

Bacterial and Fungal Co-Occurrence in the Nudibranch, Pteraeolidia semperi

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. DNA Extraction and Amplification
2.2. Bioinformatics
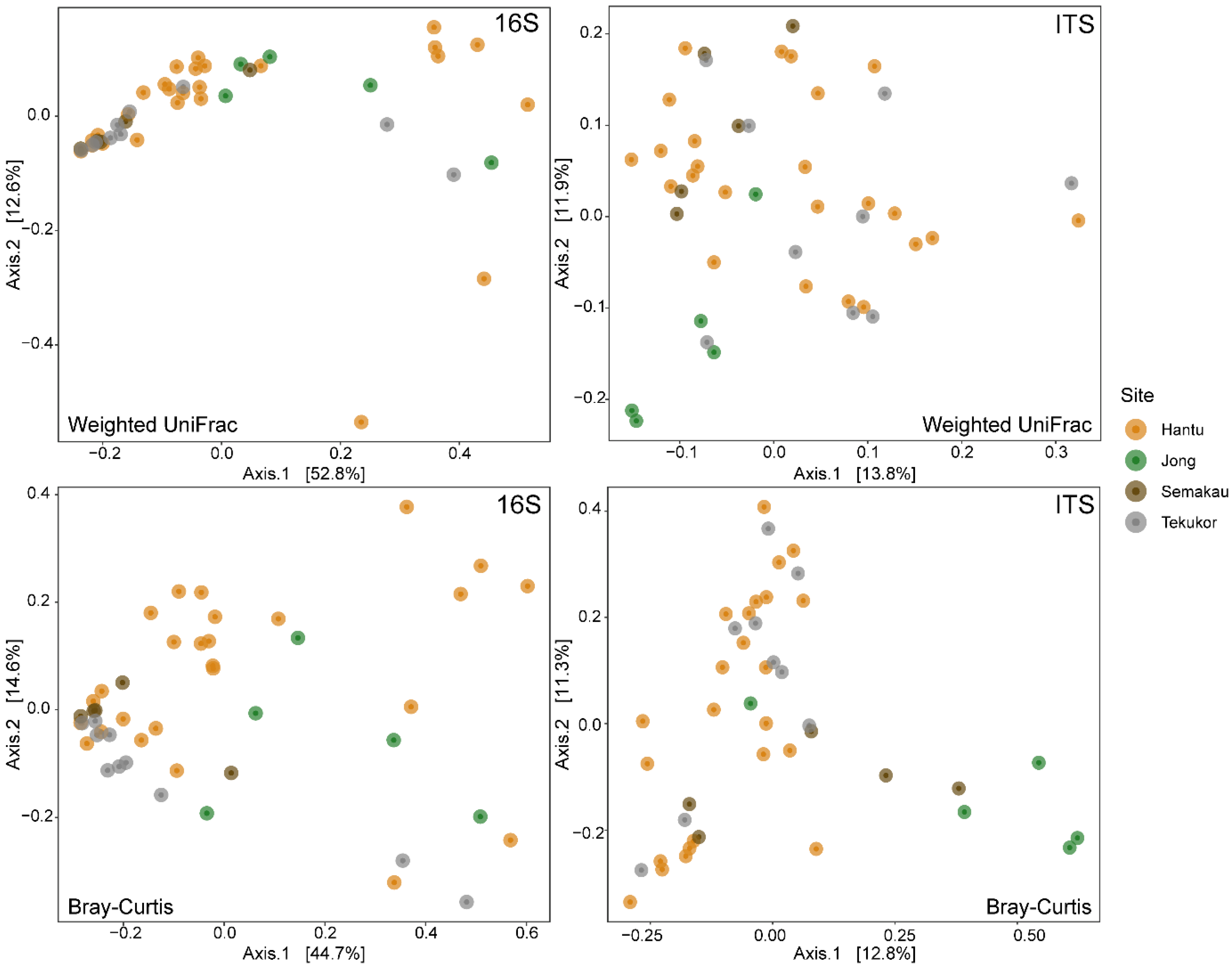
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Amend, A.S.; Swift, S.O.I.; Darcy, J.L.; Belcaid, M.; Nelson, C.E.; Buchanan, J.; Cetraro, N.; Fraiola, K.M.S.; Frank, K.; Kajihara, K.; et al. A Ridge-to-Reef Ecosystem Microbial Census Reveals Environmental Reservoirs for Animal and Plant Microbiomes. Proc. Natl. Acad. Sci. USA 2022, 119, e2204146119. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ Warning to Humanity: Microorganisms and Climate Change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilber-Rosenberg, I.; Rosenberg, E. Role of Microorganisms in the Evolution of Animals and Plants: The Hologenome Theory of Evolution. FEMS Microbiol. Rev. 2008, 32, 723–735. [Google Scholar] [CrossRef]
- Kanisan, D.P.; Quek, Z.B.R.; Oh, R.M.; Afiq-Rosli, L.; Lee, J.N.; Huang, D.; Wainwright, B.J. Diversity and Distribution of Microbial Communities Associated with Reef Corals of the Malay Peninsula. Microb. Ecol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.L.Y.; Huang, D.; Quek, Z.B.R.; Lee, J.N.; Wainwright, B.J. Mangrove-Associated Fungal Communities Are Differentiated by Geographic Location and Host Structure. Front. Microbiol. 2019, 10, 2456. [Google Scholar] [CrossRef]
- Deveau, A.; Bonito, G.; Uehling, J.; Paoletti, M.; Becker, M.; Bindschedler, S.; Hacquard, S.; Hervé, V.; Labbé, J.; Lastovetsky, O.A.; et al. Bacterial–Fungal Interactions: Ecology, Mechanisms and Challenges. FEMS Microbiol. Rev. 2018, 42, 335–352. [Google Scholar] [CrossRef] [Green Version]
- Risely, A. Applying the Core Microbiome to Understand Host–Microbe Systems. J. Anim. Ecol. 2020, 89, 1549–1558. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Schlaeppi, K.; van der Heijden, M.G. Keystone Taxa as Drivers of Microbiome Structure and Functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef]
- Todd, C.D.; Walter, J.; Daviee, J. Some Perspectives on the Biology and Ecology of Nudibranch Molluscs: Generalisations and Variations on the Theme That Prove the Rule. Boll. Malacol. 2001, 37, 105–120. [Google Scholar]
