cancers. Onco Targets Ther. 2018, 11, 7109–7123. [Google Scholar] [CrossRef] [Green Version]
- Poel, D.; Boyd, L.N.C.; Beekhof, R.; Schelfhorst, T.; Pham, T.V.; Piersma, S.R.; Knol, J.C.; Jimenez, C.R.; Verheul, H.M.W.; Buffart, T.E. Proteomic Analysis of miR-195 and miR-497 Replacement Reveals Potential Candidates that Increase Sensitivity to Oxaliplatin in MSI/P53wt Colorectal Cancer Cells. Cells 2019, 8, 1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Q.; Zhang, Z.; Dai, Z.; Basnet, S.; Li, S.; Xu, B.; Ge, H. Tumor-suppressive microRNA-195-5p regulates cell growth and inhibits cell cycle by targeting cyclin dependent kinase 8 in colon cancer. Am. J. Transl. Res. 2016, 8, 2088–2096. [Google Scholar]
- Lin, X.; Wang, S.; Sun, M.; Zhang, C.; Wei, C.; Yang, C.; Dou, R.; Liu, Q.; Xiong, B. miR-195-5p/NOTCH2-mediated EMT modulates IL-4 secretion in colorectal cancer to affect M2-like TAM polarization. J. Hematol. Oncol. 2019, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Cojocneanu, R.; Braicu, C.; Raduly, L.; Jurj, A.; Zanoaga, O.; Magdo, L.; Irimie, A.; Muresan, M.-S.; Ionescu, C.; Grigorescu, M.; et al. Plasma and Tissue Specific miRNA Expression Pattern and Functional Analysis Associated to Colorectal Cancer Patients. Cancers 2020, 12, 843. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Song, H.; Wang, S.; Zhang, C.; Zheng, L.; Chen, F.; Shi, D.; Chen, Y.; Yang, C.; Xiang, Z.; et al. Integrated analysis identifies microRNA-195 as a suppressor of Hippo-YAP pathway in colorectal cancer. J. Hematol. Oncol. 2017, 10, 79. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xu, M.; Ding, L.; Tang, J. MiR-27a: A Novel Biomarker and Potential Therapeutic Target in Tumors. J. Cancer 2019, 10, 2836–2848. [Google Scholar] [CrossRef] [Green Version]
- Colangelo, T.; Polcaro, G.; Ziccardi, P.; Pucci, B.; Muccillo, L.; Galgani, M.; Fucci, A.; Milone, M.R.; Budillon, A.; Santopaolo, M.; et al. Proteomic screening identifies calreticulin as a miR-27a direct target repressing MHC class I cell surface exposure in colorectal cancer. Cell Death Dis. 2016, 7, e2120. [Google Scholar] [CrossRef] [PubMed]
- Kara, M.; Yumrutas, O.; Ozcan, O.; Celik, O.I.; Bozgeyik, E.; Bozgeyik, I.; Tasdemir, S. Differential expressions of cancer-associated genes and their regulatory miRNAs in colorectal carcinoma. Gene 2015, 567, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Vychytilova-Faltejskova, P.; Radova, L.; Sachlova, M.; Kosarova, Z.; Slaba, K.; Fabian, P.; Grolich, T.; Prochazka, V.; Kala, Z.; Svoboda, M.; et al. Serum-based microRNA signatures in early diagnosis and prognosis prediction of colon cancer. Carcinogenesis 2016, 37, 941–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Sun, Z.; Sun, H.; Li, Y. MicroRNA-27a promotes tumorigenesis via targeting AKT in triple negative breast cancer. Mol. Med. Rep. 2018, 17, 562–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.-Z.; Wang, K.-P.; Song, H.-J.; Xia, J.-H.; Jiang, Y.; Wang, Y.-L. MiR-27a-3p promotes esophageal cancer cell proliferation via F-box and WD repeat domain-containing 7 (FBXW7) suppression. Int. J. Clin. Exp. Med. 2015, 8, 15556–15562. [Google Scholar]
- Su, C.; Huang, D.-P.; Liu, J.-W.; Liu, W.-Y.; Cao, Y.-O. miR-27a-3p regulates proliferation and apoptosis of colon cancer cells by potentially targeting BTG1. Oncol. Lett. 2019, 18, 2825–2834. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Kim, A.Y.; Lee, H.W.; Son, Y.H.; Lee, G.Y.; Lee, J.-W.; Lee, Y.S.; Kim, J.B. miR-27a is a negative regulator of adipocyte differentiation via suppressing PPARgamma expression. Biochem. Biophys. Res. Commun. 2010, 392, 323–328. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, J.-F.; Chen, W.-J.; Tang, S.-L.; Mo, Z.-C.; Tang, Y.-Y.; Li, Y.; Wang, J.-L.; Liu, X.-Y.; Peng, J.; et al. MicroRNA-27a/b regulates cellular cholesterol efflux, influx and esterification/hydrolysis in THP-1 macrophages. Atherosclerosis 2014, 234, 54–64. [Google Scholar] [CrossRef]
- Cui, Q. Significance of miR-27a and miR-31 in early diagnosis and prognosis of colorectal cancer. Oncol. Lett. 2019, 18, 3092–3096. [Google Scholar] [CrossRef]
- Barisciano, G.; Colangelo, T.; Rosato, V.; Muccillo, L.; Taddei, M.L.; Ippolito, L.; Chiarugi, P.; Galgani, M.; Bruzzaniti, S.; Matarese, G.; et al. miR-27a is a master regulator of metabolic reprogramming and chemoresistance in colorectal cancer. Br. J. Cancer 2020, 122, 1354–1366. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zhu, D.; Hou, L.; Wang, Y.; Huang, X.; Zhou, C.; Zhu, L.; Wang, Y.; Li, L.; Gu, Y.; et al. MiR-107 confers chemoresistance to colorectal cancer by targeting calcium-binding protein 39. Br. J. Cancer 2020, 122, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Wang, M.; Hu, H.; Huang, Q.; Chen, Y.; Wang, G. Overcoming stemness and chemoresistance in colorectal cancer through miR-195-5p-modulated inhibition of notch signaling. Int. J. Biol. Macromol. 2018, 117, 445–453. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | CRC Patients (n = 39) |
|---|---|
| Age (mean ± SD) | 65.62 ± 11.00 |
| Sex | 21F; 18M |
| Moderate alcohol use * | 46.6% Yes; 53.4% No |
| Obesity * | 13.3% Yes; 86.7% No |
| Diabetes * | 10% Yes; 90% No |
| Tumor Characteristics | |
| Localization | Ascending colon (n = 5) Transverse colon (n = 5) Descending colon (n = 5) Sigmoid (n = 9) RSJ (n = 9) Rectum (n = 6) |
| Grade | G1 (n = 6) G2 (n = 26) G3 (n = 7) |
| TNM (T = size of the primary tumor; N = degree of spread to regional lymph nodes; M = presence of distant metastasis) | T2 N0 M0 (n = 4); T2 N1 M0 (n = 1); T2 N2 M0 (n = 1); T3 N0 M0 (n = 16); T3 N0 M1 (n = 1); T3 N1 M0 (n = 7); T3 N2 M0 (n = 4); T4 N0 M0 (n = 3); T4 N1 M0 (n = 1); T4 N2 M0 (n = 1); |
| Lymphovascular invasion | Yes (n = 10); No (n = 29) |
| Perineural invasion | Yes (n = 6); No (n = 33) |
| Tumoral Markers | |
| CEA—carcinoembryonic antigen (ng/mL) mean (min–max) (n = 26) | 28.08 (0.65–444) |
| CA 19-9—carbohydrate antigen (U/mL) mean (min–max) (n = 26) | 35.58 (3.47–291.25) |
| AFP—alpha-fetoprotein (ng/mL) mean (min–max) (n = 20) | 3.70 (0.60–11.84) |
| Biochemical Parameters | |
| Hemoglobin (g/dL) mean (min–max) (n = 36) | 12.37 (6.90–17.90) |
| WBC—white blood cells (N/µL) mean (min–max) (n = 36) | 6377 (1016–14840) |
| Platelets (N103/µL) mean (min–max) (n = 36) | 303 (152–777) |
| INR—international normalized ratio mean (min–max) (n = 36) | 1.04 (0.87–1.58) |
| Fibrinogen (mg/dL) mean (min–max) (n = 32) | 426.58 (138–796) |
| Albumin (g/dL) mean (min–max) (n = 31) | 3.93 (2.30–5.48) |
| Genes | 39 T vs. 39 PT | 39 PT vs. 18 CTRL | 39 T vs. 18 CTRL | 308 T vs. 41 CTRL (OncoDB) | ||||
|---|---|---|---|---|---|---|---|---|
| p-Value * | FR | p-Value ** | FR | p-Value ** | FR | p-Value $ | FR | |
| CD36 | <0.001 | −5.13 | 0.223 | 1.47 | <0.001 | −3.49 | <0.001 | −3.79 |
| FASN | 0.003 | 1.41 | <0.001 | 2.97 | <0.001 | 4.2 | <0.001 | 2.53 |
| SLC27A3 | <0.001 | −1.77 | <0.001 | 2.18 | 0.46 | 1.23 | 0.015 | −1.16 |
| SLC27A4 | <0.001 | −1.73 | 0.81 | −1.12 | 0.001 | −1.94 | <0.001 | −1.77 |
| GPC4 | 0.586 | −1.38 | 0.027 | 1.88 | 0.107 | 1.37 | <0.001 | 1.63 |
| miRNAs | T vs. PT | T vs. CTRL | PT vs. CTRL | |||
|---|---|---|---|---|---|---|
| p-Value * | FR | p-Value ** | FR | p-Value ** | FR | |
| miR-155-5p | 0.339 | −1.24 | 0.127 | −1.14 | 0.768 | 1.09 |
| miR-16-5p | 0.009 | −1.31 | 0.290 | 1.09 | 0.313 | 1.42 |
| miR-27a-3p | 0.065 | −1.38 | 0.025 | 1.79 | 0.002 | 2.48 |
| miR-26b-5p | <0.001 | −1.83 | 0.052 | −1.23 | 0.127 | 1.48 |
| miR-29a-3p | 0.737 | 1.11 | 0.140 | 1.38 | 0.389 | 1.24 |
| miR-107 | 0.008 | −1.45 | 0.200 | −1.14 | 0.257 | 1.27 |
| miR-195-5p | <0.001 | −5.78 | 0.001 | −1.86 | 0.001 | 3.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niculae, A.M.; Dobre, M.; Herlea, V.; Vasilescu, F.; Ceafalan, L.C.; Trandafir, B.; Milanesi, E.; Hinescu, M.E. Lipid Handling Protein Gene Expression in Colorectal Cancer: CD36 and Targeting miRNAs. Life 2022, 12, 2127. https://doi.org/10.3390/life12122127
Niculae AM, Dobre M, Herlea V, Vasilescu F, Ceafalan LC, Trandafir B, Milanesi E, Hinescu ME. Lipid Handling Protein Gene Expression in Colorectal Cancer: CD36 and Targeting miRNAs. Life. 2022; 12(12):2127. https://doi.org/10.3390/life12122127
Chicago/Turabian StyleNiculae, Andrei Marian, Maria Dobre, Vlad Herlea, Florina Vasilescu, Laura Cristina Ceafalan, Bogdan Trandafir, Elena Milanesi, and Mihail Eugen Hinescu. 2022. "Lipid Handling Protein Gene Expression in Colorectal Cancer: CD36 and Targeting miRNAs" Life 12, no. 12: 2127. https://doi.org/10.3390/life12122127