
Application of Solid-State Fermentation for the Improving of Extruded Corn Dry-Milling By-Products and Their Protein Functional Properties
 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Raw Material
2.2. Microorganisms
2.3. Experimental Design
2.4. Microbiological Analysis
2.5. Determination of Amino Acids Profile
2.6. Chemical Analyzes
2.7. Determination of Xylanase and Protease Activities
2.8. Protein Fractionation
2.9. Determination of Albumin and Globulin Functional Properties
2.9.1. Water-Absorption Capacity and Solubility
2.9.2. Emulsifying Capacity and Emulsion Stability
2.9.3. Foam-Forming Capacity and Foam Stability
2.10. In Vitro Protein Digestibility
2.11. Degree of Hydrolysis
2.12. Total Phenolic Content
2.13. Determination of Antioxidant Activity
2.14. Statistical Analysis
3. Results and Discussion
3.1. Characterization of Stabilized CMB Material
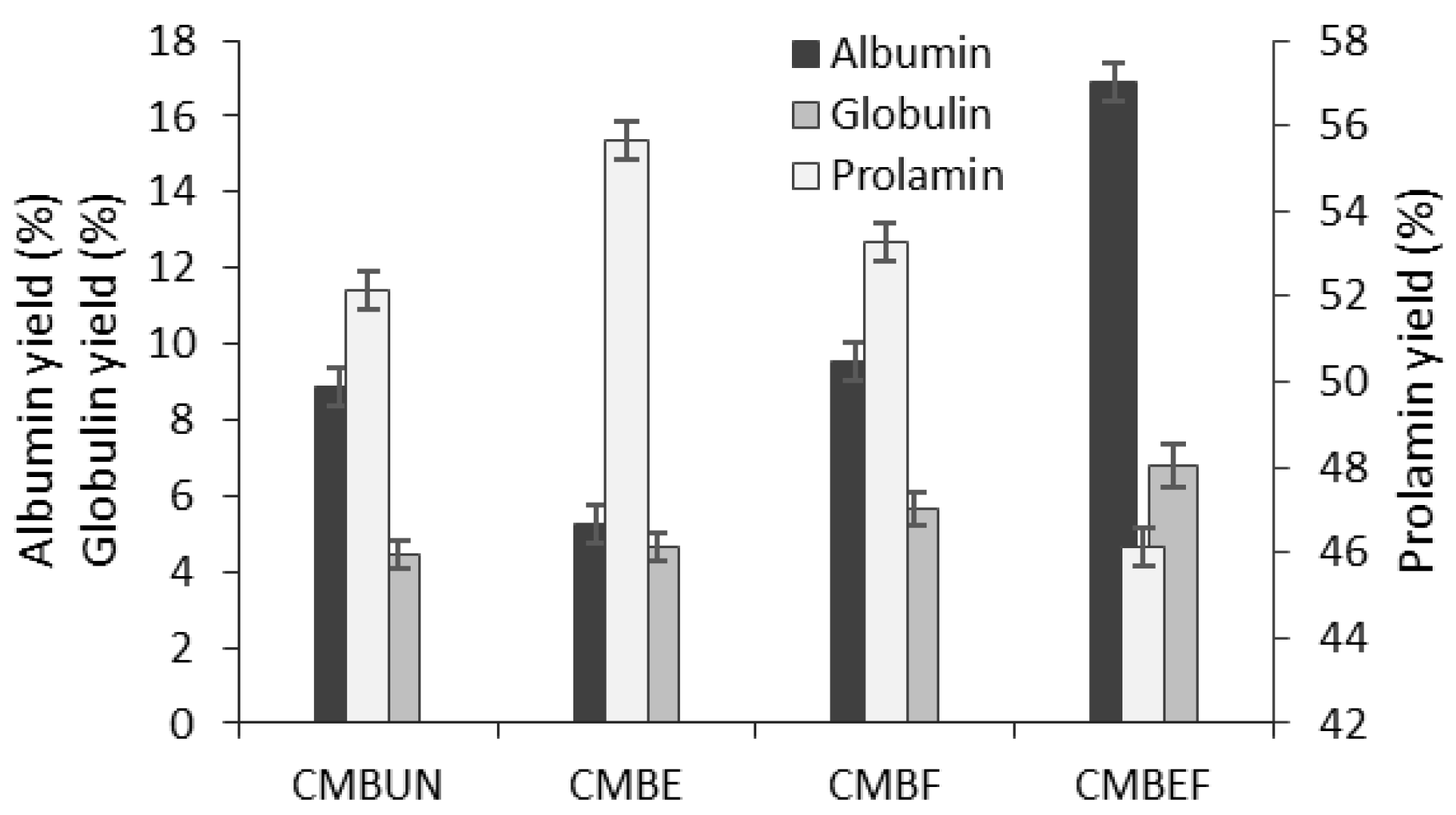
3.2. The Influence of the Extrusion and Solid-State Fermentation on Protein Yield and the Retention of Amino Acids in CMB
3.2.1. The Effect of Extrusion on the Protein Extraction Yields and the Amino Acid Profile
3.2.2. The Effect of SSF on the Improvement of Protein Yields and Amino Acid Profile
3.3. The Influence of Extrusion and SSF on CMB Albumin and Globulin Functional Properties
3.3.1. Solubility and Water-Absorption Capacity
3.3.2. Foaming Capacity and Foam Stability
3.3.3. Emulsifying Capacity and Emulsion Stability
3.3.4. The Influence of SSF on the Changes of Extruded Albumins and Globulins Functional Properties
3.4. The Effect of Extrusion and SSF on the Functional Properties and Bioactivity of CMB Prolamins
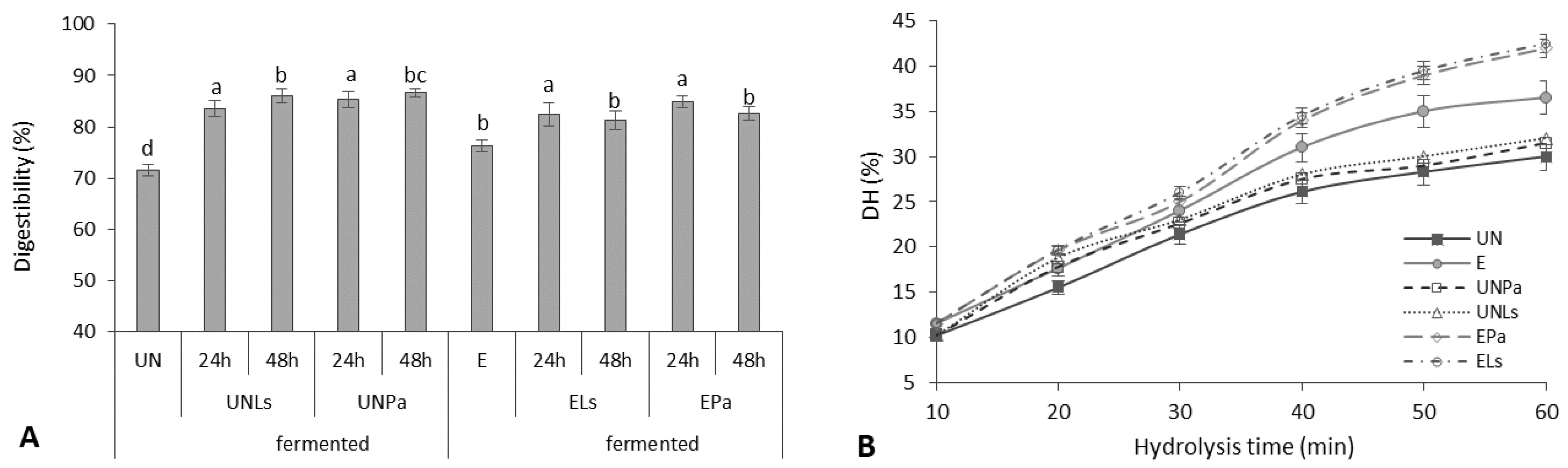
3.4.1. Digestibility and Degree of Hydrolysis
3.4.2. Antioxidant Activity of Prolamins
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Revilla, P.; Alves, M.L.; Andelković, V.; Balconi, C.; Dinis, I.; Mendes-Moreira, P.; Redaelli, R.; Ruiz de Galarreta, J.I.; Vaz Patto, M.C.; Žilić, S.; et al. Traditional foods from maize (Zea mays L.) in Europe. Front. Nutr. 2022, 8, 683399. [Google Scholar] [CrossRef] [PubMed]
- Deepak, T.S.; Jayadeep, P.A. Prospects of maize (corn) wet milling by-products as a source of functional food ingredients and nutraceuticals. Food Technol. Biotechnol. 2022, 60, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Saeed, F.; Niaz, B.; Afzaal, M.; Ikram, A.; Hussain, S.; Mohamed, A.A.; Alamri, M.S.; Anjum, F.M. Biochemical and nutritional profile of maize bran-enriched flour in relation to its end-use quality. Food Sci. Nutr. 2021, 9, 3336–3345. [Google Scholar] [CrossRef] [PubMed]
