
Uncovering the Inhibitory Molecular Mechanism of Pomegranate Peel to Urinary Bladder Urothelial Carcinoma Using Proteomics Techniques

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Preparation of Protein Lysates
2.3. Two-Dimensional Gel Electrophoresis (2-DE)—Isoelectric Focusing (IEF) and SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.4. Silver Staining
2.5. Image Analysis and Statistical Analysis
2.6. Protein Recognition
2.7. Western Immunoblotting
2.8. Immunohistochemical Staining
2.9. Immunohistochemical Staining
3. Results
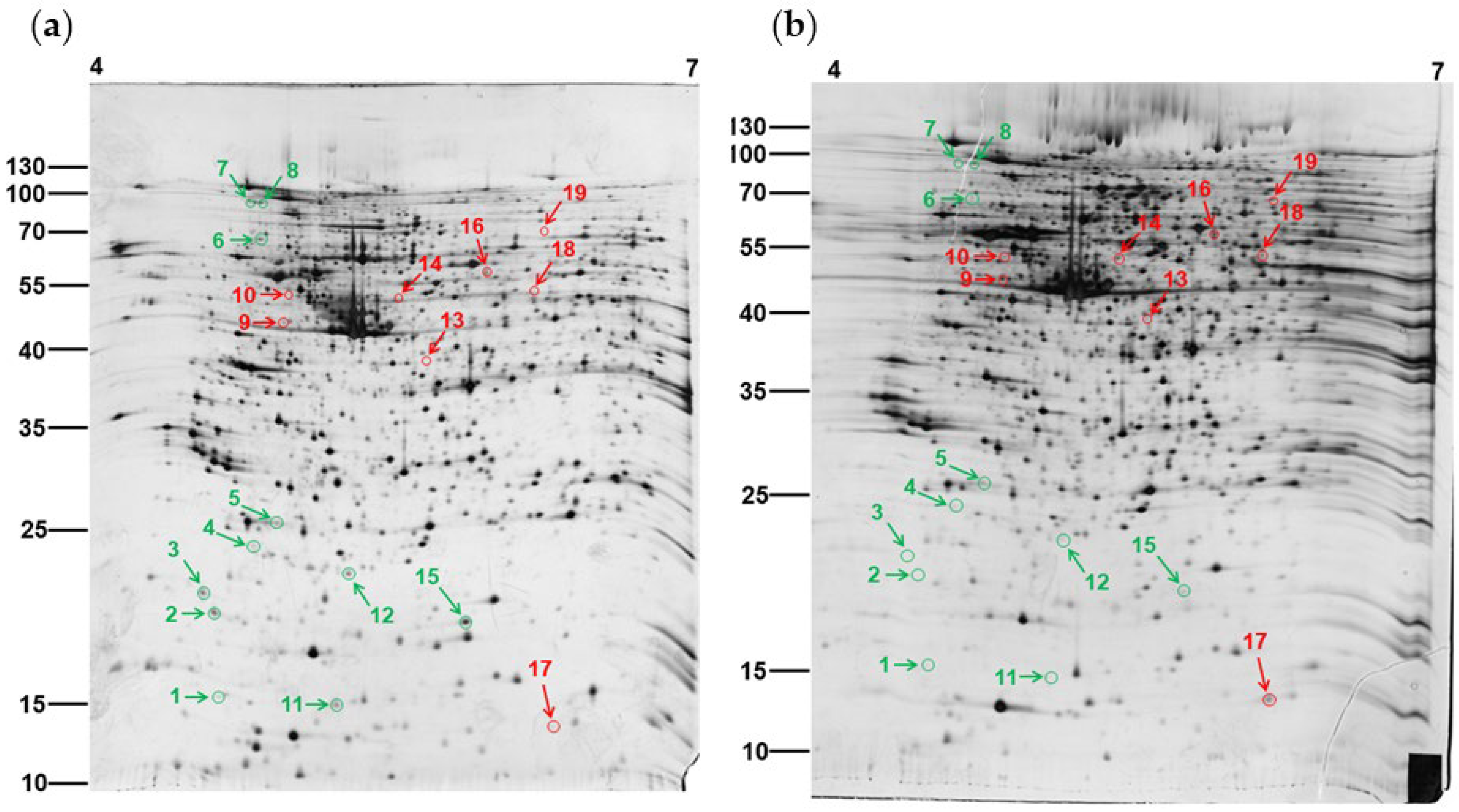
3.1. Two-Dimensional Gel Electrophoresis of PEPE2-Treated T24 Cells
3.2. Recognition of the Differentially Expressed Proteins in PEPE2-Influenced T24 Cells
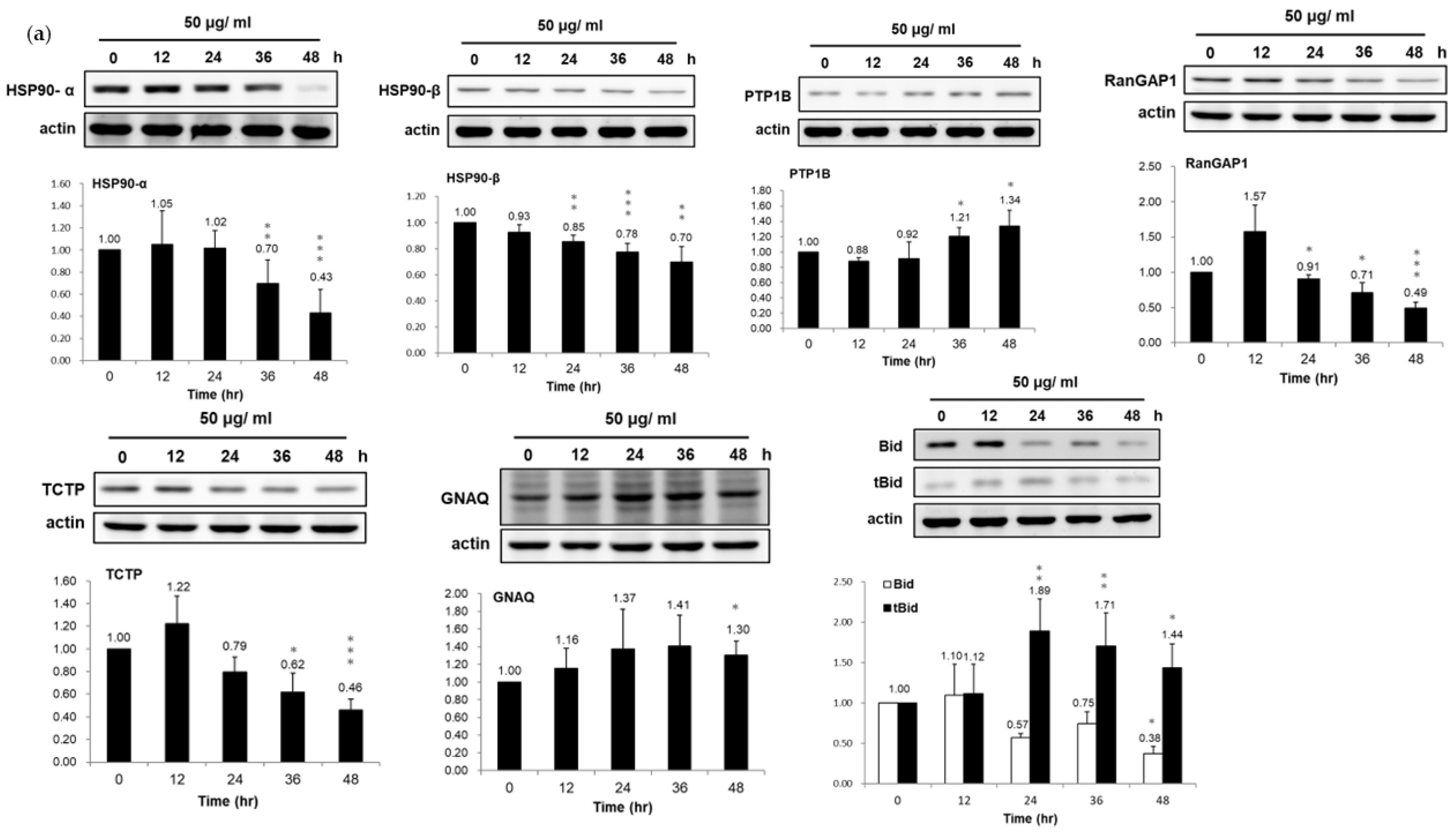
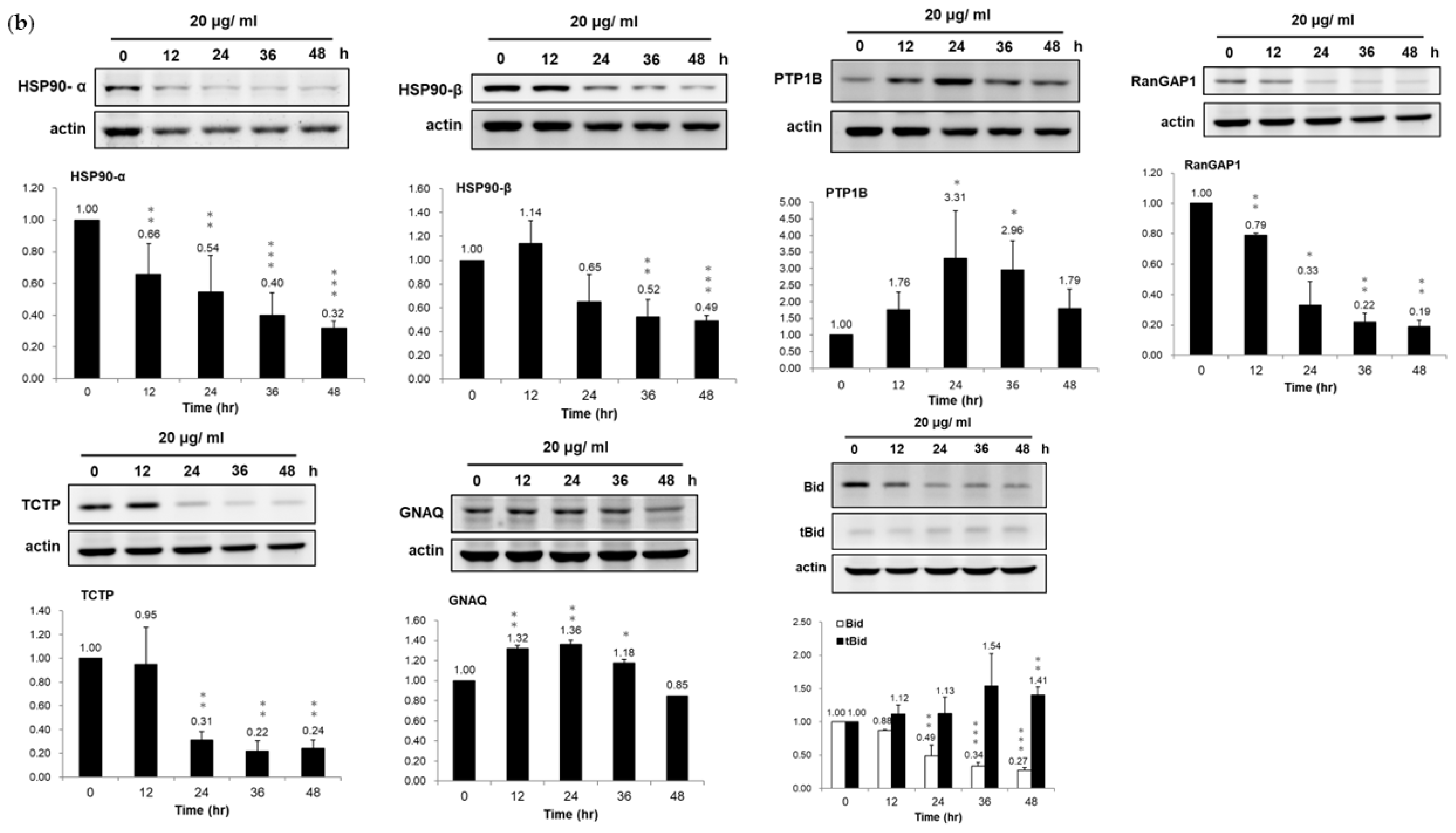
3.3. Validation of Dysregulated Proteins
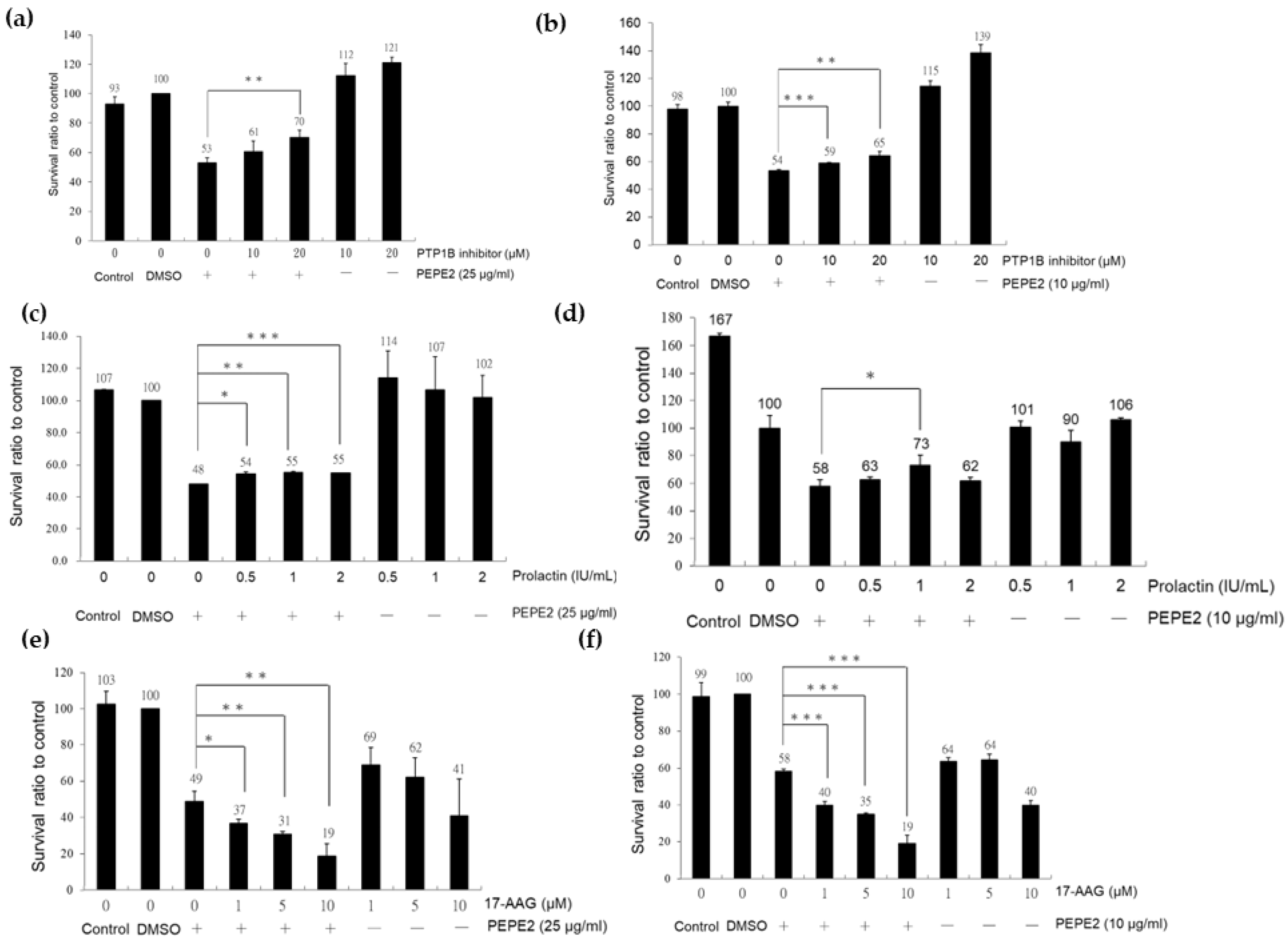
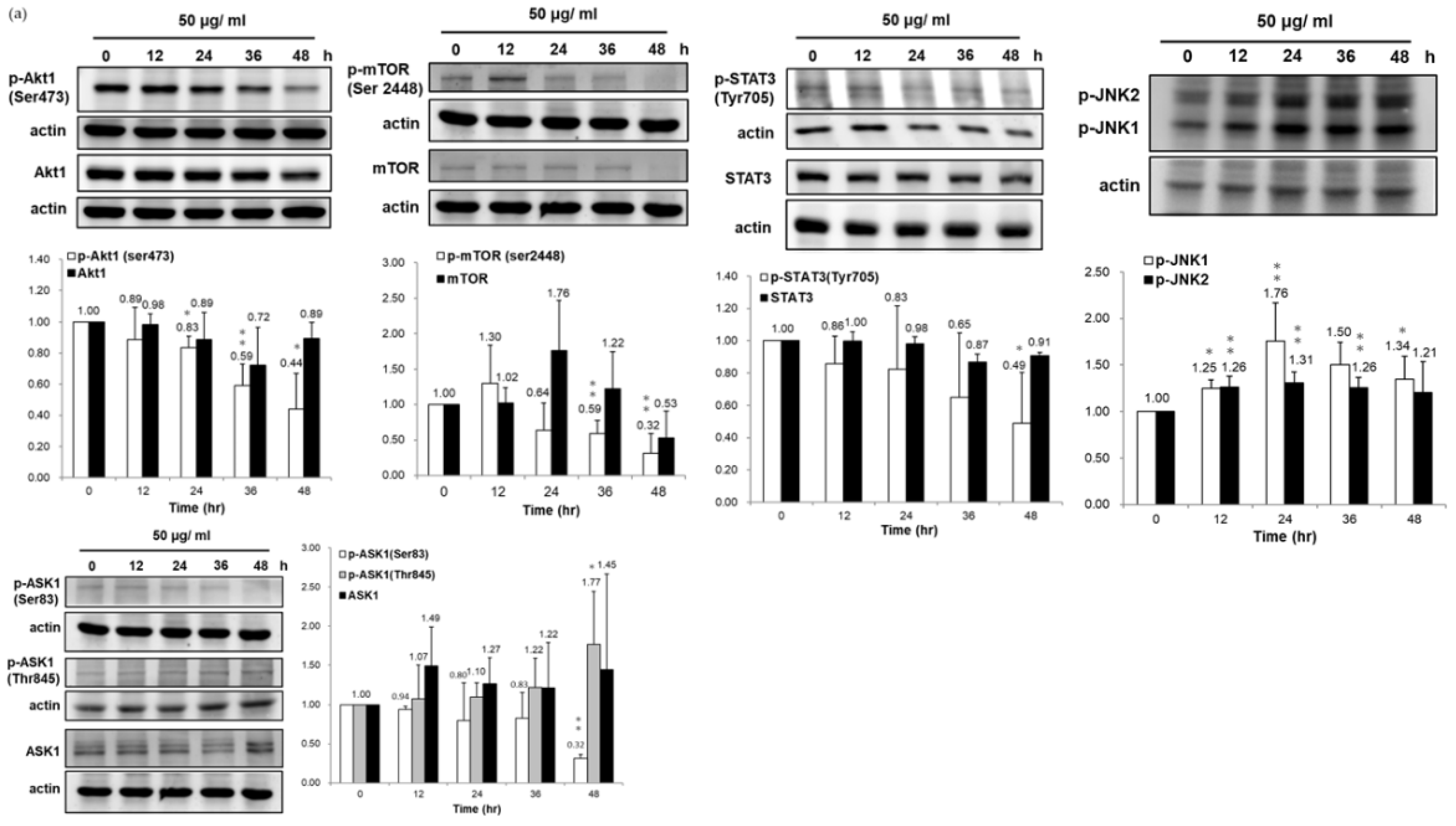
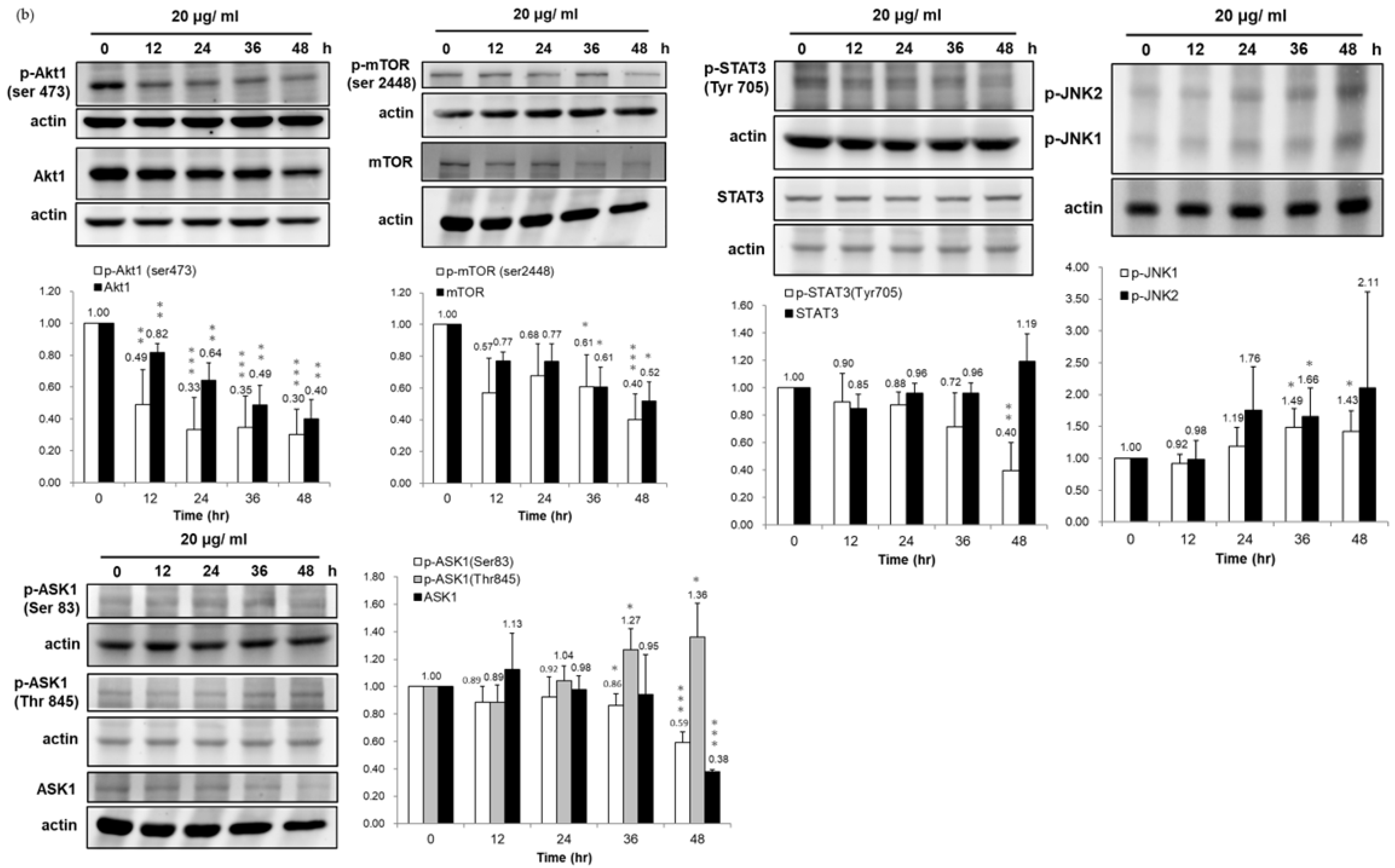
3.4. Association of HSP90/Akt/ASK-1/JNK Pathway with Bladder Cancer Cell Apoptosis
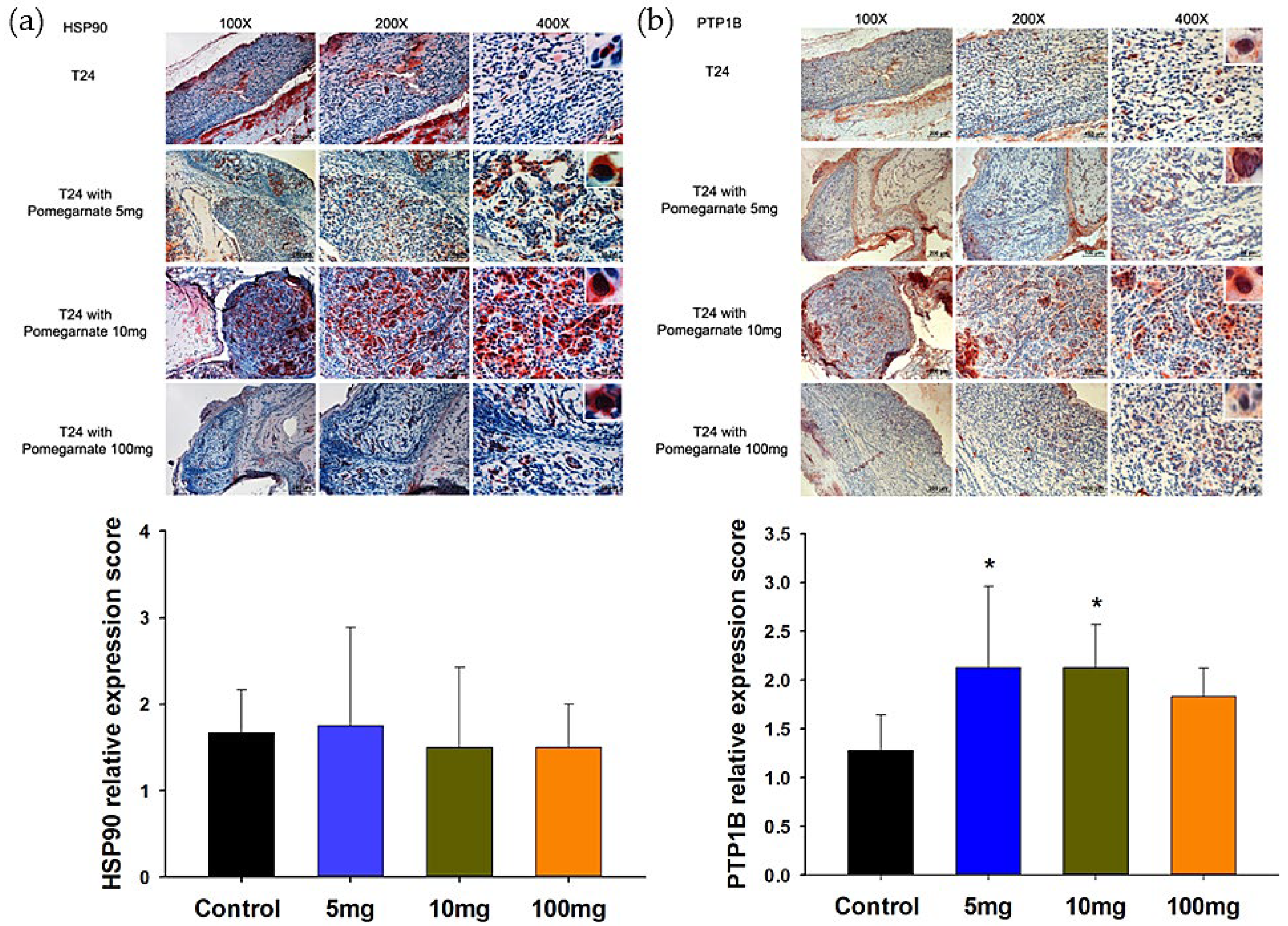
3.5. The Validation of PTPB1 and HSP90α Expression in Xenografted Bladder Tumor Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33, Erratum in: CA Cancer J. Clin. 2021, 71, 359. [Google Scholar] [CrossRef] [PubMed]
- Health Promotion Administration, Ministry of Health and Welfare, Taiwan, Cancer Registry Annual Report. 2018. Available online: https://www.hpa.gov.tw/Pages/Detail.aspx?nodeid=269&pid=13498 (accessed on 30 May 2022).
- Zieger, K.; Wolf, H.; Olsen, P.; Højgaard, K. Long-term follow-up of noninvasive bladder tumours(stage Ta): Recurrence and progression. Br. J. Urol. 2001, 85, 824–828. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; McClees, S.F.; Afaq, F. Pomegranate for Prevention and Treatment of Cancer: An Update. Molecules 2017, 22, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Hadi, N.; Afaq, F.; Syed, D.N.; Kweon, M.-H.; Mukhtar, H. Pomegranate fruit extract inhibits prosurvival pathways in human A549 lung carcinoma cells and tumor growth in athymic nude mice. Carcinogenesis 2007, 28, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Afaq, F.; Kweon, M.-H.; Kim, K.; Mukhtar, H. Oral Consumption of Pomegranate Fruit Extract Inhibits Growth and Progression of Primary Lung Tumors in Mice. Cancer Res. 2007, 67, 3475–3482. [Google Scholar] [CrossRef] [Green Version]
- Malik, A.; Afaq, F.; Sarfaraz, S.; Adhami, V.M.; Syed, D.N.; Mukhtar, H. Pomegranate fruit juice for chemoprevention and chemotherapy of prostate cancer. Proc. Natl. Acad. Sci. USA 2005, 102, 14813–14818. [Google Scholar] [CrossRef] [Green Version]
- Adhami, V.M.; Siddiqui, I.A.; Syed, D.N.; Lall, R.K.; Mukhtar, H. Oral infusion of pomegranate fruit extract inhibits prostate carcinogenesis in the TRAMP model. Carcinogenesis 2011, 33, 644–651. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-T.; Wu, Y.-L.; Chien, L.-H.; Chen, S.-T.; Tzeng, Y.-K.; Wu, T.-F. Proteomic exploration of the impacts of pomegranate fruit juice on the global gene expression of prostate cancer cell. Proteomics 2012, 12, 3251–3262. [Google Scholar] [CrossRef]
- Koyama, S.; Cobb, L.J.; Mehta, H.H.; Seeram, N.P.; Heber, D.; Pantuck, A.J.; Cohen, P. Pomegranate extract induces apoptosis in human prostate cancer cells by modulation of the IGF–IGFBP axis. Growth Horm. IGF Res. 2010, 20, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Rettig, M.B.; Heber, D.; An, J.; Seeram, N.P.; Rao, J.Y.; Liu, H.; Klatte, T.; Belldegrun, A.; Moro, A.; Henning, S.M.; et al. Pomegranate extract inhibits androgen-independent prostate cancer growth through a nuclear factor-κB-dependent mechanism. Mol. Cancer Ther. 2008, 7, 2662–2671. [Google Scholar] [CrossRef]
- Pantuck, A.J.; Leppert, J.T.; Zomorodian, N.; Aronson, W.; Hong, J.; Barnard, R.J.; Seeram, N.; Liker, H.; Wang, H.; Elashoff, R.; et al. Phase II Study of Pomegranate Juice for Men with Rising Prostate-Specific Antigen following Surgery or Radiation for Prostate Cancer. Clin. Cancer Res. 2006, 12, 4018–4026. [Google Scholar] [CrossRef] [Green Version]
- Pantuck, A.J.; Pettaway, C.A.; Dreicer, R.; Corman, J.M.; Katz, A.; Ho, A.; Aronson, W.J.; Clark, W.; Simmons, G.W.; Heber, D. A randomized, double-blind, placebo-controlled study of the effects of pomegranate extract on rising PSA levels in men following primary therapy for prostate cancer. Prostate Cancer Prostatic Dis. 2015, 18, 242–248. [Google Scholar] [CrossRef]
- Chang, C.-P.; Chan, Y.-Y.; Li, C.-F.; Chien, L.-H.; Lee, S.-T.; Wu, T.-F. Deciphering the Molecular Mechanism Underlying the Inhibitory Efficacy of Taiwanese Local Pomegranate Peels against Urinary Bladder Urothelial Carcinoma. Nutrients 2018, 10, 543. [Google Scholar] [CrossRef] [Green Version]
- Sheng, K.-H.; Yao, Y.-C.; Chuang, S.-S.; Wu, H.; Wu, T.-F. Search for the tumor-related proteins of transition cell carcinoma in Taiwan by proteomic analysis. Proteomics 2006, 6, 1058–1065. [Google Scholar] [CrossRef]
- Li, C.-F.; Shen, K.-H.; Chien, L.-H.; Huang, C.-H.; Wu, T.-F.; He, H.-L. Proteomic Identification of the Galectin-1-Involved Molecular Pathways in Urinary Bladder Urothelial Carcinoma. Int. J. Mol. Sci. 2018, 19, 1242. [Google Scholar] [CrossRef] [Green Version]
- Lanneau, D.; Brunet, M.; Frisan, E.; Solary, E.; Fontenay, M.; Garrido, C. Heat shock proteins: Essential proteins for apoptosis regulation. J. Cell. Mol. Med. 2008, 12, 743–761. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wulfkuhle, J.; Zhang, H.; Gu, P.; Yang, Y.; Deng, J.; Margolick, J.B.; Liotta, L.A.; Petricoin, E., 3rd; Zhang, Y. Activation of the PTEN/mTOR/STAT3 pathway in breast cancer stem-like cells is required for viability and maintenance. Proc. Natl. Acad. Sci. USA 2007, 104, 16158–16163. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Luo, D.; Miao, R.; Bai, L.; Ge, Q.; Sessa, W.C.; Min, W. Hsp90–Akt phosphorylates ASK1 and inhibits ASK1-mediated apoptosis. Oncogene 2005, 24, 3954–3963. [Google Scholar] [CrossRef] [Green Version]
- Madan, E.; Gogna, R.; Kuppusamy, P.; Bhatt, M.; Mahdi, A.A.; Pati, U. SCO2 Induces p53-Mediated Apoptosis by Thr 845 Phosphorylation of ASK-1 and Dissociation of the ASK-1–Trx Complex. Mol. Cell. Biol. 2013, 33, 1285–1302. [Google Scholar] [CrossRef] [Green Version]
- Kolosenko, I.; Grander, D.; Tamm, K. IL-6 activated JAK/STAT3 pathway and sensitivity to Hsp90 inhibitors in multiple myeloma. Curr. Med. Chem. 2014, 21, 3042–3047. [Google Scholar] [CrossRef]
- Tsunekawa, T.; Banno, R.; Mizoguchi, A.; Sugiyama, M.; Tominaga, T.; Onoue, T.; Hagiwara, D.; Ito, Y.; Iwama, S.; Goto, M.; et al. Deficiency of PTP1B Attenuates Hypothalamic Inflammation via Activation of the JAK2-STAT3 Pathway in Microglia. eBioMedicine 2017, 16, 172–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-J.; Song, H.; Yoon, Y.J.; Park, S.-J.; Kim, S.-Y.; Han, D.C.; Kwon, B.-M. Ethacrynic acid inhibits STAT3 activity through the modulation of SHP2 and PTP1B tyrosine phosphatases in DU145 prostate carcinoma cells. Biochem. Pharmacol. 2020, 175, 113920. [Google Scholar] [CrossRef] [PubMed]
- Lund, I.K.; Hansen, J.A.; Andersen, H.S.; Møller, N.P.H.; Billestrup, N. Mechanism of protein tyrosine phosphatase 1B-mediated inhibition of leptin signalling. J. Mol. Endocrinol. 2005, 34, 339–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warabi, M.; Nemoto, T.; Ohashi, K.; Kitagawa, M.; Hirokawa, K. Expression of Protein Tyrosine Phosphatases and Its Significance in Esophageal Cancer. Exp. Mol. Pathol. 2000, 68, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Dubé, N.; Bourdeau, A.; Heinonen, K.M.; Cheng, A.; Loy, A.L.; Tremblay, M.L. Genetic Ablation of Protein Tyrosine Phosphatase 1B Accelerates Lymphomagenesis of p53-Null Mice through the Regulation of B-Cell Development. Cancer Res. 2005, 65, 10088–10095. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Meza, S.; Díaz, J.; Sandoval-Bórquez, A.; Valenzuela-Valderrama, M.; Díaz-Valdivia, N.; Rojas-Celis, V.; Contreras, P.; Huilcaman, R.; Ocaranza, M.P.; Chiong, M.; et al. AT2 Receptor Mediated Activation of the Tyrosine Phosphatase PTP1B Blocks Caveolin-1 Enhanced Migration, Invasion and Metastasis of Cancer Cells. Cancers 2019, 11, 1299. [Google Scholar] [CrossRef] [Green Version]
- Koziol, M.J.; Gurdon, J.B. TCTP in Development and Cancer. Biochem. Res. Int. 2012, 2012, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Schadendorf, D.; Fisher, D.E.; Garbe, C.; Gershenwald, J.E.; Grob, J.-J.; Halpern, A.; Herlyn, M.; Marchetti, M.A.; McArthur, G.; Ribas, A.; et al. Melanoma. Nat. Rev. Dis. Prim. 2015, 1, 15003. [Google Scholar] [CrossRef]
- Chattopadhyay, C.; Kim, D.W.; Gombos, D.S.; Oba, J.; Qin, Y.; Williams, M.D.; Esmaeli, B.; Grimm, E.A.; Wargo, J.A.; Woodman, S.E.; et al. Uveal melanoma: From diagnosis to treatment and the science in between. Cancer 2016, 122, 2299–2312. [Google Scholar] [CrossRef] [Green Version]
- Overholtzer, M.; Zhang, J.; Smolen, G.A.; Muir, B.; Li, W.; Sgroi, D.C.; Deng, C.-X.; Brugge, J.S.; Haber, D.A. Transforming properties of YAP, a candidate oncogene on the chromosome 11q22 amplicon. Proc. Natl. Acad. Sci. USA 2006, 103, 12405–12410. [Google Scholar] [CrossRef]
- Dong, J.; Feldmann, G.; Huang, J.; Wu, S.; Zhang, N.; Comerford, S.A.; Gayyed, M.F.; Anders, R.A.; Maitra, A.; Pan, D. Elucidation of a Universal Size-Control Mechanism in Drosophila and Mammals. Cell 2007, 130, 1120–1133. [Google Scholar] [CrossRef] [Green Version]
- Nobes, C.D.; Hall, A. Rho, Rac, and Cdc42 GTPases regulate the assembly of multimolecular focal complexes associated with actin stress fibers, lamellipodia, and filopodia. Cell 1995, 81, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Matsunuma, R.; Chan, D.W.; Kim, B.-J.; Singh, P.; Han, A.; Saltzman, A.B.; Cheng, C.; Lei, J.T.; Wang, J.; da Silva, L.R.; et al. DPYSL3 modulates mitosis, migration, and epithelial-to-mesenchymal transition in claudin-low breast cancer. Proc. Natl. Acad. Sci. USA 2018, 115, E11978–E11987. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Jiang, Y.; Xie, D.; Liu, M.; Song, N.; Zhu, J.; Fan, J.; Zhu, C. Inhibition of cell-adhesion protein DPYSL3 promotes metastasis of lung cancer. Respir. Res. 2018, 19, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kanda, M.; Nomoto, S.; Oya, H.; Shimizu, D.; Takami, H.; Hibino, S.; Hashimoto, R.; Kobayashi, D.; Tanaka, C.; Yamada, S.; et al. Dihydropyrimidinase-like 3 facilitates malignant behavior of gastric cancer. J. Exp. Clin. Cancer Res. 2014, 33, 66. [Google Scholar] [CrossRef]