- Todd, C.D.; Lambert, W.J.; Thorpe, J.P. The Genetic Structure of Intertidal Populations of Two Species of Nudibranch Molluscs with Planktotrophic and Pelagic Lecithotrophic Larval Stages: Are Pelagic Larvae “for” Dispersal? J. Exp. Mar. Bio. Ecol. 1998, 228, 1–28. [Google Scholar] [CrossRef]
- Todd, C.D.; Havenhand, J.N.; Thorpe, J.P. Genetic Differentiation, Pelagic Larval Transport and Gene Flow between Local Populations of the Intertidal Marine Mollusc Adalaria Proxima (Alder & Hancock). Funct. Ecol. 1988, 2, 441–451. [Google Scholar] [CrossRef]
- Chavanich, S.; Viyakarn, V.; Sanpanich, K.; Harris, L.G. Diversity and Occurrence of Nudibranchs in Thailand. Mar. Biodiv. 2013, 43, 31–36. [Google Scholar] [CrossRef]
- Fritts-Penniman, A.L.; Gosliner, T.M.; Mahardika, G.N.; Barber, P.H. Cryptic Ecological and Geographic Diversification in Coral-Associated Nudibranchs. Mol. Phylogenet. Evol. 2020, 144, 106698. [Google Scholar] [CrossRef] [PubMed]
- Carmona, L.; Pola, M.; Gosliner, T.M.; Cervera, J.L. A Tale That Morphology Fails to Tell: A Molecular Phylogeny of Aeolidiidae (Aeolidida, Nudibranchia, Gastropoda). PLoS ONE 2013, 8, e63000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kienberger, K.; Carmona, L.; Pola, M.; Padula, V.; Gosliner, T.M.; Cervera, J.L. Aeolidia papillosa (Linnaeus, 1761) (Mollusca: Heterobranchia: Nudibranchia), Single Species or a Cryptic Species Complex? A Morphological and Molecular Study. Zoo J. Linn. Soc. 2016, 177, 481–506. [Google Scholar] [CrossRef] [Green Version]
- Yorifuji, M.; Takeshima, H.; Mabuchi, K.; Nishida, M. Hidden Diversity in a Reef-Dwelling Sea Slug, Pteraeolidia Ianthina (Nudibranchia, Aeolidina), in the Northwestern Pacific. Jzoo 2012, 29, 359–367. [Google Scholar] [CrossRef]
- Kubanek, J.; Graziani, E.I.; Andersen, R.J. Investigations of Terpenoid Biosynthesis by the Dorid Nudibranch Cadlina luteomarginata. J. Org. Chem. 1997, 62, 7239–7246. [Google Scholar] [CrossRef]
- Winters, A.E.; White, A.M.; Dewi, A.S.; Mudianta, I.W.; Wilson, N.G.; Forster, L.C.; Garson, M.J.; Cheney, K.L. Distribution of Defensive Metabolites in Nudibranch Molluscs. J. Chem. Ecol. 2018, 44, 384–396. [Google Scholar] [CrossRef]
- Böhringer, N.; Fisch, K.M.; Schillo, D.; Bara, R.; Hertzer, C.; Grein, F.; Eisenbarth, J.-H.; Kaligis, F.; Schneider, T.; Wägele, H.; et al. Antimicrobial Potential of Bacteria Associated with Marine Sea Slugs from North Sulawesi, Indonesia. Front. Microbiol. 2017, 8, 1092. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahman, S.M.; Patin, N.V.; Hanora, A.; Aboseidah, A.; Desoky, S.; Desoky, S.G.; Stewart, F.J.; Lopanik, N.B. The Natural Product Biosynthetic Potential of Red Sea Nudibranch Microbiomes. PeerJ 2021, 9, e10525. [Google Scholar] [CrossRef]
- Abdelrahman, S.M.; Dosoky, N.S.; Hanora, A.M.; Lopanik, N.B. Metabolomic Profiling and Molecular Networking of Nudibranch-Associated Streptomyces sp. SCSIO 001680. Molecules 2022, 27, 4542. [Google Scholar] [CrossRef] [PubMed]
- Kristiana, R.; Sibero, M.T.; Farisa, M.Y.; Ayuningrum, D.; Dirgantara, D.; Hanafi, M.; Radjasa, O.K.; Sabdono, A.; Trianto, A. Antibacterial Potential of Nudibranch-Associated Bacteria from Saparua and Nusa Laut Islands, Indonesia. Biodivers. J. Biol. Divers. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Zhukova, N.V.; Eliseikina, M.G.; Balakirev, E.S.; Ayala, F.J. Multiple Bacterial Partners in Symbiosis with the Nudibranch Mollusk Rostanga alisae. Sci. Rep. 2022, 12, 169. [Google Scholar] [CrossRef] [PubMed]
- Zhukova, N.V.; Eliseikina, M.G. Symbiotic Bacteria in the Nudibranch Mollusk Dendrodoris Nigra: Fatty Acid Composition and Ultrastructure Analysis. Mar. Biol. 2012, 159, 1783–1794. [Google Scholar] [CrossRef]
- Austin, B.; Zhang, X.-H. Vibrio harveyi: A Significant Pathogen of Marine Vertebrates and Invertebrates. Lett. Appl. Microbiol. 2006, 43, 119–124. [Google Scholar] [CrossRef]
- Oliver, J.D. Wound Infections Caused by Vibrio Vulnificus and Other Marine Bacteria. Epidemiol. Infect. 2005, 133, 383–391. [Google Scholar] [CrossRef]
- Trianto, A.; Radjasa, O.K.; Sibero, M.T.; Sabdono, A.; Haryanti, D.; Zilullah, W.O.M.; Syanindyta, A.R.; Bahry, M.S.; Widiananto, P.A.; Helmi, M.; et al. The Effect of Culture Media on the Number and Bioactivity of Marine Invertebrates Associated Fungi. Biodiversitas 2020, 21. [Google Scholar] [CrossRef] [Green Version]
- Neu, A.T.; Allen, E.E.; Roy, K. Defining and Quantifying the Core Microbiome: Challenges and Prospects. Proc. Natl. Acad. Sci. USA 2021, 118, e2104429118. [Google Scholar] [CrossRef]
- Bergh, R. Malacologische Untersuchungen; Kreidel: Wiesbaden, Germany, 2009; Volume 1, pp. 1–592. [Google Scholar]
- Dikou, A.; van Woesik, R. Survival under Chronic Stress from Sediment Load: Spatial Patterns of Hard Coral Communities in the Southern Islands of Singapore. Mar. Pollut. Bull. 2006, 52, 1340–1354. [Google Scholar] [CrossRef]
- Januchowski-Hartley, F.A.; Bauman, A.G.; Morgan, K.M.; Seah, J.C.L.; Huang, D.; Todd, P.A. Accreting Coral Reefs in a Highly Urbanized Environment. Coral Reefs 2020, 39, 717–731. [Google Scholar] [CrossRef]
- Goh, B.P.L.; Lim, D.Y.F. Distribution and Abundance of Sea Urchins in Singapore Reefs and Their Potential Ecological Impacts on Macroalgae and Coral Communities. Ocean Sci. J. 2015, 50, 211–219. [Google Scholar] [CrossRef]
- Gosliner, T.M.; Valdés, A.; Behrens, D.W. Nudibranch and Sea Slugs Identification; Jacksonville, F.L., Ed.; New World Publications: Jacksonville, FL, USA, 2018. [Google Scholar]
- Soon, N.; Quek, R.; Pohl, S.; Wainwright, B.J. Symbiodiniaceae Communities in the Reef-Dwelling Pteraeolidia “semperi” (Nudibranchia: Aeolidioidea) Species Complex in Singapore. Available online: https://www.nathanielsoon.co/research (accessed on 15 June 2022).
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global Patterns of 16S RRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, R.M.; Bollati, E.; Maithani, P.; Huang, D.; Wainwright, B.J. The Microbiome of the Reef Macroalga Sargassum Ilicifolium in Singapore. Microorganisms 2021, 9, 898. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. 38—Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Smith, D.P.; Peay, K.G. Sequence Depth, Not PCR Replication, Improves Ecological Inference from Next Generation DNA Sequencing. PLoS ONE 2014, 9, e90234. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; Version 4.0.4; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple Statistical Identification and Removal of Contaminant Sequences in Marker-Gene and Metagenomics Data. Microbiome 2018, 6, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2013, 2, 321–326. [Google Scholar]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An Effective Distance Metric for Microbial Community Comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 2018, 1, 3. [Google Scholar]
- Martinez Arbizu, P. PairwiseAdonis: Pairwise Multilevel Comparison Using Adonis. R Package Version 0.4 2020. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 17 June 2022).
- Lahti, L.; Shetty, S.; Blake, T.; Salojarvi, J. Tools for Microbiome Analysis in R Version 2.1.26. 2017. Available online: https://microbiome.github.io/tutorials/ (accessed on 15 June 2022).
- Kurtz, Z.D.; Müller, C.L.; Miraldi, E.R.; Littman, D.R.; Blaser, M.J.; Bonneau, R.A. Sparse and Compositionally Robust Inference of Microbial Ecological Networks. PLoS Comp. Biol. 2015, 11, e1004226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. In Proceedings of the International AAAI Conference on Web and Social Media, San Jose, CA, USA, 17–20 May 2009; Volume 3, pp. 361–362. [Google Scholar]
- Birt, H.W.G.; Dennis, P.G. Inference and Analysis of SPIEC-EASI Microbiome Networks. In The Plant Microbiome: Methods and Protocols, Methods in Molecular Biology; Carvalhais, L.C., Dennis, P.G., Eds.; Springer: New York, NY, USA, 2021; pp. 155–171. ISBN 978-1-07-161040-4. [Google Scholar]
- Berry, D.; Widder, S. Deciphering Microbial Interactions and Detecting Keystone Species with Co-Occurrence Networks. Front. Microbiol. 2014, 5, 219. [Google Scholar] [CrossRef] [Green Version]
- Noirungsee, N.; Hackbusch, S.; Viamonte, J.; Bubenheim, P.; Liese, A.; Müller, R. Influence of Oil, Dispersant, and Pressure on Microbial Communities from the Gulf of Mexico. Sci. Rep. 2020, 10, 7079. [Google Scholar] [CrossRef] [PubMed]
- Nogi, Y.; Abe, M.; Kawagucci, S.; Hirayama, H. 2014 Psychrobium Conchae Gen. Nov., Sp. Nov., a Psychrophilic Marine Bacterium Isolated from the Iheya North Hydrothermal Field. Int. J. Syst. Evol. Microbiol. 2014, 64, 3668–3675. [Google Scholar] [CrossRef] [PubMed]
- Vinjusha, N.; ArunKumar, T.K. Neofomitella guangxiensis and Sebipora aquosa Newly Recorded White Rot Polypores from India. Stud. Fungi 2021, 6, 307–314. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, S.; Ren, G.; Ji, X.; He, S. Three Species of Wood-Decaying Fungi in Polyporales New to China. Mycotaxon 2017, 132, 29–42. [Google Scholar] [CrossRef]
- Rabbani, G.; Huang, D.; Wainwright, B.J. The Mycobiome of Pocillopora Acuta in Singapore. Coral Reefs 2021, 40, 1419–1427. [Google Scholar] [CrossRef]
- Sin, T.M.; Ang, H.P.; Buurman, J.; Lee, A.C.; Leong, Y.L.; Ooi, S.K.; Steinberg, P.; Teo, S.L.-M. The Urban Marine Environment of Singapore. Reg. Stud. Mar. Sci. 2016, 8, 331–339. [Google Scholar] [CrossRef]
- Wainwright, B.J.; Zahn, G.L.; Zushi, J.; Lee, N.L.Y.; Ooi, J.L.S.; Lee, J.N.; Huang, D. Seagrass-associated Fungal Communities Show Distance Decay of Similarity That Has Implications for Seagrass Management and Restoration. Ecol. Evol. 2019, 9, 11288–11297. [Google Scholar] [CrossRef]
- Freundt, E.A. Changing Concepts in Mycoplasmataceae Host Diversity. Zent. Bakteriol. 1990, 20, 468–475. [Google Scholar]
- Neave, M.J.; Apprill, A.; Ferrier-Pagès, C.; Voolstra, C.R. Diversity and Function of Prevalent Symbiotic Marine Bacteria in the Genus Endozoicomonas. Appl. Microbiol. Biotechnol. 2016, 100, 8315–8324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, F.L.; Iida, T.; Swings, J. Biodiversity of Vibrios. Microbiol. Mol. Biol. Rev. 2004, 68, 403–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, W.; Kim, P.S.; Tak, E.J.; Sung, H.; Shin, N.-R.; Hyun, D.-W.; Whon, T.W.; Kim, H.S.; Lee, J.-Y.; Yun, J.-H.; et al. Host Phylogeny, Habitat, and Diet Are Main Drivers of the Cephalopod and Mollusk Gut Microbiome. Anim. Microbiome 2022, 4, 30. [Google Scholar] [CrossRef]
- Johansson, K.-E.; Pettersson, B. Taxonomy of Mollicutes. In Molecular Biology and Pathogenicity of Mycoplasmas; Razin, S., Herrmann, R., Eds.; Springer: Boston, MA, USA, 2002; pp. 1–29. ISBN 978-0-306-47606-8. [Google Scholar]
- Razin, S.; Yogev, D.; Naot, Y. Molecular Biology and Pathogenicity of Mycoplasmas. Microbiol. Mol. Biol. Rev. 1998, 62, 1094–1156. [Google Scholar] [CrossRef] [PubMed]
- McAuliffe, L.; Ellis, R.J.; Miles, K.; Ayling, R.D.; Nicholas, R.A. Biofilm Formation by Mycoplasma Species and Its Role in Environmental Persistence and Survival. Microbiology 2006, 152, 913–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, H.; Haltli, B.; Duque, C.; Kerr, R. Bacterial Communities of the Gorgonian Octocoral Pseudopterogorgia elisabethae. Microb. Ecol. 2013, 66, 972–985. [Google Scholar] [CrossRef]
- Ziegler, M.; Roik, A.; Porter, A.; Zubier, K.; Mudarris, M.S.; Ormond, R.; Voolstra, C.R. Coral Microbial Community Dynamics in Response to Anthropogenic Impacts near a Major City in the Central Red Sea. Mar. Pollut. Bull. 2016, 105, 629–640. [Google Scholar] [CrossRef] [Green Version]
- Pantos, O.; Bongaerts, P.; Dennis, P.G.; Tyson, G.W.; Hoegh-Guldberg, O. Habitat-Specific Environmental Conditions Primarily Control the Microbiomes of the Coral Seriatopora hystrix. ISME J. 2015, 9, 1916–1927. [Google Scholar] [CrossRef]
- Yang, C.-S.; Chen, M.-H.; Arun, A.B.; Chen, C.A.; Wang, J.-T.; Chen, W.-M. Endozoicomonas montiporae Sp. Nov., Isolated from the Encrusting Pore Coral Montipora aequituberculata. Int. J. Syst. Evol. Microbiol. 2010, 60, 1158–1162. [Google Scholar] [CrossRef]
- Bourne, D.G.; Dennis, P.G.; Uthicke, S.; Soo, R.M.; Tyson, G.W.; Webster, N. Coral Reef Invertebrate Microbiomes Correlate with the Presence of Photosymbionts. ISME J. 2013, 7, 1452–1458. [Google Scholar] [CrossRef]
- Nishijima, M.; Adachi, K.; Katsuta, A.; Shizuri, Y.; Yamasato, K. Endozoicomonas numazuensis sp. Nov., a Gammaproteobacterium Isolated from Marine Sponges, and Emended Description of the Genus Endozoicomonas Kurahashi and Yokota 2007. Int. J. Syst. Evol. Microbiol. 2013, 63, 709–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rola, M.; Frankenbach, S.; Bleidissel, S.; Sickinger, C.; Donath, A.; Frommlet, J.C.; Greve, C.; Serôdio, J.; Preisfeld, A.; Clavijo, J.M. Cladobranchia (Gastropoda, Nudibranchia) as a Promising Model to Understand the Molecular Evolution of Photosymbiosis in Animals. Front. Mar. Sci. 2022. [Google Scholar] [CrossRef]
- Consiglio, G.; Contu, M. Some Rare, Interesting Species of Leucoagaricus subgen. Sericeomyces Micol. Veg. Mediterr. 2004, 19, 57–72. [Google Scholar]
- Gennari, A.; Migliozzi, V.; Virgós, J.L.S. Tre Interessanti Leucocoprinee Sabulicole. Leucoagaricus gaillardii Bon & Biffard, Sericomyces subvolvatus (Malençon & Bertault) Contu fo. Subvolvatus e S. subvolvatus fo. Pictus (Malençon & Bertault) Bon. Micol. E Veg. Mediterr. 1995, 10, 9–21. [Google Scholar]
- Saccardo, P.A.; Traverso, G.B.; Trotter, A. Sylloge Fungorum Omnium Hucusque Cognitorum; Patavii, Sumptibus Auctoris: Padova, Italy, 1882; pp. 1–956. [Google Scholar]
- Kumar, T.A.; Manimohan, P. The Genera Leucoagaricus and Leucocoprinus (Agaricales, Basidiomycota) in Kerala State, India. Mycotaxon 2009, 108, 385–428. [Google Scholar] [CrossRef]
- Ma, Y.; Liu, T.; Yu, X.; Wei, T.; Ge, Z.-W. Six New Species of Leucoagaricus (Agaricaceae) from Northeastern China. Diversity 2022, 14, 314. [Google Scholar] [CrossRef]
- Quinlan, R.J.; Cherrett, J.M. The Role of Fungus in the Diet of the Leaf-cutting Ant Atta Cephalotes (L.). Ecol. Entomol. 1979, 4, 151–160. [Google Scholar] [CrossRef]
- Aylward, F.O.; Burnum-Johnson, K.E.; Tringe, S.G.; Teiling, C.; Tremmel, D.M.; Moeller, J.A.; Scott, J.J.; Barry, K.W.; Piehowski, P.D.; Nicora, C.D. Leucoagaricus gongylophorus Produces Diverse Enzymes for the Degradation of Recalcitrant Plant Polymers in Leaf-Cutter Ant Fungus Gardens. Appl. Environ. Microbiol. 2013, 79, 3770–3778. [Google Scholar] [CrossRef] [Green Version]
- Boonrung, S.; Mongkolthanaruk, W.; Aimi, T.; Boonlue, S. Cellulase and Xylanase Acting at Alkaline PH from Mushroom, Leucoagaricus meleagris KKU-C1. Chiang Mai J. Sci. 2014, 41, 84–96. [Google Scholar]
- Adiwijaya, C.; Bengen, D.G.; Zamani, N.P. Coral Reefs Substrate Composition Influence on Nudibranch Diversity. IOP Conf. Ser. Earth Environ. Sci. 2021, 771, 012009. [Google Scholar] [CrossRef]
- Imbs Andrey, B.; Valeria, P.G. Lipidomic Study of the Influence of Dietary Fatty Acids on Structural Lipids of Cold-Water Nudibranch Molluscs. Sci. Rep. 2019, 9, 20013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Góes-Neto, A.; Marcelino, V.R.; Verbruggen, H.; da Silva, F.F.; Badotti, F. Biodiversity of Endolithic Fungi in Coral Skeletons and Other Reef Substrates Revealed with 18S RDNA Metabarcoding. Coral Reefs 2020, 39, 229–238. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CABI: Wallingford, UK, 2008. [Google Scholar]
- Godinho, V.M.; de Paula, M.T.R.; Silva, D.A.S.; Paresque, K.; Martins, A.P.; Colepicolo, P.; Rosa, C.A.; Rosa, L.H. Diversity and Distribution of Hidden Cultivable Fungi Associated with Marine Animals of Antarctica. Fungal Biol. 2019, 123, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Ogaki, M.B.; Pinto, O.H.B.; Vieira, R.; Neto, A.A.; Convey, P.; Carvalho-Silva, M.; Rosa, C.A.; Câmara, P.E.A.S.; Rosa, L.H. Fungi Present in Antarctic Deep-Sea Sediments Assessed Using DNA Metabarcoding. Microb. Ecol. 2021, 82, 157–164. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.; Hartmann, M. Networking in the Plant Microbiome. PLoS Biol. 2016, 14, e1002378. [Google Scholar] [CrossRef] [Green Version]
- Del Frari, G.; Oliveira, H.; Boavida Ferreira, R. White Rot Fungi (Hymenochaetales) and Esca of Grapevine: Insights from Recent Microbiome Studies. J. Fungi 2021, 7, 770. [Google Scholar] [CrossRef] [PubMed]
- Osono, T. Ecology of Ligninolytic Fungi Associated with Leaf Litter Decomposition. Ecol. Res. 2007, 22, 955–974. [Google Scholar] [CrossRef]
- Bhadury, P.; Bik, H.; Lambshead, J.D.; Austen, M.C.; Smerdon, G.R.; Rogers, A.D. Molecular Diversity of Fungal Phylotypes Co-Amplified Alongside Nematodes from Coastal and Deep-Sea Marine Environments. PLoS ONE 2011, 6, e26445. [Google Scholar] [CrossRef] [Green Version]
- Amend, A.S.; Barshis, D.J.; Oliver, T.A. Coral-Associated Marine Fungi Form Novel Lineages and Heterogeneous Assemblages. ISME J. 2012, 6, 1291–1301. [Google Scholar] [CrossRef] [Green Version]