- Loy, D.D.; Lundy, E.L. Nutritional properties and feeding value of corn and its coproducts. In Corn: Chemistry and Technology; Serna-Saldivar, S.O., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 633–659. [Google Scholar]
- Trinidad-Calderón, P.A.; Acosta-Cruz, E.; Rivero-Masante, M.N.; Díaz-Gómez, J.L.; García-Lara, S.; López-Castillo, L.M. Maize bioactive peptides: From structure to human health. J. Cereal. Sci. 2021, 100, 103232. [Google Scholar] [CrossRef]
- Li, G.; Liu, W.; Wang, Y.; Jia, F.; Wang, Y.; Ma, Y.; Gu, R.; Lu, J. Chapter one—Functions and applications of bioactive peptides from corn gluten meal. Adv. Food Nutr. Res. 2019, 87, 1–41. [Google Scholar]
- Juodeikiene, G.; Zadeike, D.; Vidziunaite, I.; Bartkiene, E.; Bartkevics, V.; Pugajeva, I. Effect of heating method on the microbial levels and acrylamide in corn grits and subsequent use as functional ingredient for bread making. Food Bioprod. Process. 2018, 112, 22–30. [Google Scholar] [CrossRef]
- Rodríguez-España, M.; Figueroa-Hernández, C.Y.; de Dios Figueroa-Cárdenas, J.; Rayas-Duarte, P.; Josué Hernández-Estrada, Z. Effects of germination and lactic acid fermentation on nutritional and rheological properties of sorghum: A graphical review. Curr. Res. Food Sci. 2022, 5, 807–812. [Google Scholar] [CrossRef]
- Velikova, P.; Stoyanov, A.; Blagoeva, G.; Popova, L.; Petrov, K.; Gotcheva, V.; Angelov, A.; Petrova, P. Starch utilization routes in lactic acid bacteria: New insight by gene expression assay. Starch-Stärke 2016, 68, 953–960. [Google Scholar] [CrossRef]
- Yang, S.; Zheng, M.; Cao, Y.; Dong, Y.; Yaqoob, S.; Liu, J. Optimization of liquid fermentation conditions for biotransformation zein by Cordyceps militaris 202 and characterization of physicochemical and functional properties of fermentative hydrolysates. Braz. J. Microbiol. 2018, 49, 621–631. [Google Scholar] [CrossRef]
- Hegazy, H.S.; El-Bedawey, A.E.-F.A.; El-Sayed, H.R.; Gaafar, A.M. Effect of extrusion processs on nutritional, functional properties and antioxidant activity of germinated chickpea incorporated corn extrudates. Am. J. Food Sci. Nutr. Res. 2017, 4, 59–66. [Google Scholar]
- Wang, X.; Jin, S.; Gou, C.; Hu, L.; Zhang, J.; Li, F.; Zhai, D.; Zhao, Y.; Jihong Huang, J.; Hui, M. Extraction optimization and functional properties of corn germ meal albumin protein as a potential source of novel food ingredients. J. Food Process. Preserv. 2022, 46, e16218. [Google Scholar] [CrossRef]
- Digaitiene, A.; Hansen, A.S.; Juodeikiene, G.; Eidukonyte, D.; Josephsen, J. Lactic acid bacteria isolated from rye sourdoughs produce bacteriocin-like inhibitory substances active against Bacillus subtilis and fungi. J. Appl. Microbiol. 2012, 112, 732–742. [Google Scholar] [CrossRef]
- Jukonyte, R.; Zadeike, D.; Bartkiene, E.; Lele, V.; Juodeikiene, G.; Cernauskas, D.; Suproniene, S. A potential of brown rice polish as a substrate for the lactic acid and bioactive compounds production by the lactic acid bacteria newly isolated from cereal-based fermented products. LWT Food Sci. Technol. 2018, 97, 323–331. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 17th ed.; Approved Methods of Analysis 920.152, 978.10, 942.05, 996.01; The Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2018. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Malumba, P.; Vanderghem, C.; Deroanne, C.; Béra, F. Influence of drying temperature on the solubility, the purity of isolates and the electrophoretic patterns of corn proteins. Food Chem. 2008, 111, 564–572. [Google Scholar] [CrossRef]
- Silva-Sanchez, C.; Gonzalez-Castaneda, J.; de Leon-Rodriguez, A.; De La Rosa, B.A.P. Functional and rheological properties of amaranth albumins extracted from two Mexican varieties. Plant Food Hum. Nutr. 2004, 59, 169–174. [Google Scholar] [CrossRef]
- Almeida, C.C.; Monteiro, M.L.G.; Costa-Lima, B.R.C.D.; Alvares, T.S.; Conte-Junior, C.A. In vitro digestibility of commercial whey protein supplements. LWT Food Sci. Technol. 2015, 61, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysate by trinitrobenzensulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphofungistic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Tang, N.; Zhuang, H. Evaluation of antioxidant activities of zein protein fractions. J. Food Sci. 2014, 79, C2174–C2184. [Google Scholar] [CrossRef]
- Moisio, T.; Forssell, P.; Partanen, R.; Damerau, A.; Hill, S.E. Reorganisation of starch, proteins and lipids in extrusion of oats. J. Cereal Sci. 2015, 64, 48–55. [Google Scholar] [CrossRef]
- Zhang, M.; Bai, X.; Zhang, Z. Extrusion process improves the functionality of soluble dietary fiber in oat bran. J. Cereal Sci. 2011, 54, 98–103. [Google Scholar] [CrossRef]
- Petry, A.L.; Huntley, N.F.; Bedford, M.R.; Patience, J.F. The influence of xylanase on the fermentability, digestibility, and physicochemical properties of insoluble corn-based fiber along the gastrointestinal tract of growing pigs. J. Anim. Sci. 2021, 99, 159. [Google Scholar] [CrossRef] [PubMed]
- Kamau, E.H.; Nkhata, S.G.; Ayua, E.O. Extrusion and nixtamalization conditions influence the magnitude of change in the nutrients and bioactive components of cereals and legumes. Food Sci. Nutr. 2020, 8, 1753–1765. [Google Scholar] [CrossRef] [PubMed]
- Sethi, M.; Singh, A.; Kaur, H.; Phagna, R.K.; Rakshit, S.; Chaudhary, D.P. Expression profile of protein fractions in the developing kernel of normal, Opaque-2 and quality protein maize. Sci. Rep. 2021, 11, 2469. [Google Scholar] [CrossRef] [PubMed]
- Paes, M.C.D.; Maga, J. Effect of extrusion on essential amino acids profile and color of whole-grain flours of quality protein maize (qpm) and normal maize cultivars. Rev. Bras. Milho Sorgo 2004, 3, 10–20. [Google Scholar] [CrossRef]
- Xiao, X.; Li, J.; Xiong, H.; Tui, W.; Zhu, Y.; Zhang, J. Effect of extrusion or fermentation on physicochemical and digestive properties of barley powder. Front. Nutr. 2022, 8, 794355. [Google Scholar] [CrossRef]
- Cui, L.; Li, D.-J.; Liu, C.-Q. Effect of fermentation on the nutritive value of maize. Int. J. Food Sci. Technol. 2012, 47, 755–760. [Google Scholar] [CrossRef]
- Thompson, H.O.; Onning, G.; Holmgren, K.; Strandler, S.; Hultberg, M. Fermentation of cauliflower and white beans with Lactobacillus plantarum—Impact on levels of riboflavin, folate, vitamin B-12, and amino acid composition. Plant Foods Hum. Nutr. 2020, 75, 236–242. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Sun, Y.; Zhang, Y.; Sun, Y.; Jin, T. Extrusion modification: Effect of extrusion on the functional properties and structure of rice protein. Processes 2022, 10, 1871. [Google Scholar] [CrossRef]
- Singh, N.; Kaur, M.; Singh, K.S. Physicochemical and functional properties of freeze-dried and oven dried corn gluten meals. Dry Technol. 2005, 23, 975–988. [Google Scholar] [CrossRef]
- Pedroche, J.; Yust, M.M.; Lqari, H.; Giron-Calle, J.; Alaiz, M.; Vioque, J.; Millan, F. Brassica carinata protein isolates: Chemical composition, protein characterization and improvement of functional properties by protein hydrolysis. Food Chem. 2004, 88, 337–346. [Google Scholar] [CrossRef]
- Ulloa, J.A.; Villalobos Barbosa, M.K.; Resendiz Vazquez, J.A.; Ulloa, P.R.; Ramírez Ramírez, J.C.; Carrillo, Y.S.; González Torres, L. Production, physico-chemical and functional characterization of a protein isolate from jackfruit (Artocarpus heterophyllus) seeds. CyTA J. Food 2017, 15, 497–507. [Google Scholar]
- Lawal, O.S.; Adebowale, K.O.; Ogunsanwo, B.M.; Sosanwo, O.A.; Bankole, S.A. On the functional properties of globulin and albumin protein fractions and flours of African locust bean (Parkia biglobossa). Food Chem. 2005, 92, 681–691. [Google Scholar] [CrossRef]
- Bora, P.S. Functional properties of native and succinylated lentil (Lens culinaris) globulins. Food Chem. 2002, 77, 171–176. [Google Scholar] [CrossRef]
- Liu, C.-M.; Peng, Q.; Zhong, J.-Z.; Liu, W.; Zhong, Y.-J.; Wang, F. Molecular and functional properties of protein fractions and isolate from cashew nut (Anacardium occidentale L.). Molecules 2018, 23, 393. [Google Scholar] [CrossRef] [Green Version]
- Sathe, S.K.; Deshpande, S.S.; Salunkhe, D.K. Functional properties of winged bean (Psophocarpus tetragonolobus (L.) DC) proteins. J. Food Sci. 1982, 47, 503–509. [Google Scholar] [CrossRef]
- Schwenzfeier, A.; Wierenga, A.P.; Gruppen, H. Isolation and characterization of soluble protein from the green microalgae Tetraselmis sp. Bioresour. Technol. 2011, 102, 9121–9127. [Google Scholar] [CrossRef]
- Hojilla-Evangelista, M.P. Extraction and functional properties of non-zein proteins in corn germ from wet-milling. J. Am. Oil Chem. Soc. 2012, 89, 167–174. [Google Scholar] [CrossRef]
- Maruatona, N.G.; Duodu, G.K.; Minnaar, A. Physicochemical, nutritional and functional properties of marama bean flour. Food Chem. 2010, 121, 400–405. [Google Scholar] [CrossRef]
- Saetae, D.; Kleekayai, T.; Jayasena, V.; Suntornsuk, W. Functional properties of protein isolate obtained from physic nut (Jatropha curcas L.) seed cake. Food Sci. Biotechnol. 2011, 20, 29–37. [Google Scholar] [CrossRef]
- Deb, S.; Kumar, Y.; Saxena, D.C. Functional, thermal and structural properties of fractionated protein from waste banana peel. Food Chem. X 2022, 13, 100205. [Google Scholar] [CrossRef] [PubMed]
- Jayasena, V.; Chih, H.J.; Nasar-Abbas, S.M. Functional properties of Australian sweet lupin protein isolated and tested at various pH levels. Res. J. Agric. Biol. Sci. 2010, 6, 130–137. [Google Scholar]
- Liu, C.; Wang, X.; Ma, H.; Zhang, Z.; Gao, W.; Xiao, L. Functional properties of protein isolates from soybeans stored under various conditions. Food. Chem. 2008, 111, 29–37. [Google Scholar] [CrossRef]
- Ngui, S.P.; Nyobe, C.E.; Bakwo Bassogog, C.B.; Tang, E.N.; Minka, S.R.; Mune Mune, M.A. Influence of pH and temperature on the physicochemical and functional properties of Bambara bean protein isolate. Heliyon 2021, 7, e07824. [Google Scholar] [CrossRef] [PubMed]
- Park, B.Y.; Yoon, K.Y. Functional properties of enzymatic hydrolysate and peptide fractions from perilla seed meal protein. Pol. J. Food Nutr. Sci. 2019, 69, 119–127. [Google Scholar] [CrossRef]
- Obatolu, V.A.; Asoyiro, S.B.; Ogunsunmi, L. Processing and functional properties of yam beans (Sphenostylis stenocarpa). J. Food Process. Preserv. 2007, 31, 240–249. [Google Scholar] [CrossRef]
- Lampart-Szczapa, E.; Konieczny, P.; Nogala-Kałucka, M.; Walczak, S.; Kossowska, I.; Malinowska, M. Some functional properties of lupin proteins modified by lactic fermentation and extrusion. Food Chem. 2006, 96, 290–296. [Google Scholar] [CrossRef]
