
Natural Strategies as Potential Weapons against Bacterial Biofilms

 ,
,  , ,
, ,  , , ,
, , ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
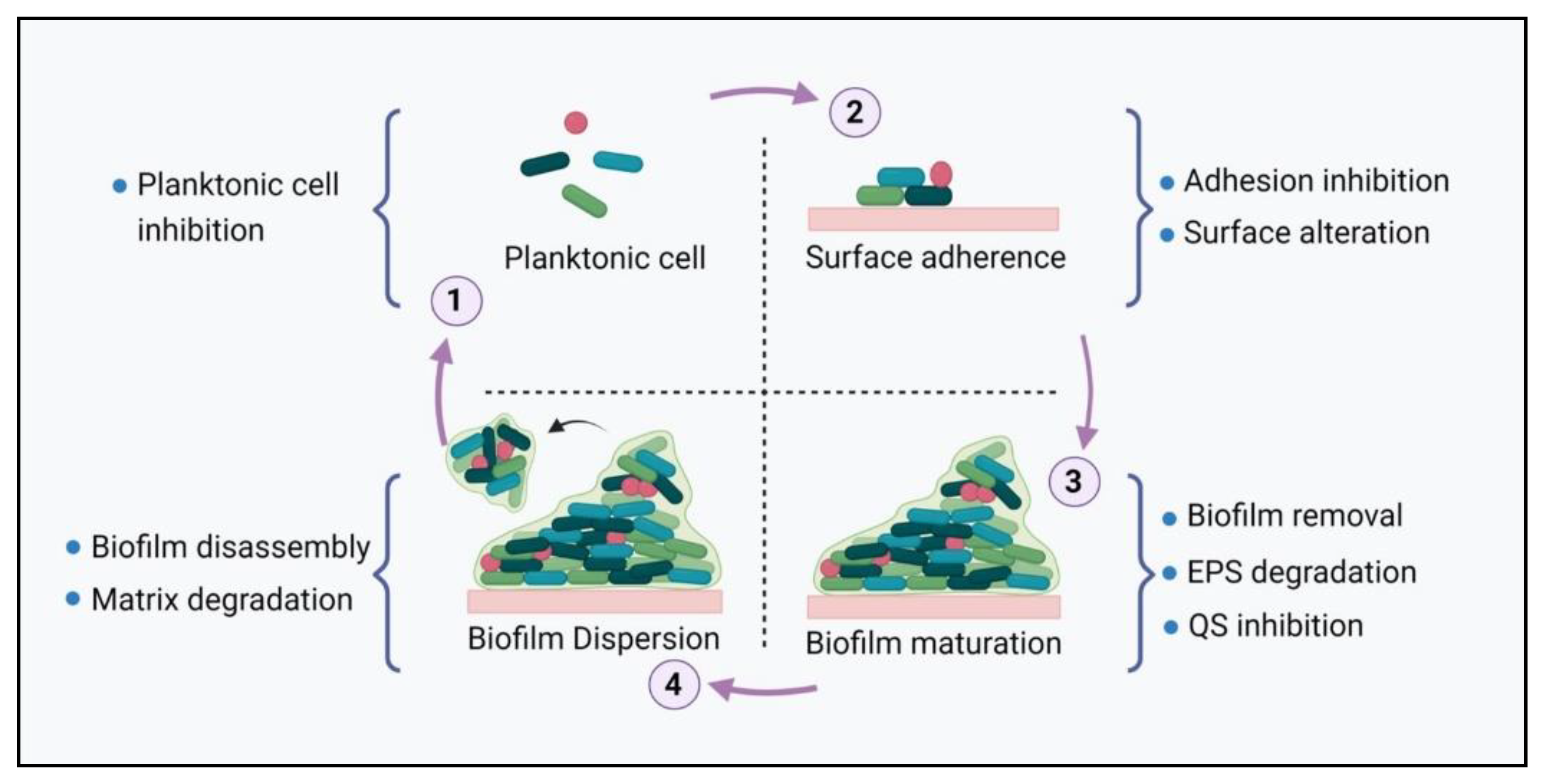
2. Biofilm Formation
2.1. Surface Adhesion
2.2. Biofilm Maturation
2.3. Biofilm Dispersal
3. Biofilm Composition
3.1. Polysaccharides
3.2. Extracellular Proteins
3.3. Extracellular DNA (eDNA)
3.4. Surfactants and Lipids
3.5. Water
4. Biofilm Structural Stability Parameters
4.1. Proteins
4.2. Genes and Signaling Cascades
5. Resistance Mechanism in Biofilms
5.1. The Structural Complexity of Biofilms
5.2. The Heterogeneity of Biofilms
5.3. Quorum Sensing
5.4. Enhanced Efflux Pumps
6. Pathogenicity of Biofilm Microbes
6.1. Health Problems and Infections Caused by Biofilm Bacteria
6.2. Biofilms in the Food Industry
7. Biofilm Control
8. Biofilm Controlling Compounds
8.1. Natural Plants and Bee Products
8.2. Nanotechnology and Phyto-Nanotechnology
8.2.1. Liposomes
8.2.2. Solid Lipid NPs
8.2.3. QS Inhibiting NMs
8.2.4. Cyclodextrins
8.2.5. Hydrogels
8.3. Microbes and Marine-Derived Anti-Biofilm Compounds
9. Methodology
Data Collection Criteria
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Percival, S.L.; Malic, S.; Cruz, H.; Williams, D.W. Introduction to biofilms. In Biofilms and Veterinary Medicine; Springer: Berlin/Heidelberg, Germany, 2011; pp. 41–68. [Google Scholar]
- Asma, S.T.; Imre, K.; Morar, A.; Herman, V.; Acaroz, U.; Mukhtar, H.; Arslan-Acaroz, D.; Shah, S.R.A.; Gerlach, R. An overview of biofilm formation–combating strategies and mechanisms of action of antibiofilm agents. Life 2022, 12, 1110. [Google Scholar] [CrossRef] [PubMed]
- Burmølle, M.; Ren, D.; Bjarnsholt, T.; Sørensen, S.J. Interactions in multispecies biofilms: Do they actually matter? Trends Microbiol. 2014, 22, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Sendamangalam, V.R.; Gruden, C.L.; Seo, Y. Multiple roles of extracellular polymeric substances on resistance of biofilm and detached clusters. ES&T 2012, 46, 13212–13219. [Google Scholar]
- Del Pozo, J.L. Biofilm-related disease. Expert Rev. Anti-Infect. 2018, 16, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.M.; Islam, M.; Wahab, A.; Billah, M. Antimicrobial resistance crisis and combating approaches. J. Med. 2019, 20, 38–45. [Google Scholar] [CrossRef]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, J.R. Importance of a One Health approach in advancing global health security and the Sustainable Development Goals. Rev. Sci. Tech. Off. Int. Épizoot. 2019, 38, 145–154. [Google Scholar] [CrossRef]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm matrixome: Extracellular components in structured microbial communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- Muhammad, M.H.; Idris, A.L.; Fan, X.; Guo, Y.; Yu, Y.; Jin, X.; Qiu, J.; Guan, X.; Huang, T. Beyond risk: Bacterial biofilms and their regulating approaches. Front. Microbiol. 2020, 11, 928. [Google Scholar] [CrossRef] [PubMed]
- Kimkes, T.E.; Heinemann, M. How bacteria recognise and respond to surface contact. FEMS Microbiol. Rev. 2020, 44, 106–122. [Google Scholar] [CrossRef]