- Martone, P.T.; Estevez, J.M.; Lu, F.; Ruel, K.; Denny, M.W.; Somerville, C.; Ralph, J. Discovery of Lignin in Seaweed Reveals Convergent Evolution of Cell-Wall Architecture. Curr. Biol. 2009, 19, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lu, Y.; Nawaz, M.Z.; Xu, J. Comparative Genomics Reveals Evidence of Genome Reduction and High Extracellular Protein Degradation Potential in Kangiella. Front. Microbiol. 2018, 9, 1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korzhenkov, A.A.; Toshchakov, S.V.; Golyshina, O.V.; Ferrer, M.; Chernikova, T.N.; Jaeger, K.-E.; Yakimov, M.M.; Golyshin, P.N. Aerobic Hydrocarbon-Degrading Gammaproteobacteria: Oleiphilaceae and Relatives. In Taxonomy, Genomics and Ecophysiology of Hydrocarbon-Degrading Microbes, Handbook of Hydrocarbon and Lipid Microbiology; McGenity, T.J., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 153–166. ISBN 978-3-030-14796-9. [Google Scholar]
- Bowman, J.P. The Family Colwelliaceae. In The Prokaryotes: Gammaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 179–195. ISBN 978-3-642-38922-1. [Google Scholar]
- Ivanova, E.P.; Flavier, S.; Christen, R. Phylogenetic relationships among marine Alteromonas-like proteobacteria: Emended description of the family Alteromonadaceae and proposal of Pseudoalteromonadaceae fam. nov., Colwelliaceae fam. nov., Shewanellaceae fam. nov., Moritellaceae fam. nov., Ferrimonadaceae fam. nov., Idiomarinaceae fam. nov. and Psychromonadaceae fam. nov. Int. J. Syst. Evo. Microbiol. 2014, 54, 1773–1788. [Google Scholar]
- Hwang, D.S.; Masic, A.; Prajatelistia, E.; Iordachescu, M.; Waite, J.H. Marine hydroid perisarc: A chitin-and melanin-reinforced composite with DOPA–iron (III) complexes. Acta Biomater. 2013, 9, 8110–8117. [Google Scholar] [CrossRef] [Green Version]
- Coleman, N. Nudibranchs Encyclopedia: Catalogue of Asia/Indo-Pacific Sea Slugs, 2nd ed.; Masalai Press: Oakland, CA, USA, 2015. [Google Scholar]
- Prabhu, M.; Chemodanov, A.; Gottlieb, R.; Kazir, M.; Nahor, O.; Gozin, M.; Israel, A.; Livney, Y.D.; Golberg, A. Starch from the Sea: The Green Macroalga Ulva Ohnoi as a Potential Source for Sustainable Starch Production in the Marine Biorefinery. Algal Res. 2019, 37, 215–227. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Ng, H.J.; Webb, H.K. The Family Pseudoalteromonadaceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014; pp. 575–582. [Google Scholar]
- Bowman, J.P.; McMeekin, T.A. Alteromonadaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Ltd.: New York, NY, USA, 2015; pp. 1–2. ISBN 978-1-118-96060-8. [Google Scholar]
- Sabdono, A.; Radjasa, O.; Trianto, A.; Korshunova, T.; Martynov, A.; Sibero, M. Diversity and Antimicrobial Activity of Marine Nudibranch Associated Bacteria against Tropical Human Skin Pathogens. F1000Research 2022, 11, 421. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ng, M.S.; Soon, N.; Chang, Y.; Wainwright, B.J. Bacterial and Fungal Co-Occurrence in the Nudibranch, Pteraeolidia semperi. Life 2022, 12, 1988. https://doi.org/10.3390/life12121988
Ng MS, Soon N, Chang Y, Wainwright BJ. Bacterial and Fungal Co-Occurrence in the Nudibranch, Pteraeolidia semperi. Life. 2022; 12(12):1988. https://doi.org/10.3390/life12121988
Chicago/Turabian StyleNg, Ming Sheng, Nathaniel Soon, Ying Chang, and Benjamin J. Wainwright. 2022. "Bacterial and Fungal Co-Occurrence in the Nudibranch, Pteraeolidia semperi" Life 12, no. 12: 1988. https://doi.org/10.3390/life12121988