- Elkhalifa, A.E.O.; Schiffler, B.; Bernhardt, R. Effect of fermentation on the functional properties of sorghum flour. Food Chem. 2005, 92, 1–5. [Google Scholar] [CrossRef]
- Tian, L.; Hu, S.; Jia, J.; Tan, W.; Yang, L.; Zhang, Q.; Liu, X.; Duan, X. Effects of short-term fermentation with lactic acid bacteria on the characterization, rheological and emulsifying properties of egg yolk. Food Chem. 2021, 341, 128163. [Google Scholar] [CrossRef]
- Zheng, X.-Q.; Li, L.-T.; Liu, X.-L.; Wang, X.-J.; Lin, J.; Li, D. Production of hydrolysate with antioxidative activity by enzymatic hydrolysis of extruded corn gluten. Appl. Microbiol. Biotechnol. 2006, 73, 763–770. [Google Scholar] [CrossRef]
- Elyas, S.H.A.; Tinay, A.H.; Yousif, E.N.; Elsheikh, E.E. Effect of natural fermentation on nutritive value and in vitro protein digestibility of pearl millet. Food Chem. 2002, 78, 75–79. [Google Scholar] [CrossRef]
- Krungleviciute, V.; Starkute, V.; Bartkiene, E.; Bartkevics, V.; Juodeikiene, G.; Vidmantiene, D.; Maknickiene, Z. Design of lupin seeds lactic acid fermentation—Changes of digestibility, amino acid profile and antioxidant activity. Vet. Zootech. 2016, 73, 47–52. [Google Scholar]
- Chu, J.; Zhao, H.; Lu, Z.; Lu, F.; Bie, X.; Zhang, C. Improved physicochemical and functional properties of dietary fiber from millet bran fermented by Bacillus natto. Food Chem. 2019, 294, 79–86. [Google Scholar] [CrossRef]
- Soria-Hernández, C.; Serna-Saldívar, S.; Chuck-Hernández, C. Physicochemical and functional properties of vegetable and cereal proteins as potential sources of novel food ingredients. Food Technol. Biotechnol. 2015, 53, 269–277. [Google Scholar] [CrossRef]
- Gammoh, S.; Aludatt, M.H.; Alhamad, M.N.; Rababah, T.; Ereifej, K.; Almajwal, A.; Ammar, Z.A.; Al Khateeb, W.; Hussein, N.M. Characterization of phenolic compounds extracted from wheat protein fractions using high-performance liquid chromatography/liquid chromatography mass spectrometry in relation to anti-allergenic, anti-oxidant, anti-hypertension, and anti-diabetic properties. Int. J. Food Prop. 2017, 20, 2383–2395. [Google Scholar] [CrossRef]
- Stanisavljević, N.S.; Vukotić, G.N.; Pastor, F.T.; Sužnjević, D.; Jovanović, Ž.S.; Strahinić, I.D.; Fira, Đ.A.; Radović, S.S. Antioxidant activity of pea protein hydrolysates produced by batch fermentation with lactic acid bacteria. Arch. Biol. Sci. 2015, 67, 1033–1042. [Google Scholar] [CrossRef]
- Tang, X.; He, Z.; Dai, Y.; Xiong, Y.L.; Xie, M.; Chen, J. Peptide fractionation and free radical scavenging activity of zein hydrolysate. J. Agric. Food Chem. 2010, 58, 587–593. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CMB | Protein | Carbohydrates | Free Sugars | Crude Fiber | Fat | Ash |
|---|---|---|---|---|---|---|
| Untreated | 12.14 ± 0.16 a | 77.37 ± 0.36 a | 1.13 ± 0.07 b | 1.24 ± 0.11 a | 4.36 ± 0.31 a | 4.89 ± 0.08 a |
| Extruded | 10.64 ± 0.11 b | 79.65 ± 0.64 a | 1.69 ± 0.06 a | 1.31 ± 0.08 a | 2.96 ± 0.17 b | 5.12 ± 0.06 a |
| CMB Samples | TCM, log10 CFU/g | Mass Density, g/cm3 | WAC, g/g | Damaged Starch, % | DG, % | Xylanase, XU/100 g dw | Protease, PU/100 g dw |
|---|---|---|---|---|---|---|---|
| Control | 6.89 ± 0.21 a | 0.547 a | 2.24 ± 0.02 b | 33.6 ± 0.1 b | 48.8 ± 0.7 b | 77.8 ± 2.7 a | 33.4 ± 0.9 a |
| Extruded | 1.56 ± 0.19 b | 0.481 b | 3.82 ± 0.01 a | 44.2 ± 0.1 a | 59.9 ± 0.8 a | 34.7 ± 1.2 b | 23.2 ± 1.1 b |
| Amino Acids | UN | E | F | EF |
|---|---|---|---|---|
| EAA | ||||
| Valine (VAL) | 38.8 a | 33.6 b | 39.4 a | 30.4 c |
| Isoleucine (ILE) | 35.0 a | 32.2 b | 34.5 a | 32.8 b |
| Leucine (LEU) | 73.5 b | 66.2 c | 72.0 b | 81.4 a |
| Tryptophan (TRP) | 3.6 b | 3.2 c | 4.6 a | 4.3 a |
| Lysine (LYS) | 25.0 b | 21.4 c | 33.5 a | 33.6 a |
| Methionine (MET) | 16.3 c | 15.4 d | 18.2 b | 22.4 a |
| Phenylalanine (PHE) | 52.6 a | 48.2 b | 53.5 a | 46.7 b |
| Threonine (THR) | 33.2 a | 29.9 b | 33.4 a | 31.2 b |
| Histidine (HIS) | 13.7 b | 12.7 c | 16.3 a | 12.2 c |
| Total EAA | 291.7 b | 262.8 c | 305.4 a | 295.0 b |
| NEAA and CEAA | ||||
| Alanine (ALA) | 52.9 ab | 49.5 c | 54.4 a | 57.0 a |
| Asparagine (ASP) | 44.2 a | 40.4 ab | 42.5 a | 39.8 b |
| Serine (SER) | 24.3 a | 22.1 b | 25.2 a | 21.7 b |
| Glutamine (GLU) | 128.4 c | 116.5 d | 162.2 a | 146.6 b |
| Cysteine (CYS) | 7.8 b | 7.3 bc | 8.6 a | 7.5 b |
| Proline (PRO) | 33.7 a | 30.2 b | 35.2 a | 34.5 a |
| Glycine (GLY) | 22.3 b | 20.4 c | 24.7 a | 18.3 d |
| Tyrosine (TYR) | 28.6 b | 26.1 c | 33.7 a | 34.9 a |
| Arginine (ARG) | 19.3 a | 17.6 b | 20.9 a | 13.5 c |
| Total NEAA and CEAA | 361.5 bc | 330.1 d | 397.4 a | 373.8 b |
| Total Amino Acids | 653.2 b | 592.9 c | 712.8 a | 668.8 b |
| Samples | Albumins | Globulins | ||||
|---|---|---|---|---|---|---|
| pH 4 | pH 7 | pH 9 | pH 4 | pH 7 | pH 9 | |
| Untreated | ||||||
| WAC | 2.42 ± 0.12 bc | 2.52 ± 0.11 b | 2.64 ± 0.08 a | 2.12 ± 0.07 d | 2.27 ± 0.10 c | 2.47 ± 0.06 b |
| WS | 34.9 ± 0.1 c | 64.8 ± 0.2 b | 73.7 ± 0.3 a | 39.4 ± 0.1 d | 63.5 ± 0.2 b | 72.3 ± 0.2 a |
| FFC | 249 ± 4 c | 266 ± 3 b | 324 ± 6 a | 178 ± 3 e | 186 ± 2 e | 209 ± 3 d |
| FS | 56.4 ± 0.5 d | 69.6 ± 0.7 a | 63.2 ± 0.4 b | 55.7 ± 0.6 d | 64.8 ± 0.3 b | 58.3 ± 0.2 c |