- Aparna, M.S.; Yadav, S. Biofilms: Microbes and disease. Braz. J. Infect. Dis. 2008, 12, 526–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, J.K.; Lee, J. Prevalence and antimicrobial-resistance of Pseudomonas aeruginosa in swimming pools and hot tubs. Int. J. Environ. Res. Public Health 2011, 8, 554–564. [Google Scholar] [CrossRef] [Green Version]
- Beloin, C.; Houry, A.; Froment, M.; Ghigo, J.-M.; Henry, N. A short–time scale colloidal system reveals early bacterial adhesion dynamics. PLoS Biol. 2008, 6, e167. [Google Scholar] [CrossRef] [PubMed]
- Dunne, W.M., Jr. Bacterial adhesion: Seen any good biofilms lately? Clin. Microbiol. Rev. 2002, 15, 155–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting microbial biofilms: Current and prospective therapeutic strategies. Nat. Rev. Microbiol. 2017, 15, 740–755. [Google Scholar] [CrossRef]
- Kokare, C.; Chakraborty, S.; Khopade, A.; Mahadik, K.R. Biofilm: Importance and applications. Indian J. Biotechnol. 2009, 8, 159–168. [Google Scholar]
- Sauer, K.; Camper, A.K.; Ehrlich, G.D.; Costerton, J.W.; Davies, D.G. Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm. J. Bacteriol. 2002, 184, 1140–1154. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.K.; Collins, J.J. Dispersing biofilms with engineered enzymatic bacteriophage. Proc. Natl. Acad. Sci. USA 2007, 104, 11197–11202. [Google Scholar] [CrossRef] [Green Version]
- Evans, L.V. Biofilms: Recent Advances in Their Study and Control, 1st ed.; CRC Press: London, UK, 2000. [Google Scholar]
- Flemming, H.-C.; Wingender, J.; Griegbe, T.; Mayer, C. Physico-chemical properties of biofilms. In Biofilms: Recent Advances in Their Study and Control; Harwood Academic Publishers: Amsterdam, The Netherlands, 2000; pp. 19–34. [Google Scholar]
- Flemming, H.; Wingender, J. The biofilm matrix. Nat. Publ. Gr. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Lembre, P.; Lorentz, C.; Di Martino, P. Exopolysaccharides of the biofilm matrix: A complex biophysical world. In The Complex World of Polysaccharides; Karunaratne, D., Ed.; IntechOpen: London, UK, 2012. [Google Scholar]
- Limoli, D.H.; Jones, C.J.; Wozniak, D.J. Bacterial extracellular polysaccharides in biofilm formation and function. Microbiol Spectr. 2015, 3, 10.1128. [Google Scholar] [CrossRef] [PubMed]
- Rehm, B.H. Bacterial polymers: Biosynthesis, modifications and applications. Nat. Rev. Microbiol. 2010, 8, 578–592. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.B.; Velliyagounder, K.; Ragunath, C.; Rohde, H.; Mack, D.; Knobloch, J.K.-M.; Ramasubbu, N. Genes involved in the synthesis and degradation of matrix polysaccharide in Actinobacillus actinomycetemcomitans and Actinobacillus pleuropneumoniae biofilms. J. Bacteriol. 2004, 186, 8213–8220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevenson, G.; Andrianopoulos, K.; Hobbs, M.; Reeves, P.R. Organization of the Escherichia coli K-12 gene cluster responsible for production of the extracellular polysaccharide colanic acid. J. Bacteriol. 1996, 178, 4885–4893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayem, S.; Manzo, E.; Ciavatta, L.; Tramice, A.; Cordone, A.; Zanfardino, A.; De Felice, M.; Varcamonti, M. Anti-biofilm activity of an exopolysaccharide from a sponge-associated strain of Bacillus licheniformis. Microb. Cell Factories 2011, 10, 74. [Google Scholar] [CrossRef] [Green Version]
- Wingender, J.; Strathmann, M.; Rode, A.; Leis, A.; Flemming, H.-C. [25] Isolation and biochemical characterization of extracellular polymeric substances from Pseudomonas aeruginosa. Meth Enzymol. 2001, 336, 302–314. [Google Scholar]
- Ma, L.; Jackson, K.D.; Landry, R.M.; Parsek, M.R.; Wozniak, D.J. Analysis of Pseudomonas aeruginosa conditional psl variants reveals roles for the psl polysaccharide in adhesion and maintaining biofilm structure postattachment. J. Bacteriol. 2006, 188, 8213–8221. [Google Scholar] [CrossRef] [Green Version]
- Zogaj, X.; Nimtz, M.; Rohde, M.; Bokranz, W.; Römling, U. The multicellular morphotypes of Salmonella Typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol. Microbiol. 2001, 39, 1452–1463. [Google Scholar] [CrossRef]
- Conrad, A.; Kontro, M.; Keinänen, M.M.; Cadoret, A.; Faure, P.; Mansuy-Huault, L.; Block, J.-C. Fatty acids of lipid fractions in extracellular polymeric substances of activated sludge flocs. Lipids 2003, 38, 1093–1105. [Google Scholar] [CrossRef]
- Fong, J.; Yildiz, F. Biofilm matrix proteins. Microbiol. Spectr. 2015, 3, 10.1128. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, J.B.; Ragunath, C.; Ramasubbu, N.; Fine, D.H. Detachment of Actinobacillus actinomycetemcomitans biofilm cells by an endogenous β-hexosaminidase activity. J. Bacteriol. 2003, 185, 4693–4698. [Google Scholar] [CrossRef] [Green Version]
- Martí, M.; Trotonda, M.P.; Tormo-Más, M.Á.; Vergara-Irigaray, M.; Cheung, A.L.; Lasa, I.; Penadés, J.R. Extracellular proteases inhibit protein-dependent biofilm formation in Staphylococcus aureus. Microbes Infect. 2010, 12, 55–64. [Google Scholar] [CrossRef]
- Nijland, R.; Hall, M.J.; Burgess, J.G. Dispersal of biofilms by secreted, matrix degrading, bacterial DNase. PLoS ONE 2010, 5, e15668. [Google Scholar] [CrossRef] [Green Version]
- Toyofuku, M.; Roschitzki, B.; Riedel, K.; Eberl, L. Identification of proteins associated with the Pseudomonas aeruginosa biofilm extracellular matrix. J. Proteome Res. 2012, 11, 4906–4915. [Google Scholar] [CrossRef]
- Wingender, J.; Jaeger, K.-E.; Flemming, H.-C. Interaction between extracellular polysaccharides and enzymes. In Microbial Extracellular Polymeric Substances; Springer: Berlin/Heidelberg, Germany, 1999; pp. 231–251. [Google Scholar]
- Izano, E.A.; Amarante, M.A.; Kher, W.B.; Kaplan, J.B. Differential roles of poly-N-acetylglucosamine surface polysaccharide and extracellular DNA in Staphylococcus aureus and Staphylococcus epidermidis biofilms. Appl. Environ. Microbiol. 2008, 74, 470–476. [Google Scholar] [CrossRef] [Green Version]
- Wingender, J.; Neu, T.R.; Flemming, H.C. What are bacterial extracellular polymeric substances? In Microbial Extracellular Polymeric Substances; Springer: Berlin/Heidelberg, Germany, 1999; pp. 1–19. [Google Scholar]
- Molin, S.; Tolker-Nielsen, T. Gene transfer occurs with enhanced efficiency in biofilms and induces enhanced stabilisation of the biofilm structure. Curr. Opin. Biotechnol. 2003, 14, 255–261. [Google Scholar] [CrossRef]
- Yang, L.; Barken, K.B.; Skindersoe, M.E.; Christensen, A.B.; Givskov, M.; Tolker-Nielsen, T. Effects of iron on DNA release and biofilm development by Pseudomonas aeruginosa. Microbiology 2007, 153, 1318–1328. [Google Scholar] [CrossRef] [Green Version]
- Whitchurch, C.B.; Tolker-Nielsen, T.; Ragas, P.C.; Mattick, J.S. Extracellular DNA required for bacterial biofilm formation. Science 2002, 295, 1487. [Google Scholar] [CrossRef]
- Vilain, S.; Pretorius, J.M.; Theron, J.; Brözel, V.S. DNA as an adhesin: Bacillus cereus requires extracellular DNA to form biofilms. Appl. Environ. Microbiol. 2009, 75, 2861–2868. [Google Scholar] [CrossRef] [Green Version]
- Okshevsky, M.; Meyer, R.L. The role of extracellular DNA in the establishment, maintenance and perpetuation of bacterial biofilms. Crit. Rev. Microbiol. 2015, 41, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, M.; Lappann, M.; Knøchel, S.; Molin, S. Role of extracellular DNA during biofilm formation by Listeria monocytogenes. Appl. Environ. Microbiol. 2010, 76, 2271–2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilton, M.; Charron-Mazenod, L.; Moore, R.; Lewenza, S. Extracellular DNA acidifies biofilms and induces aminoglycoside resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2016, 60, 544–553. [Google Scholar] [CrossRef]
- Neu, T.R.; Poralla, K. An amphiphilic polysaccharide from an adhesive Rhodococcus strain. FEMS Microbiol. Lett. 1988, 49, 389–392. [Google Scholar] [CrossRef]
- Kjelleberg, S.; Givskov, M. Biofilm Mode of Life; Horizon Bioscience; The University of New South Wales: Sydney, Australia; Technical University of Denmark: Lyngby, Denmark, 2007. [Google Scholar]
- Sutherland, I.W. The biofilm matrix–an immobilized but dynamic microbial environment. Trends Microbiol. 2001, 9, 222–227. [Google Scholar] [CrossRef]
- Schmitt, J.; Flemming, H.-C. Water binding in biofilms. Water Sci. Technol. 1999, 39, 77–82. [Google Scholar] [CrossRef]
- Lewis, K. Riddle of biofilm resistance. Antimicrob. Agents Chemother. 2001, 45, 999–1007. [Google Scholar] [CrossRef] [Green Version]
- Rani, S.A.; Pitts, B.; Beyenal, H.; Veluchamy, R.A.; Lewandowski, Z.; Davison, W.M.; Buckingham-Meyer, K.; Stewart, P.S. Spatial patterns of DNA replication, protein synthesis, and oxygen concentration within bacterial biofilms reveal diverse physiological states. J. Bacteriol. 2007, 189, 4223–4233. [Google Scholar] [CrossRef] [Green Version]
- Bridier, A.; Dubois-Brissonnet, F.; Boubetra, A.; Thomas, V.; Briandet, R. The biofilm architecture of sixty opportunistic pathogens deciphered using a high throughput CLSM method. J. Microbiol. Methods 2010, 82, 64–70. [Google Scholar] [CrossRef]
- Lear, G.; Lewis, G.D. Microbial Biofilms: Current Research and Applications; Lincoln University: Christchurch, New Zealand; University of Auckland: Auckland, New Zealand, 2012. [Google Scholar]
- Toyofuku, M.; Inaba, T.; Kiyokawa, T.; Obana, N.; Yawata, Y.; Nomura, N. Environmental factors that shape biofilm formation. Biosci. Biotechnol. Biochem. 2016, 80, 7–12. [Google Scholar] [CrossRef]
- Tielen, P.; Strathmann, M.; Jaeger, K.-E.; Flemming, H.-C.; Wingender, J. Alginate acetylation influences initial surface colonization by mucoid Pseudomonas aeruginosa. Microbiol. Res. 2005, 160, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Watnick, P.I.; Kolter, R. Steps in the development of a Vibrio cholerae El Tor biofilm. Mol. Microbiol. 1999, 34, 586–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danese, P.N.; Pratt, L.A.; Kolter, R. Exopolysaccharide production is required for development of Escherichia coli K-12 biofilm architecture. J. Bacteriol. 2000, 182, 3593–3596. [Google Scholar] [CrossRef] [PubMed]
- Branda, S.S.; Chu, F.; Kearns, D.B.; Losick, R.; Kolter, R. A major protein component of the Bacillus subtilis biofilm matrix. Mol. Microbiol. 2006, 59, 1229–1238. [Google Scholar] [CrossRef]
- Manfiolli, A.O.; Dos Reis, T.F.; de Assis, L.J.; de Castro, P.A.; Silva, L.P.; Hori, J.I.; Walker, L.A.; Munro, C.A.; Rajendran, R.; Ramage, G. Mitogen activated protein kinases (MAPK) and protein phosphatases are involved in Aspergillus fumigatus adhesion and biofilm formation. Cell Surf. 2018, 1, 43–56. [Google Scholar] [CrossRef]
- McCourt, J.; O′Halloran, D.P.; McCarthy, H.; O′Gara, J.P.; Geoghegan, J.A. Fibronectin-binding proteins are required for biofilm formation by community-associated methicillin-resistant Staphylococcus aureus strain LAC. FEMS Microbiol. Lett. 2014, 353, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Ling, N.; Gao, J.; Zhang, X.; Zhang, M.; Tong, L.; Zeng, H.; Zhang, J.; Wu, Q. Roles of outer membrane protein W (OmpW) on survival, morphology, and biofilm formation under NaCl stresses in Cronobacter sakazakii. Int. J. Dairy Sci. 2018, 101, 3844–3850. [Google Scholar] [CrossRef]
- Liang, X.; Chen, Y.-Y.M.; Ruiz, T.; Wu, H. New cell surface protein involved in biofilm formation by Streptococcus parasanguinis. Infect. Immun. 2011, 79, 3239–3248. [Google Scholar] [CrossRef] [Green Version]
- Camilli, A.; Bassler, B.L. Bacterial small-molecule signaling pathways. Science 2006, 311, 1113–1116. [Google Scholar] [CrossRef] [Green Version]
- Kalia, V.C. Quorum sensing inhibitors: An overview. Biotechnol. Adv. 2013, 31, 224–245. [Google Scholar] [CrossRef]
- Xue, T.; Ni, J.; Shang, F.; Chen, X.; Zhang, M. Autoinducer-2 increases biofilm formation via an ica-and bhp-dependent manner in Staphylococcus epidermidis RP62A. Microbes Infect. 2015, 17, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Yoon, S.S. Pseudomonas aeruginosa biofilm, a programmed bacterial life for fitness. J. Microbiol. Biotechnol. 2017, 27, 1053–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikkelsen, H.; Sivaneson, M.; Filloux, A. Key two-component regulatory systems that control biofilm formation in Pseudomonas aeruginosa. Environ. Microbiol. 2011, 13, 1666–1681. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Bassler, B.L. Quorum-sensing non-coding small RNAs use unique pairing regions to differentially control mRNA targets. Mol. Microbiol. 2012, 83, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Moreno, R.; Fonseca, P.; Rojo, F. Two small RNAs, CrcY and CrcZ, act in concert to sequester the Crc global regulator in Pseudomonas putida, modulating catabolite repression. Mol. Microbiol. 2012, 83, 24–40. [Google Scholar] [CrossRef]
- Jørgensen, M.G.; Nielsen, J.S.; Boysen, A.; Franch, T.; Møller-Jensen, J.; Valentin-Hansen, P. Small regulatory RNAs control the multi-cellular adhesive lifestyle of Escherichia coli. Mol. Microbiol. 2012, 84, 36–50. [Google Scholar] [CrossRef] [Green Version]
- Thomason, M.K.; Fontaine, F.; De Lay, N.; Storz, G. A small RNA that regulates motility and biofilm formation in response to changes in nutrient availability in Escherichia coli. Mol. Microbiol. 2012, 84, 17–35. [Google Scholar] [CrossRef]
- Holmqvist, E.; Reimegård, J.; Sterk, M.; Grantcharova, N.; Römling, U.; Wagner, E.G.H. Two antisense RNAs target the transcriptional regulator CsgD to inhibit curli synthesis. EMBO J. 2010, 29, 1840–1850. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, C.; Papenfort, K.; Hentrich, K.; Ahmad, I.; Le Guyon, S.; Reimann, R.; Grantcharova, N.; Römling, U. Hfq and Hfq-dependent small RNAs are major contributors to multicellular development in Salmonella enterica serovar Typhimurium. RNA Biol. 2012, 9, 489–502. [Google Scholar] [CrossRef] [Green Version]
- Mika, F.; Busse, S.; Possling, A.; Berkholz, J.; Tschowri, N.; Sommerfeldt, N.; Pruteanu, M.; Hengge, R. Targeting of csgD by the small regulatory RNA RprA links stationary phase, biofilm formation and cell envelope stress in Escherichia coli. Mol. Microbiol. 2012, 84, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Papenfort, K.; Said, N.; Welsink, T.; Lucchini, S.; Hinton, J.C.; Vogel, J. Specific and pleiotropic patterns of mRNA regulation by ArcZ, a conserved, Hfq-dependent small RNA. Mol. Microbiol. 2009, 74, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Mandin, P.; Gottesman, S. Integrating anaerobic/aerobic sensing and the general stress response through the ArcZ small RNA. EMBO J. 2010, 29, 3094–3107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, D.W.; Suzuki, K.; Oakford, L.; Simecka, J.W.; Hart, M.E.; Romeo, T. Biofilm formation and dispersal under the influence of the global regulator CsrA of Escherichia coli. J. Bacteriol. 2002, 184, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Heroven, A.K.; Böhme, K.; Rohde, M.; Dersch, P. A Csr-type regulatory system, including small non-coding RNAs, regulates the global virulence regulator RovA of Yersinia pseudotuberculosis through RovM. Mol. Microbiol. 2008, 68, 1179–1195. [Google Scholar] [CrossRef] [PubMed]
- Sonnleitner, E.; Gonzalez, N.; Sorger-Domenigg, T.; Heeb, S.; Richter, A.S.; Backofen, R.; Williams, P.; Hüttenhofer, A.; Haas, D.; Bläsi, U. The small RNA PhrS stimulates synthesis of the Pseudomonas aeruginosa quinolone signal. Mol. Microbiol. 2011, 80, 868–885. [Google Scholar] [CrossRef] [PubMed]
- Majdalani, N.; Chen, S.; Murrow, J.; St John, K.; Gottesman, S. Regulation of RpoS by a novel small RNA: The characterization of RprA. Mol. Microbiol. 2001, 39, 1382–1394. [Google Scholar] [CrossRef]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The microbial “protective clothing” in extreme environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef] [Green Version]
- Hathroubi, S.; Mekni, M.A.; Domenico, P.; Nguyen, D.; Jacques, M. Biofilms: Microbial shelters against antibiotics. Microb. Drug Resist. 2017, 23, 147–156. [Google Scholar] [CrossRef]
- Tseng, B.S.; Zhang, W.; Harrison, J.J.; Quach, T.P.; Song, J.L.; Penterman, J.; Singh, P.K.; Chopp, D.L.; Packman, A.I.; Parsek, M.R. The extracellular matrix protects Pseudomonas aeruginosa biofilms by limiting the penetration of tobramycin. Environ. Microbiol. 2013, 15, 2865–2878. [Google Scholar]
- Zhou, G.; Shi, Q.-S.; Huang, X.-M.; Xie, X.-B. The three bacterial lines of defense against antimicrobial agents. Int. J. Mol. Sci. 2015, 16, 21711–21733. [Google Scholar] [CrossRef] [Green Version]
- Stewart, P.S.; Zhang, T.; Xu, R.; Pitts, B.; Walters, M.C.; Roe, F.; Kikhney, J.; Moter, A. Reaction–diffusion theory explains hypoxia and heterogeneous growth within microbial biofilms associated with chronic infections. NPJ Biofilms Microbiomes 2016, 2, 16012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Y.; Zhou, J.; Chen, C.; Shen, D.; Song, W.; Feng, Y. In vitro adsorption revealing an apparent strong interaction between endophyte Pantoea agglomerans YS19 and host rice. Curr. Microbiol. 2008, 57, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Miao, Y.; Yi, T.; Zhou, J.; Lu, Z.; Feng, Y. SPM43. 1 contributes to acid-resistance of non-symplasmata-forming cells in Pantoea agglomerans YS19. Curr. Microbiol. 2012, 64, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yu, Z.; Ding, T. Quorum-sensing regulation of antimicrobial resistance in bacteria. Microorganisms 2020, 8, 425. [Google Scholar] [CrossRef] [PubMed]
- Passos da Silva, D.; Schofield, M.C.; Parsek, M.R.; Tseng, B.S. An update on the sociomicrobiology of quorum sensing in gram-negative biofilm development. Pathogens 2017, 6, 51. [Google Scholar] [CrossRef]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and promise of bacterial quorum sensing research. Nature 2017, 551, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Wolska, K.I.; Grudniak, A.M.; Rudnicka, Z.; Markowska, K. Genetic control of bacterial biofilms. J. Appl. Genet. 2016, 57, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Pena, R.T.; Blasco, L.; Ambroa, A.; González-Pedrajo, B.; Fernández-García, L.; López, M.; Bleriot, I.; Bou, G.; García-Contreras, R.; Wood, T.K. Relationship between quorum sensing and secretion systems. Front. Microbiol. 2019, 10, 1100. [Google Scholar] [CrossRef] [Green Version]
- Blanco, P.; Hernando-Amado, S.; Reales-Calderon, J.A.; Corona, F.; Lira, F.; Alcalde-Rico, M.; Bernardini, A.; Sanchez, M.B.; Martinez, J.L. Bacterial multidrug efflux pumps: Much more than antibiotic resistance determinants. Microorganisms 2016, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Blöcher, R.; Rodarte Ramírez, A.; Castro-Escarpulli, G.; Curiel-Quesada, E.; Reyes-Arellano, A. Design, synthesis, and evaluation of alkyl-quinoxalin-2 (1 1H)-one derivatives as anti-quorum sensing molecules, inhibiting biofilm formation in Aeromonas caviae Sch3. