

Prokaryotic Diversity of Ruminal Content and Its Relationship with Methane Emissions in Cattle from Kazakhstan
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Average Daily Rations
2.3. Methane Measurement
2.4. DNA Extraction, PCR Amplification, and 16S Metabarcoding
2.5. Bioinformatics and Data Analysis
3. Results
3.1. Dynamics of Live Weight of Animals
3.2. Methane Emissions from Animals
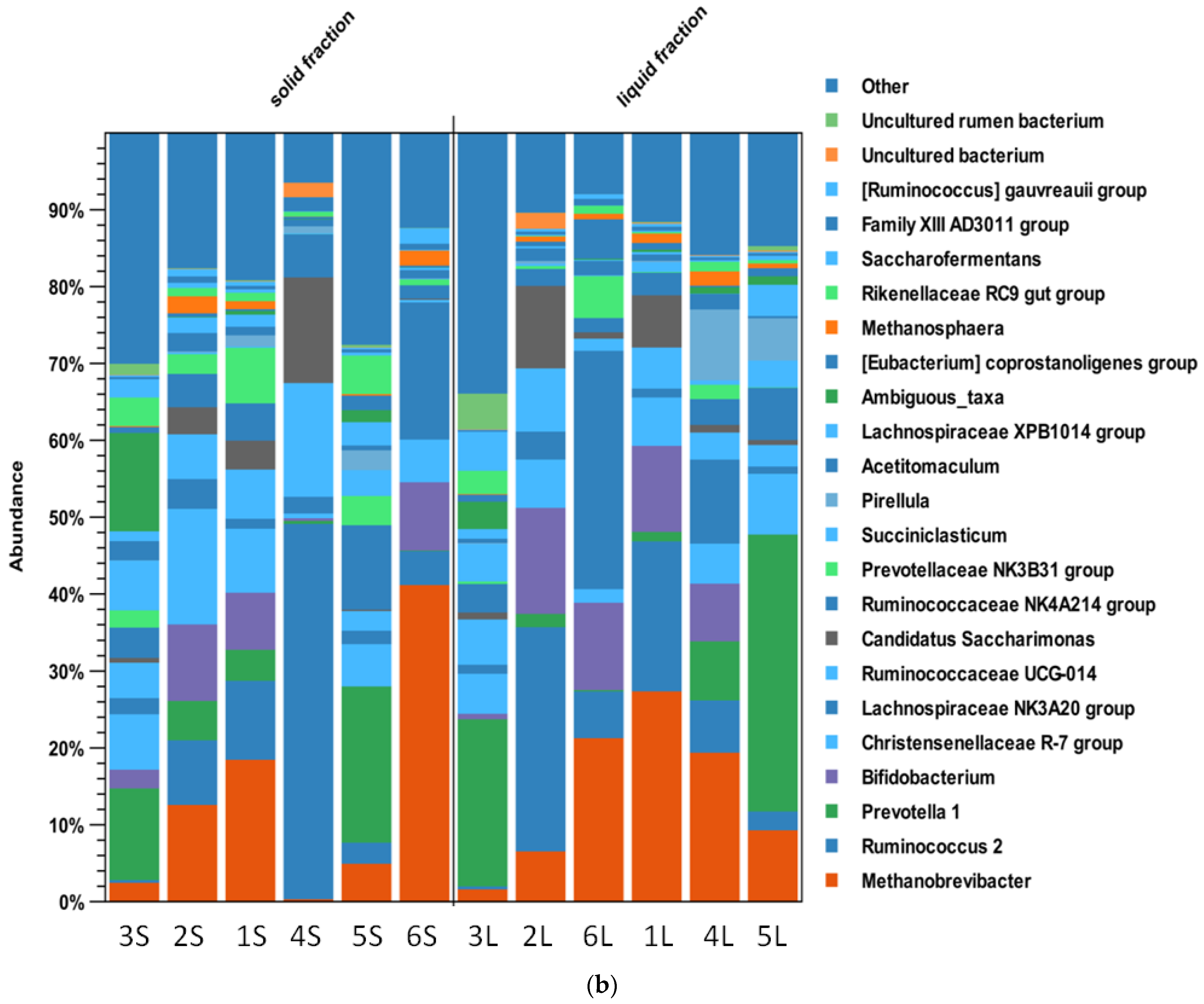
3.3. Rumen Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, M.; Park, T.; Yu, Z. Metagenomic investigation of gastrointestinal microbiome in cattle. Asian Australas. J. Anim. Sci. 2017, 30, 1515–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, R.J.; Sasson, G.; Garnsworthy, P.C.; Tapio, I.; Gregson, E.; Bani, P.; Huhtanen, P.; Bayat, A.R.; Strozzi, F.; Biscarini, F.; et al. A heritable subset of the core rumen microbiome dictates dairy cow productivity and emissions. Sci. Adv. 2019, 5, eaav8391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinfeld, H.; Gerber, P.; Wassenaar, T.; Caste, V.; Rosales, M.; de Haan, C. Livestock’s Long Shadow; FAO: Rome, Italy, 2006. [Google Scholar]
- Fouts, D.E.; Szpakowski, S.; Purushe, J.; Torralba, M.; Waterman, R.C.; MacNeil, M.D.; Alexander, L.J.; Nelson, K.E. Next Generation Sequencing to Define Prokaryotic and Fungal Diversity in the Bovine Rumen. PLoS ONE 2012, 7, e48289. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The rumen microbiome: A crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes 2019, 10, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef]
- Johnson, D.E.; Ward, G.M. Estimates of animal methane emissions. Environ. Monit. Assess. 1996, 42, 133–141. [Google Scholar] [CrossRef]
- Ishler, V.; Heinrichs, A.J.; Varga, G. From Feed to Milk: Understanding Rumen Function; Pennsylvania State University Extension Circular: University Park, PA, USA, 1996; Volume 422. [Google Scholar]
- Brulc, J.M.; Antonopoulos, D.A.; Miller, M.E.B.; Wilson, M.K.; Yannarell, A.C.; Dinsdale, E.A.; Edwards, R.E.; Frank, E.D.; Emerson, J.B.; Wacklin, P.; et al. Gene-centric metagenomics of the fiber-adherent bovine rumen microbiome reveals forage specific glycoside hydrolases. Proc. Natl. Acad. Sci. USA 2009, 106, 1948–1953. [Google Scholar] [CrossRef] [Green Version]
- Hungate, R.E.; Bryant, M.P.; Mah, R.A. The rumen bacteria and protozoa. Annu. Rev. Microbiol. 1964, 18, 131–166. [Google Scholar] [CrossRef]
- Khafipour, E.; Li, S.; Plaizier, J.C.; Krause, D.O. Rumen Microbiome Composition Determined Using Two Nutritional Models of Subacute Ruminal Acidosis. Appl. Environ. Microbiol. 2009, 75, 7115–7124. [Google Scholar] [CrossRef] [Green Version]
- Hook, S.E.; Wright, A.-D.G.; McBride, B.W. Methanogens: Methane producers of the rumen and mitigation strategies. Archaea 2010, 2010, 945785. [Google Scholar] [CrossRef]
- Lin, C.; Raskin, L.; Stahl, D.A. Microbial community structure in gastrointestinal tracts of domestic animals: Comparative analyses using rRNA-targeted oligonucleotide probes. FEMS Microbiol. Ecol. 1997, 22, 281–294. [Google Scholar] [CrossRef]