| EC | 46.5 ± 0.2 c | 52.2 ± 0.1 b | 59.6 ± 0.5 a | 39.4 ± 0.4 e | 43.8 ± 0.6 d | 49.5 ± 0.3 a |
| ES | 45.4 ± 0.8 c | 50.5 ± 0.7 b | 56.8 ± 0.2 a | 38.3 ± 0.1 de | 39.2 ± 0.4 d | 42.9 ± 0.5 d |
| Extruded | ||||||
| WAC | 2.54 ± 0.13 b | 2.72 ± 0.14 a | 2.68 ± 0.11 a | 2.38 ± 0.08 c | 2.61 ± 0.12 b | 2.59 ± 0.10 b |
| WS | 37.6 ± 0.6 d | 78.2 ± 0.4 b | 88.6 ± 1.2 a | 38.7 ± 0.9 e | 76.7 ± 0.8 b | 87.4 ± 1.1 a |
| FFC | 215 ± 5 c | 238 ± 4 b | 256 ± 6 a | 141 ± 3 f | 152 ± 3 e | 179± 2 d |
| FS | 53.8 ± 0.2 c | 65.1 ± 0.6 a | 59.2 ± 0.8 b | 52.1 ± 0.7 d | 60.3 ± 0.4 b | 54.8 ± 0.3 c |
| EC | 40.7 ± 0.2 b | 42.8 ± 0.3 b | 44.3 ± 0.3 a | 33.4 ± 0.4 e | 35.5 ± 0.3 e | 38.9 ± 0.2 d |
| ES | 39.3 ± 0.1 b | 41.2 ± 0.2 b | 43. 6 ± 0.4 a | 32.7 ± 0.2 d | 34.2 ± 0.1 d | 36.7 ± 0.2 c |
| Samples | Albumins | Globulins | ||||||
|---|---|---|---|---|---|---|---|---|
| L. sakei | P. acidilactici | L. sakei | P. acidilactici | |||||
| 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | |
| WAC | 3.40 ± 0.12 a | 3.11 ± 0.09 b | 3.30 ± 0.10 a | 3.22 ± 0.07 ab | 2.89 ± 0.04 c | 2.92 ± 0.07 c | 2.96 ± 0.10 c | 3.02 ± 0.11 bc |
| FFC | 287 ± 2 a | 295 ± 2 a | 288 ± 2 a | 290± 2 a | 192 ± 1 d | 189 ± 1 d | 223 ± 2 b | 211 ± 2 c |
| FS | 78.8 ± 0.6 a | 68.7 ± 0.5 c | 82.4 ± 0.7 a | 69.2 ± 0.3 c | 72.9 ± 0.5 b | 56.8 ± 0.6 d | 73.5 ± 0.5 b | 56.1 ± 0.2 d |
| EC | 47.6 ± 0.3 a | 48.7 ± 0.5 a | 48.1 ± 0.6 a | 49.6 ± 0.4 a | 41.8 ± 0.2 b | 42.7 ± 0.5 b | 40.6 ± 0.7 b | 42.3 ± 0.6 b |
| ES | 46.5 ± 0.2 a | 47.0 ± 0.4 a | 46.0 ± 0.6 a | 47.5 ± 0.7 a | 37.6 ± 0.3 c | 39.8 ± 0.5 b | 38.9 ± 0.5 bc | 40.9 ± 0.7 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zadeike, D.; Gaizauskaite, Z.; Svazas, M.; Gruzauskas, R.; Gruzauskas, V.; Damasius, J.; Juodeikiene, G. Application of Solid-State Fermentation for the Improving of Extruded Corn Dry-Milling By-Products and Their Protein Functional Properties. Life 2022, 12, 1909. https://doi.org/10.3390/life12111909
Zadeike D, Gaizauskaite Z, Svazas M, Gruzauskas R, Gruzauskas V, Damasius J, Juodeikiene G. Application of Solid-State Fermentation for the Improving of Extruded Corn Dry-Milling By-Products and Their Protein Functional Properties. Life. 2022; 12(11):1909. https://doi.org/10.3390/life12111909
Chicago/Turabian StyleZadeike, Daiva, Zydrune Gaizauskaite, Mantas Svazas, Romas Gruzauskas, Valentas Gruzauskas, Jonas Damasius, and Grazina Juodeikiene. 2022. "Application of Solid-State Fermentation for the Improving of Extruded Corn Dry-Milling By-Products and Their Protein Functional Properties" Life 12, no. 11: 1909. https://doi.org/10.3390/life12111909