- Li, B.; Li, C. Suppression of Prostate Cancer Metastasis by DPYSL3-Targeted saRNA. RNA Act. 2017, 983, 207–216. [Google Scholar] [CrossRef]
- Wagner, K.-D.; Wagner, N. Peroxisome proliferator-activated receptor beta/delta (PPARβ/δ) acts as regulator of metabolism linked to multiple cellular functions. Pharmacol. Ther. 2010, 125, 423–435. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot | Protein Identity | Incidences | Experiment PI/MW | Theoretical PI/MW | Matched Peptide Number | Coverage (%) 1 | Accession Number (NCBI) | Fold |
|---|---|---|---|---|---|---|---|---|
| 1 | myosin light polypeptide 6 (MYL6) | 6/6 | 4.6/16.8 | 4.56/17 | 3 | 43.1 | 119617305 | −8.9 |
| 2 | myosin regulatory light chain MRLC2 (MYL2) | 6/6 | 4.6/20.6 | 4.67/19.7 | 7 | 48.84 | 119622081 | −8.1 |
| 3 | coatomer protein complex, subunit zeta 1 (COPZ1) | 5/6 | 4.6/21.8 | 4.69/20.2 | 3 | 28.81 | 119617180 | −50.7 |
| 4 | Chromobox protein homolog 3 (CBX3) | 5/6 | 4.8/24.9 | 5.23/20.8 | 6 | 33.33 | 116241284 | −3.7 |
| 5 | Human Translationally Controlled Tumor Protein (TCTP) | 5/6 | 4.9/26.3 | 5.24/20.7 | 3 | 21.11 | 114794484 | −7.3 |
| 6 | Ran GTPase-activating protein 1 (RanGAP1) | 5/6 | 4.8/66.7 | 4.63/63.5 | 14 | 30.83 | 1172922 | −4 |
| 7 | Heat shock protein HSP 90-beta (HSP90-β) | 5/6 | 4.8/82.8 | 4.96/83.3 | 10 | 20.02 | 17865718 | −4.1 |
| 8 | heat shock protein HSP 90-alpha (HSP90-α) | 5/6 | 4.8/82.7 | 5.11/77.3 | 6 | 9.95 | 153792590 | −2.5 |
| 9 | vimentin (V.I.M.) | 6/6 | 4.9/44.2 | 5.41/42.8 | 13 | 36.69 | 119606621 | 2.6 |
| 10 | Mitochondrial ATP synthase subunit beta (ATP5B) | 5/6 | 4.9/50.8 | 5.26/56.6 | 18 | 50.28 | 114549 | 9.1 |
| 11 | Coactosin-like Protein-1 (COTL1) | 5/6 | 5.4/16.7 | 5.5/15.9 | 7 | 60.56 | 119615882 | −14.1 |
| 12 | Pro-Apoptotic Protein Bid (Bid) | 5/6 | 5.4/23.6 | 5.25/22.1 | 2 | 13.7 | 159163783 | −6.1 |
| 13 | Guanine nucleotide-binding protein alpha-q (GNAQ) | 5/6 | 6.2/40.8 | 5.48/42.1 | 6 | 18.94 | 251757492 | 4.6 |
| 14 | RuvB-like 2; 48 kDa TATA box-binding protein interacting protein (RUVBL2) | 5/6 | 5.9/52.6 | 5.49/51.2 | 12 | 35.2 | 28201890 | 3.8 |
| 15 | Superoxide dismutase 1 (SOD1) | 5/6 | 6.4/21.6 | 5.7/15.9 | 4 | 57.14 | 134611 | −4.9 |
| 16 | Human Prolidase (PEPD) | 5/6 | 6.5/58.4 | 5.64/54.6 | 6 | 14.57 | 112491419 | 2.7 |
| 17 | Fatty acid-binding protein 4 (FABP4) | 5/6 | 6.6/16.2 | 6.59/14.7 | 2 | 48.48 | 119781 | 14.6 |
| 18 | protein-tyrosine phosphatase 1B (PTP1B) | 5/6 | 6.6/51.4 | 5.88/50 | 7 | 32.87 | 131467 | 7.6 |
| 19 | dihydropyrimidinase-related protein 3 (DPYSL3) | 5/6 | 6.6/71.4 | 5.94/73.9 | 10 | 33.18 | 308818200 | 5 |
| Protein Function | Protein 2 |
|---|---|
| Carbohydrate metabolism | PTP1B, GNAQ, SOD1, BID, FABP4 |
| Cell apoptosis | PTP1B, GNAQ, SOD1, B.I.D., FABP4, HSP90-α, HSP90-β, TCTP, DPYSL3 |
| Proliferation of cells | DPYSL3, FABP4, GNAQ, HSP90-α, HSP90-β, PTP1B, RUVBL2, SOD1, TCTP, ATP5B, B.I.D., COPZ1 |
| Oxidation of fatty acid | B.I.D., FABP4 |
| Synthesis of ATP | SOD1, ATP5B |
| Catabolism of ATP | ATP5B, RUVBL2 |
| Death receptor signaling | BID |
| eNOS signaling | HSP90-α, HSP90-β, GNAQ |
| JAK/STAT signaling | PTP1B, GNAQ, HSP90-α, HSP90-β |
| PPAR pathway | HSP90-α, HSP90-β, PTP1B |
| PPARα/R.X.R. α pathway | HSP90-α, HSP90-β,GNAQ |
| Rho family GTPase signaling | GNAQ, MYL2 |
| RhoGDI signaling | GNAQ, MYL2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, K.-H.; Chang, C.-P.; Chien, L.-H.; Li, C.-F.; Tang, L.-Y.; Chan, Y.-Y.; Wu, T.-F. Uncovering the Inhibitory Molecular Mechanism of Pomegranate Peel to Urinary Bladder Urothelial Carcinoma Using Proteomics Techniques. Life 2022, 12, 1839. https://doi.org/10.3390/life12111839
Huang K-H, Chang C-P, Chien L-H, Li C-F, Tang L-Y, Chan Y-Y, Wu T-F. Uncovering the Inhibitory Molecular Mechanism of Pomegranate Peel to Urinary Bladder Urothelial Carcinoma Using Proteomics Techniques. Life. 2022; 12(11):1839. https://doi.org/10.3390/life12111839
Chicago/Turabian StyleHuang, Kuan-Hua, Ching-Ping Chang, Lan-Hsiang Chien, Chien-Feng Li, Ling-Yu Tang, Yu-Yi Chan, and Ting-Feng Wu. 2022. "Uncovering the Inhibitory Molecular Mechanism of Pomegranate Peel to Urinary Bladder Urothelial Carcinoma Using Proteomics Techniques" Life 12, no. 11: 1839. https://doi.org/10.3390/life12111839