- Wallace, R.J.; Rooke, J.A.; McKain, N.; Duthie, C.-A.; Hyslop, J.J.; Ross, D.W.; Waterhouse, A.; Watson, M.; Roehe, R. The rumen microbial metagenome associated with high methane production in cattle. BMC Genom. 2015, 16, 839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deusch, S.; Camarinha-Silva, A.; Conrad, J.; Beifuss, U.; Rodehutscord, M.; Seifert, J. A structural and functional elucidation of the rumen microbiome influenced by various diets and microenvironments. Front. Microbiol. 2017, 8, 1605. [Google Scholar] [CrossRef] [Green Version]
- Vaidya, J.D.; van den Bogert, B.; Edwards, J.E.; Boekhorst, J.; van Gastelen, S.; Saccenti, E.; Plugge, C.M.; Smidt, H. The effect of DNA extraction methods on observed microbial communities from fibrous and liquid rumen fractions of dairy cows. Front. Microbiol. 2018, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Welkie, D.G.; Stevenson, D.M.; Weimer, P.J. ARISA analysis of ruminal bacterial community dynamics in lactating dairy cows during the feeding cycle. Anaerobe 2009, 16, 94–100. [Google Scholar] [CrossRef]
- Jami, E.; Mizrahi, I. Composition and similarity of bovine rumen microbiota across individual animals. PLoS ONE 2012, 7, e33306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Hernandez-Sanabria, E.; Guan, L.L. Characterization of variation in rumen methanogenic communities under different dietary and host feed efficiency conditions, as determined by PCR-denaturing gradient gel electro-phoresis analysis. Appl. Environ. Microbiol. 2010, 76, 3776–3786. [Google Scholar] [CrossRef] [Green Version]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W. Global Rumen Census Collaborators and Janssen PH Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef] [Green Version]
- Mizrahi Jami, E. Review: The compositional variation of the rumen microbiome and its effect on host performance and methane emission. Animal 2018, 12, 220–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daugaliyeva, A.; Daugaliyeva, S.; Ashanin, A.; Kanatbayev, S.; Beltramo, C.; Peletto, S. Study of cattle microbiota in different regions of Kazakhstan using 16S metabarcoding analysis. Sci. Rep. 2022, 12, 16410. [Google Scholar] [CrossRef]
- Bach, A.; López-García, A.; González-Recio, O.; Elcoso, G.; Fàbregas, F.; Chaucheyras-Durand, F.; Castex, M. Changes in the rumen and colon microbiota and effects of live yeast dietary supplementation during the transition from the dry period to lactation of dairy cows. J. Dairy Sci. 2019, 102, 6180–6198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Guan, L.L. Metatranscriptomic profiling reveals linkages between the active rumen microbiome and feed efficiency in beef cattle. Appl. Environ. Microbiol. 2017, 83, e00061-e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myer, P.R.; Smith, T.P.; Wells, J.E.; Kuehn, L.A.; Freetly, H.C. Rumen microbiome from steers differing in feed efficiency. PLoS ONE 2015, 10, e0129174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freetly, H.C.; Dickey, A.; Lindholm-Perry, A.K.; Thallman, R.M.; Keele, J.W.; Foote, A.P.; Wells, J.E. Digestive tract microbiota of beef cattle that differed in feed efficiency. J. Anim. Sci. 2020, 98. [Google Scholar] [CrossRef] [Green Version]
