
Differences in Phenotypic Plasticity between Invasive and Native Plants Responding to Three Environmental Factors
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Experimental Set-Up
2.3. Trait Measurements and Phenotypic Plasticity Index
2.4. Statistical Analysis
3. Results
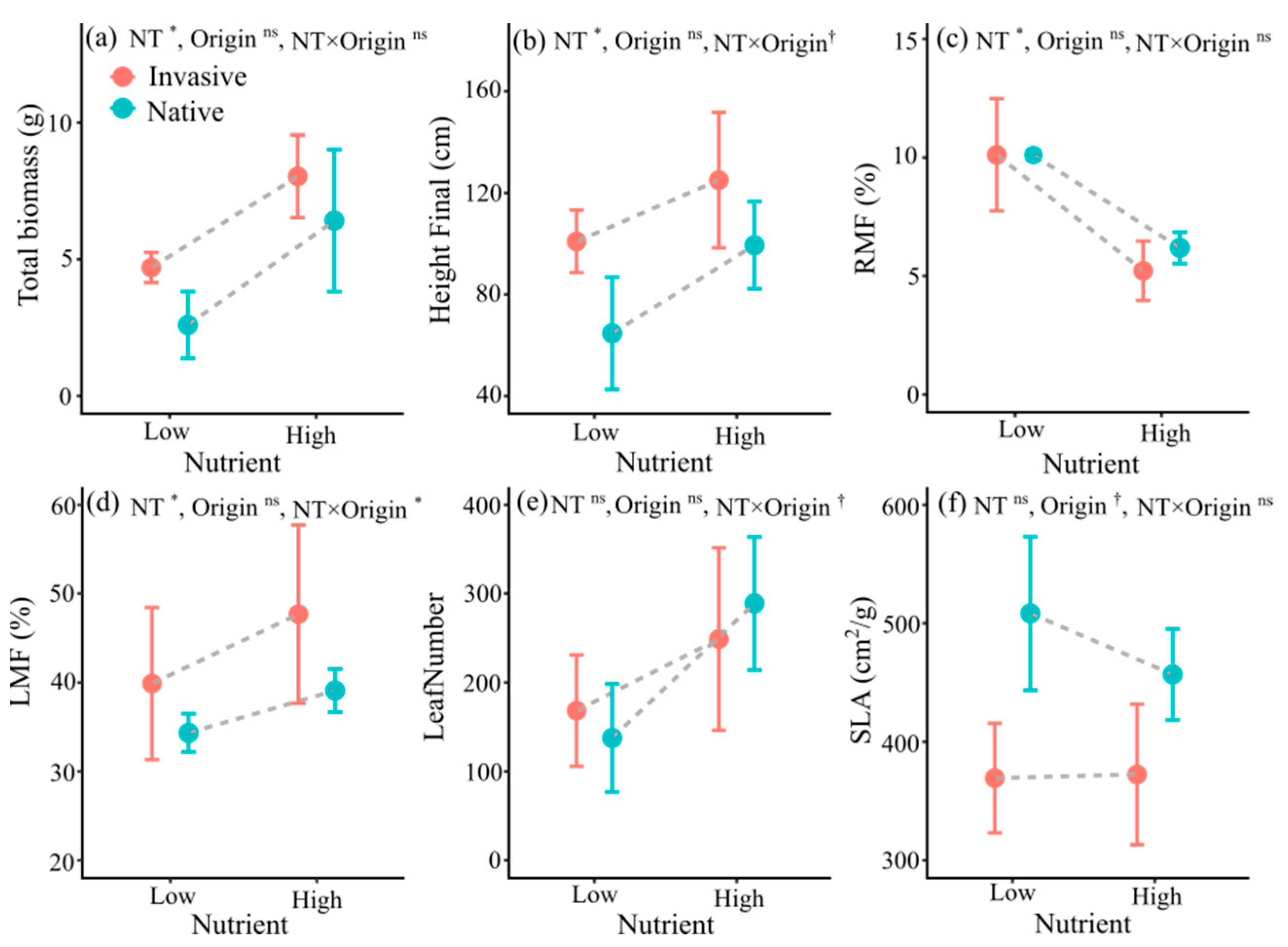
3.1. Trait Responses to Nutrient Treatment
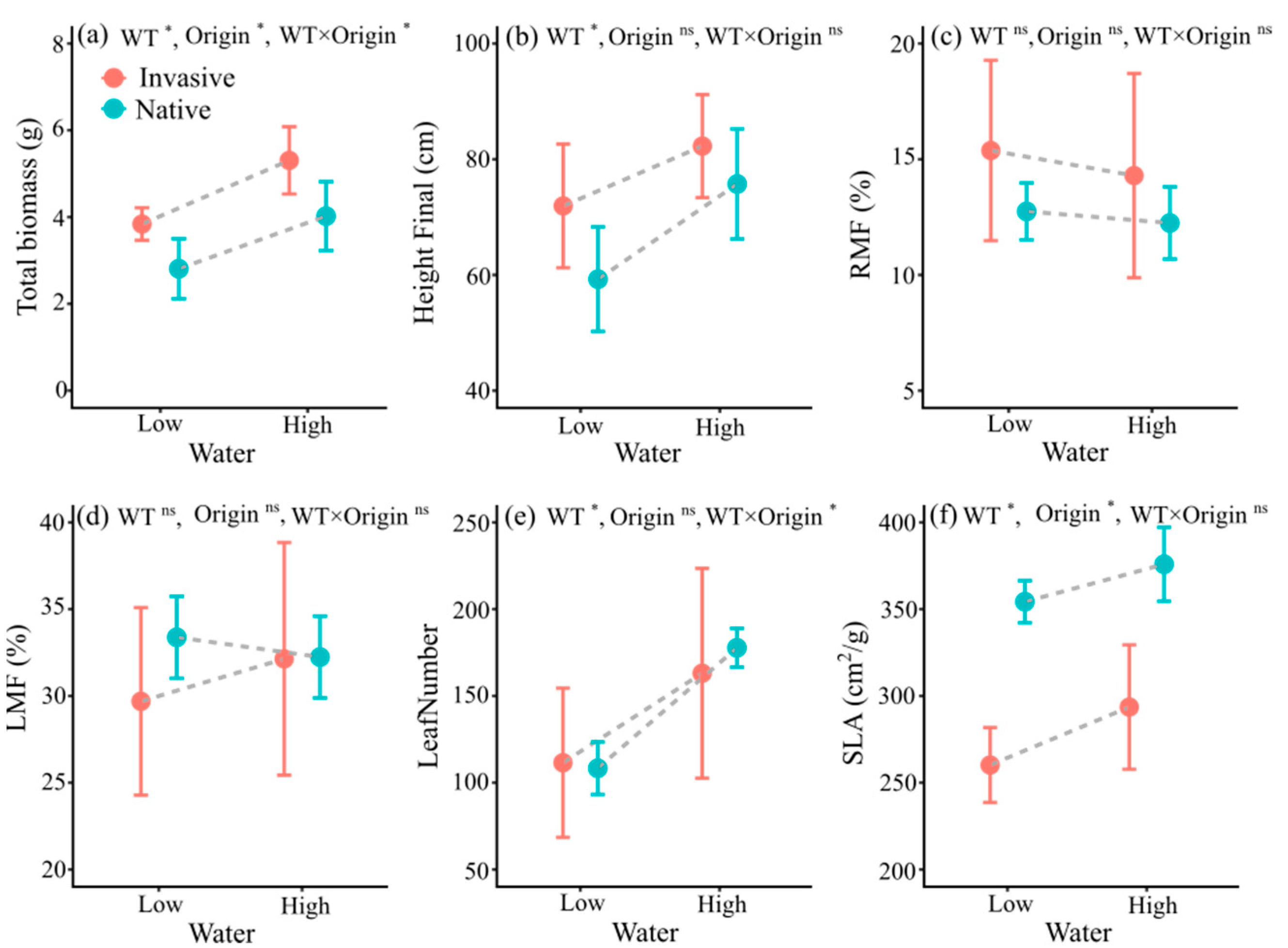
3.2. Trait Responses to Water Treatment
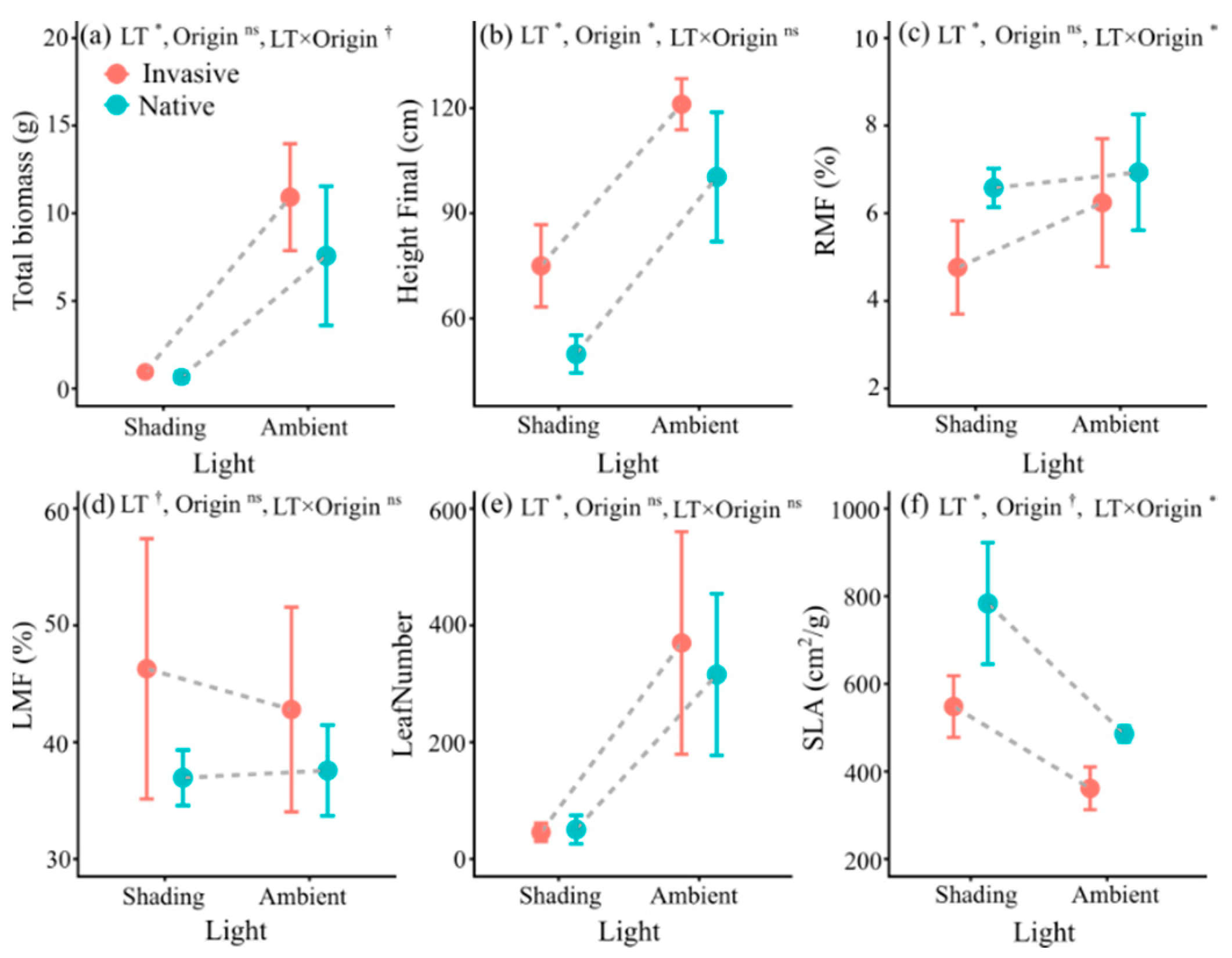
3.3. Trait Responses to Light Treatment
3.4. Variation of Phenotypic Plasticity Index
4. Discussion
4.1. Phenotypic Plasticity of Invasive and Native Plants under Different Nutrient Levels
4.2. Phenotypic Plasticity of Invasive and Native Plants under Different Water Levels
4.3. Phenotypic Plasticity of Invasive and Native Plants under Different Light Levels
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vitousek, P.M.; D’Antonio, C.M.; Loope, L.; Westbrooks, R. Biological invasions as global environmental change. Am. Sci. 1996, 84, 468–478. [Google Scholar]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Christian, C.E. Consequences of a biological invasion reveal the importance of mutualism for plant communities. Nature 2001, 413, 635–639. [Google Scholar] [CrossRef]
- Yang, W.; Zhao, H.; Leng, X.; Cheng, X.; An, S. Soil organic carbon and nitrogen dynamics following Spartina alterniflora invasion in a coastal wetland of eastern China. Catena 2017, 156, 281–289. [Google Scholar] [CrossRef]
- Prass, M.; Ramula, S.; Jauni, M.; Setälä, H.; Kotze, D.J. The invasive herb Lupinus polyphyllus can reduce plant species richness independently of local invasion age. Biol. Invasions 2021, 24, 425–436. [Google Scholar] [CrossRef]
- Ni, G.; Zhao, P.; Huang, Q.; Zhu, L.; Hou, Y.; Yu, Y.; Ye, Y.; Ouyang, L. Mikania micrantha invasion enhances the carbon (C) transfer from plant to soil and mediates the soil C utilization through altering microbial community. Sci. Total Environ. 2020, 711, 135020. [Google Scholar] [CrossRef] [PubMed]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Tatem, A.J. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fantle-Lepczyk, J.E.; Haubrock, P.J.; Kramer, A.M.; Cuthbert, R.N.; Turbelin, A.J.; Crystal-Ornelas, R.; Courchamp, F. Economic costs of biological invasions in the United States. Sci. Total Environ. 2022, 806, 151318. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarosik, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pysek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Courchamp, F.; Fournier, A.; Bellard, C.; Bertelsmeier, C.; Bonnaud, E.; Jeschke, J.M.; Russell, J.C. Invasion biology: Specific problems and possible solutions. Trends Ecol. Evol. 2017, 32, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shea, K.; Chesson, P. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Maron, J.L.; Vila, M. When do herbivores affect plant invasion? Evidence for the natural enemies and biotic resistance hypotheses. Oikos 2001, 95, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, A.D. Evolutionary significance of phenotypic plasticity in plants. Adv. Genet. 1965, 13, 115–155. [Google Scholar]
- Pigliucci, M.; Murren, C.J.; Schlichting, C.D. Phenotypic plasticity and evolution by genetic assimilation. J. Exp. Biol. 2006, 209, 2362–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, C.L.; Bossdorf, O.; Muth, N.Z.; Gurevitch, J.; Pigliucci, M. Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions. Ecol. Lett. 2006, 9, 981–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zenni, R.D.; Lamy, J.B.; Lamarque, L.J.; Porté, A.J. Adaptive evolution and phenotypic plasticity during naturalization and spread of invasive species: Implications for tree invasion biology. Biol. Invasions 2014, 16, 635–644. [Google Scholar] [CrossRef]
- Zheng, Y.; Burns, J.H.; Wang, R.; Yang, A.; Feng, Y. Identity recognition and the invasion of exotic plant. Flora 2021, 280, 151828. [Google Scholar] [CrossRef]
- Wang, M.; Ruan, W.; Kostenko, O.; Carvalho, S.; Hannula, S.E.; Mulder, P.P.; Bezemer, T.M. Removal of soil biota alters soil feedback effects on plant growth and defense chemistry. New. Phytol. 2019, 221, 1478–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, A.M.; Jennions, M.; Nicotra, A.B. Do invasive species show higher phenotypic plasticity than native species and if so, is it adaptive? A meta-analysis. Ecol. Lett. 2011, 14, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Kaur, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Phenotypic variations alter the ecological impact of invasive alien species: Lessons from Parthenium hysterophorus. J. Environ. Manage. 2019, 241, 187–197. [Google Scholar] [CrossRef]