- Gagen, E.J.; Padmanabha, J.; Denman, S.E.; McSweeney, C.S. Hydrogenotrophic culture enrichment reveals rumen Lachnospiraceae and Ruminococcaceae acetogens and hydrogen-responsive Bacteroidetes from pasturefed cattle. FEMS Microbiol. Lett. 2015, 362, fnv104. [Google Scholar] [CrossRef] [Green Version]
- Khafipour, E.; Li, S.; Tun, H.M.; Derakhshani, H.; Moossavi, S.; Plaizier, J.C. Effects of grain feeding on microbiota in the digestive tract of cattle. Anim. Front. 2016, 6, 13–19. [Google Scholar] [CrossRef]
- Liu, J.; Liu, F.; Cai, W.; Jia, C.; Bai, Y.; He, Y.; Zhu, W.; Li, R.W.; Song, J. Diet-induced Changes in Bacterial Communities in the Jejunum and Their Associations with Bile Acids in Angus Beef Cattle. Anim. Microbiome 2020, 2, 33. [Google Scholar] [CrossRef] [PubMed]
- Kelly, W.J.; Leahy, S.C.; Kamke, J.; Soni, P.; Koike, S.; Mackie, R.; Seshadri, R.; Cook, G.M.; Morales, S.E.; Greening, C.; et al. Occurrence and expression of genes encoding methyl-compound production in rumen bacteria. Anim. Microbiome. 2019, 1, 15. [Google Scholar] [CrossRef] [PubMed]
- Delgado, B.; Bach, A.; Guasch, I.; González, C.; Elcoso, G.; Pryce, J.E.; Gonzalez-Recio, O. Whole rumen metagenome sequencing allows classifying and predicting feed efficiency and intake levels in cattle. Sci. Rep. 2019, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Fernandez, G.; Denman, S.E.; Yang, C.; Cheung, J.; Mitsumori, M.; McSweeney, C.S. Methane Inhibition Alters the Microbial Community, Hydrogen Flow, and Fermentation Response in the Rumen of Cattle. Front. Microbiol. 2016, 7, 1122. [Google Scholar] [CrossRef]
- Zhang, G.; Wang, Y.; Luo, H.; Qiu, W.; Zhang, H.; Hu, L.; Wang, Y.; Dong, G.; Guo, G. The Association Between Inflammaging and Age-Related Changes in the Ruminal and Fecal Microbiota Among Lactating Holstein Cows. Front. Microbiol. 2019, 10, 1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghavendra, B.; Osamu, E.; Mitsunori, K. Measurement of Methane Production from Ruminants. Asian Aust. J. Anim. Sci. 2007, 20, 1305–1318. [Google Scholar]
- Brouček, J. Methods of methane measurement in ruminants. Slovak J. Anim. Sci. 2014, 47, 51–60. [Google Scholar]
- Garnsworthy, P.C.; Difford, G.F.; Bell, M.J.; Bayat, A.R.; Huhtanen, P.; Kuhla, B.; Lassen, J.; Peiren, N.; Pszczola, M.; Sorg, D.; et al. Comparison of Methods to Measure Methane for Use in Genetic Evaluation of Dairy Cattle. Animals 2019, 9, 837. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Quantity |
|---|---|
| Alfalfa hay, kg | 7.81 |
| Straw, kg | 1.64 |
| Concentrates, kg | 5 |
| The diet contains | |
| Energy feed units, Mj | 11.9 |
| Exchange energy, Mj | 119.4 |
| Dry matter, kg | 12.6 |
| Crude protein, g | 1970.1 |
| Digestible protein, g | 1350.8 |
| Split protein, g | 1518.7 |
| Non-degradable protein, g | 451.4 |
| Crude fiber, g | 2701.6 |
| Crude fat, g | 271.4 |
| Sugar, g | 236.1 |
| Starch, g | 2887.1 |
| Calcium, g | 140.2 |
| Phosphorus, g | 33.7 |
| Carotene, mg | 390.9 |
| Iron, mg | 2360.8 |
| Zinc, g | 360.8 |
| Manganese, g | 500.0 |
| Copper, mg | 107.3 |
| Cobalt, mg | 2.9 |
| Iodine, mg | 4.3 |
| Sample Name | ID | Fraction | Animal Breed |
|---|---|---|---|
| 1L | SimJ | Liquid | Simmental |
| 1S | SimTv | Solid | Simmental |
| 2L | AlJ | Liquid | Alatau (local breed) |
| 2S | Al TB | Solid | Alatau (local breed) |
| 3L | 8 BelKJ | Liquid | Kazakh white-headed |
| 3S | 7 KBKTv | Solid | Kazakh white-headed |
| 4L | T-1 J | Liquid | Kazakh white-headed |
| 4S | T1-Tv | Solid | Kazakh white-headed |
| 5L | T2-J | Liquid | Alatau (local breed) |
| 5S | T2-Tv | Solid | Alatau (local breed) |
| 6L | GKrJ | Liquid | Holstein-Frisian |
| 6S | GKrTv | Solid | Holstein-Frisian |
| Indicators | Result |
|---|---|
| Average live weight of steers when setting up for experiment, kg | 394 ± 16.4 |
| Live weight of steers at the end of the experiment, kg | 470 ± 24.6 |
| Absolute increase in live weight, kg | 76.8 ± 8.2 |
| Average daily live weight gain, g | 1476 ± 115.4 |
| Total feed costs, energy feed unit | 619 ± 35.7 |
| Feed costs per 1 kg of live weight gain, EFU | 8.1 ±0.74 |
| Gas (Volumetric Share, %) | Steers | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |
| Methane (CH4) | 4.02 | 3.92 | 3.05 | 3.63 | 3.21 | 4.4 |
| Carbon dioxide (CO2) | 3.62 | 3.11 | 2.92 | 3.31 | 3.91 | 4.54 |
| Liquid Fraction vs. Solid Fraction | |||
|---|---|---|---|
| Family | Log₂ Fold Change | Fold Change | p-Value |
| Streptococcaceae | 3.14 | 8.79 | 0.005 |
| Lactobacillaceae | 6.35 | 81.33 | 0.010 |
| Desulfobulbaceae | 2.48 | 5.57 | 0.013 |
| Succinivibrionaceae | 3.64 | 12.47 | 0.021 |
| Thalassospiraceae | −4.23 | −18.72 | 0.035 |
| Genus | Log₂ Fold Change | Fold Change | p-Value |
| Ruminobacter | 5.13 | 34.95 | 0.002 |
| Howardella | 3.23 | 9.41 | 0.005 |
| Streptococcus | 3.02 | 8.12 | 0.006 |
| Lactobacillus | 6.71 | 104.97 | 0.007 |
| Ruminococcaceae UCG-013 | 2.68 | 6.40 | 0.016 |
| [Eubacterium] coprostanoligenes group | 2.02 | 4.06 | 0.026 |
| Thalassospira | −4.40 | −21.14 | 0.028 |
| Succinivibrionaceae UCG-002 | 3.31 | 9.91 | 0.033 |
| Desulfobulbus | 2.39 | 5.22 | 0.036 |
| Psychrobacter | 4.79 | 27.70 | 0.040 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daugaliyeva, A.; Daugaliyeva, S.; Ashanin, A.; Beltramo, C.; Mamyrova, L.; Yessembekova, Z.; Peletto, S. Prokaryotic Diversity of Ruminal Content and Its Relationship with Methane Emissions in Cattle from Kazakhstan. Life 2022, 12, 1911. https://doi.org/10.3390/life12111911
Daugaliyeva A, Daugaliyeva S, Ashanin A, Beltramo C, Mamyrova L, Yessembekova Z, Peletto S. Prokaryotic Diversity of Ruminal Content and Its Relationship with Methane Emissions in Cattle from Kazakhstan. Life. 2022; 12(11):1911. https://doi.org/10.3390/life12111911
Chicago/Turabian StyleDaugaliyeva, Aida, Saule Daugaliyeva, Alexander Ashanin, Chiara Beltramo, Latipa Mamyrova, Zinagul Yessembekova, and Simone Peletto. 2022. "Prokaryotic Diversity of Ruminal Content and Its Relationship with Methane Emissions in Cattle from Kazakhstan" Life 12, no. 11: 1911. https://doi.org/10.3390/life12111911