- Hiatt, D.; Flory, S.L. Populations of a widespread invader and co-occurring native species vary in phenotypic plasticity. New. Phytol. 2020, 225, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Kleine, S.; Weissinger, L.; Müller, C. Impact of drought on plant populations of native and invasive origins. Oecologia 2017, 183, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Chen, Y.; Schaefer, V.; Liang, H.; Li, W.; Huang, S.; Peng, C. Responses of the hybrid between Sphagneticola trilobata and Sphagneticola calendulacea to low temperature and weak light characteristic in South China. Sci. Rep. 2015, 5, 16906. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Li, C.; Peng, S. Elevated CO2 increases energy-use efficiency of invasive Wedelia trilobata over its indigenous congener. Biol. Invasions 2010, 12, 1221–1230. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; van Kleunen, M. Increases and fluctuations in nutrient availability do not promote dominance of alien plants in synthetic communities of common natives. Funct. Ecol. 2018, 32, 2594–2604. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Oduor, A.M.; Zhang, Z.; Manea, A.; Tooth, I.M.; Leishman, M.R.; Van Kleunen, M. Do invasive alien plants benefit more from global environmental change than native plants? Global Change Biol. 2017, 23, 3363–3370. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liu, M.; Xu, X.; Tian, Y.; Zhang, Z.; van Kleunen, M. The effects of changes in water and nitrogen availability on alien plant invasion into a stand of a native grassland species. Oecologia 2018, 188, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Parendes, L.A.; Jones, J.A. Role of light availability and dispersal in exotic plant invasion along roads and streams in the HJ Andrews experimental forest. Oregon. Conserv. Biol. 2000, 14, 64–75. [Google Scholar] [CrossRef]
- Lambers, H.; Oliveira, R.S. Plant water relations. In Plant Physiological Ecology; Lambers, H., Oliveira, R.S., Eds.; Springer: Cham, Switzerland, 2019; pp. 187–263. [Google Scholar]
- Lozano, Y.M.; Aguilar-Trigueros, C.A.; Flaig, I.C.; Rillig, M.C. Root trait responses to drought are more heterogeneous than leaf trait responses. Funct. Ecol. 2020, 34, 2224–2235. [Google Scholar] [CrossRef]
- Valladares, F.; Sanchezgomez, D.; Zavala, M.A. Quantitative estimation of phenotypic plasticity: Bridging the gap between the evolutionary concept and its ecological applications. J. Ecol. 2006, 94, 1103–1116. [Google Scholar] [CrossRef]
- Gruntman, M.; Segev, U.; Tielbrger, K. Shade-induced plasticity in invasive Impatiens glandulifera populations. Weed Res. 2020, 60, 16–25. [Google Scholar] [CrossRef]
- Ma, J. The Checklist of the Chinese Invasive Plants; Higher Education Press: Beijing, China, 2013; pp. 1–324. (In Chinese) [Google Scholar]
- Liu, Y.; Oduor, A.M.; Dai, Z.; Gao, F.; Li, J.; Zhang, X.; Yu, F. Suppression of a plant hormone gibberellin reduces growth of invasive plants more than native plants. Oikos 2021, 130, 781–789. [Google Scholar] [CrossRef]
- Speißer, B.; Liu, Y.; van Kleunen, M. Biomass responses of widely and less-widely naturalized alien plants to artificial light at night. J. Ecol. 2021, 109, 1819–1827. [Google Scholar] [CrossRef]
- Valladares, F.; Wright, S.J.; Lasso, E.; Kitajima, K.; Pearcy, R.W. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest. Ecology 2000, 81, 1925–1936. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. Nlme: Linear and Nonlinear Mixed Effects Models; R Package Version 3.1-148; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Aerts, R.; Chapin, F. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. In Advances in Ecological Research; Academic Press: Cambridge, MA, USA, 1999; Volume 30, pp. 1–67. [Google Scholar]
- Koç, İ.; Nzokou, P.; Cregg, B. Biomass allocation and nutrient use efficiency in response to water stress: Insight from experimental manipulation of balsam fir, concolor fir and white pine transplants. New Forests 2022, 53, 915–933. [Google Scholar] [CrossRef]
- Kramer-Walter, K.R.; Laughlin, D.C. Root nutrient concentration and biomass allocation are more plastic than morphological traits in response to nutrient limitation. Plant Soil 2017, 416, 539–550. [Google Scholar] [CrossRef]
- Kleyer, M.; Trinogga, J.; Cebrian-Piqueras, M.A.; Trenkamp, A.; Flojgaard, C.; Ejrnaes, R.; Bouma, T.J.; Minden, V.; Maier, M.; Mantilla-Contreras, J.; et al. Trait correlation network analysis identifies biomass allocation traits and stem specific length as hub traits in herbaceous perennial plants. J. Ecol. 2019, 107, 829–842. [Google Scholar] [CrossRef] [Green Version]
- Weiner, J. Allocation, plasticity and allometry in plants. Perspect. Plant Ecol. Evol. Syst. 2004, 6, 207–215. [Google Scholar] [CrossRef]
- Rehling, F.; Sandner, T.M.; Matthies, D. Biomass partitioning in response to intraspecific competition depends on nutrients and species characteristics: A study of 43 plant species. J. Ecol. 2021, 109, 2219–2233. [Google Scholar] [CrossRef]
- Luong, J.C.; Loik, M.E. Adjustments in physiological and morphological traits suggest drought-induced competitive release of some California plants. Ecol. Evol. 2022, 12, e8773. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J.; Chapin, F.S.; Mooney, H.A. Resource limitation in plants—An economic analogy. Annu. Rev. Ecol. Syst. 1985, 363–392. [Google Scholar] [CrossRef]
- Tsukaya, H. A consideration of leaf shape evolution in the context of the primary function of the leaf as a photosynthetic organ. In The Leaf: A Platform for Performing Photosynthesis; Adams, W.W., III, Terashima, I., Eds.; Springer: Cham, Switzerland, 2018; Volume 44, pp. 1–26. [Google Scholar]
- Quan, G.; Mao, D.; Zhang, J.; Xie, J. Effects of nutrient level on plant growth and biomass allocation of invasive Chromolaena odorata. Ecol. Sci. 2015, 34, 27–33. [Google Scholar]
- Martinez, K.A.; Fridley, J.D. Acclimation of leaf traits in seasonal light environments: Are non-native species more plastic? J. Ecol. 2018, 106, 2019–2030. [Google Scholar] [CrossRef]
- Bazzaz, F.A.; Chiarirllo, N.R.; Coley, P.D.; Pitelak, L.F. Allocating resources to reproduction and defense. BioScience 1987, 37, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Harper, J.L.; Ogden, J. The reproductive strategy of higher plants: I. The concept of strategy with special reference to Senecio Vulgaris, L. J. Ecol. 1970, 58, 681–698. [Google Scholar] [CrossRef]
- Eziz, A.; Yan, Z.; Tian, D.; Han, W.; Tang, Z.; Fang, J. Drought effect on plant biomass allocation: A meta-analysis. Ecol. Evol. 2017, 7, 11002–11010. [Google Scholar] [CrossRef]
- Valliere, J.M.; Escobedo, E.B.; Bucciarelli, G.M.; Sharifi, M.R.; Rundel, P.W. Invasive annuals respond more negatively to drought than native species. New. Phytol. 2019, 223, 1647–1656. [Google Scholar] [CrossRef]
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating resources in plant communities: A general theory of invasibility. J. Ecol. 2000, 88, 528–534. [Google Scholar] [CrossRef] [Green Version]
- Goergen, E.; Daehler, C.C. Reproductive ecology of a native Hawaiian grass (Heteropogon contortus; Poaceae) versus its invasive alien competitor (Pennisetum setaceum; Poaceae). Int. J. Plant Sci. 2001, 162, 317–326. [Google Scholar] [CrossRef]
- Muth, N.Z.; Pigliucci, M. Implementation of a novel framework for assessing species plasticity in biological invasions: Responses of Centaurea and Crepis to phosphorus and water availability. J. Ecol. 2007, 95, 1001–1013. [Google Scholar] [CrossRef]
- Sorte, C.J.; Ibáñez, I.; Blumenthal, D.M.; Molinari, N.A.; Miller, L.P.; Grosholz, E.D.; Dukes, J.S. Poised to prosper? A cross-system comparison of climate change effects on native and non-native species performance. Ecol. Lett. 2013, 16, 261–270. [Google Scholar] [CrossRef]
- Huang, W.; Zhang, S.B.; Liu, T. Moderate photoinhibition of photosystem II significantly affects linear electron flow in the shade-demanding plant Panax notoginseng. Front. Plant Sci. 2018, 9, 637. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Iqbal, N.; Ting, P.; Khan, M.N.; Liu, W.; Yang, W. Weak stem under shade reveals the lignin reduction behavior. J. Integr. Agr. 2019, 18, 496–505. [Google Scholar] [CrossRef]
- Liu, W.; Ren, M.; Liu, T.; Du, Y.; Zhou, T.; Liu, X.; Liu, J.; Hussain, S.; Yang, W. Effect of shade stress on lignin biosynthesis in soybean stems. J. Integr. Agr. 2018, 17, 1594–1604. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, U.; Ntagkas, N.; Siebenkas, A.; Maenpaa, M.; Matsubara, S.; Pons, T. A meta-analysis of plant responses to light intensity for 70 traits ranging from molecules to whole plant performance. New. Phytol. 2019, 223, 1073–1105. [Google Scholar] [CrossRef] [Green Version]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Villar, R. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Dawson, W.; Prati, D.; Haeuser, E.; Feng, Y.; van Kleunen, M. Does greater specific leaf area plasticity help plants to maintain a high performance when shaded? Ann. Bot. London 2016, 118, 1329–1336. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Sun, B.; Li, J.; Wang, G.; Sun, J.; Wang, X.; Zhong, R. Effects of light intensity on the phenotypic plasticity of invasive species Ambrosia trifida. J. Appl. Ecol. 2012, 23, 1797–1802. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Origin of the Plants |
|---|---|---|
| Sphagneticola trilobata | Asteraceae | Invasive |
| Alternanthera philoxeroides | Amaranthaceae | Invasive |
| Hydrocotyle vulgaris | Araliaceae | Invasive |
| Sphagneticola calendulacea | Asteraceae | Native |
| Alternanthera sessilis | Amaranthaceae | Native |
| Hydrocotyle sibthorpioides | Araliaceae | Native |
| Fixed Effects | Height | Leaf Number | SLA | Total Biomass | RMF | LMF |
|---|---|---|---|---|---|---|
| Nutrient | ||||||
| Initial Height | 7.46 ** | 6.38 * | 0.11 | 17.94 *** | 15.17 *** | 3.29 |
| NT | 6.59 ** | 1.69 | 0.01 | 22.86 *** | 39.88 *** | 4.50 * |
| Origin | 2.27 | 0.48 | 2.77 | 1.66 | 0.43 | 0.84 |
| NT × Origin | 3.55 | 3.32 | 0.18 | 0.03 | 1.01 | 11.50 *** |
| Random Effects | SD | SD | SD | SD | SD | SD |
| Genus | 0.325 | 8.531 | 1.585 | 0.251 | 0.156 | 4.443 |
| Species | 0.333 | 1.116 | 0.210 | 0.655 | 0.136 | 0.298 |
| R2 | ||||||
| Marginal | 0.182 | 0.119 | 0.288 | 0.238 | 0.267 | 0.151 |
| Conditional | 0.540 | 0.583 | 0.740 | 0.521 | 0.496 | 0.753 |
| Water | ||||||
| Initial Height | 0.01 | 0.06 | 3.85 * | 4.17 * | 5.60 * | 12.49 *** |
| WT | 22.55 *** | 227.17 *** | 12.78 *** | 43.83 *** | 1.41 | 0.92 |
| Origin | 1.77 | 0.65 | 5.48 * | 6.13 * | 0.30 | 0.06 |
| WT × Origin | 0.36 | 10.64 *** | 0.15 | 4.52 * | 0.35 | 0.03 |
| Random Effects | SD | SD | SD | SD | SD | SD |
| Genus | 0.201 | 0.296 | 1.084 | 0.980 | 0.227 | 0.135 |
| Species | 0.112 | 0.495 | 0.132 | 0.273 | 0.274 | 0.185 |
| R2 | ||||||
| Marginal | 0.173 | 0.129 | 0.551 | 0.451 | 0.071 | 0.095 |
| Conditional | 0.555 | 0.734 | 0.819 | 0.788 | 0.908 | 0.601 |
| Light | ||||||
| Initial Height | 0.88 | 1.16 | 0.65 | 5.49 * | 0.01 | 0.83 |
| LT | 5.78 * | 85.57 *** | 91.60 *** | 153.32 *** | 3.85 * | 2.94 |
| Origin | 4.09 * | 0.32 | 3.42 | 1.42 | 1.27 | 0.78 |
| LT × Origin | 0.05 | 0.01 | 4.67 * | 3.18 | 3.86 * | 0.09 |
| Random Effects | SD | SD | SD | SD | SD | SD |
| Genus | 0.004 | 0.736 | 1.273 | 0.001 | 0.251 | 3.392 |
| Species | 14.760 | 0.126 | 0.197 | 1.062 | 0.193 | 11.054 |
| R2 | ||||||
| Marginal | 0.603 | 0.573 | 0.558 | 0.574 | 0.092 | 0.112 |
| Conditional | 0.781 | 0.872 | 0.817 | 0.806 | 0.390 | 0.891 |
| Treatment | Variables | Invasive | Native |
|---|---|---|---|
| Nutrient | Final Height | 0.189 ± 0.14a | 0.372 ± 0.12a |
| Leaf Number | 0.314 ± 0.12a | 0.597 ± 0.15a | |
| SLA | 0.049 ± 0.02b | 0.148 ± 0.09a | |
| Total Biomass | 0.364 ± 0.16a | 0.674 ± 0.10a | |
| RMF | 0.485 ± 0.01a | 0.387 ± 0.06b | |
| LMF | 0.165 ± 0.01a | 0.109 ± 0.10b | |
| Water | Final Height | 0.132 ± 0.05a | 0.222 ± 0.02a |
| Leaf Number | 0.334 ± 0.04a | 0.396 ± 0.05a | |
| SLA | 0.105 ± 0.03a | 0.072 ± 0.03a | |
| Total Biomass | 0.264 ± 0.05a | 0.312 ± 0.06a | |
| RMF | 0.098 ± 0.05b | 0.172 ± 0.01a | |
| LMF | 0.074 ± 0.04a | 0.077 ± 0.04a | |
| Light | Final Height | 0.369 ± 0.13a | 0.480 ± 0.08a |
| Leaf Number | 0.795 ± 0.10a | 0.826 ± 0.03a | |
| SLA | 0.342 ± 0.01a | 0.351 ± 0.08a | |
| Total Biomass | 0.900 ± 0.04a | 0.876 ± 0.04a | |
| RMF | 0.233 ± 0.05a | 0.252 ± 0.07a | |
| LMF | 0.061 ± 0.03a | 0.172 ± 0.08a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Chen, A.; Li, Y.; Li, D.; Cheng, S.; Cheng, L.; Liu, Y. Differences in Phenotypic Plasticity between Invasive and Native Plants Responding to Three Environmental Factors. Life 2022, 12, 1970. https://doi.org/10.3390/life12121970
Zhang L, Chen A, Li Y, Li D, Cheng S, Cheng L, Liu Y. Differences in Phenotypic Plasticity between Invasive and Native Plants Responding to Three Environmental Factors. Life. 2022; 12(12):1970. https://doi.org/10.3390/life12121970
Chicago/Turabian StyleZhang, Luna, Anqun Chen, Yanjiao Li, Duohui Li, Shiping Cheng, Liping Cheng, and Yinzhan Liu. 2022. "Differences in Phenotypic Plasticity between Invasive and Native Plants Responding to Three Environmental Factors" Life 12, no. 12: 1970. https://doi.org/10.3390/life12121970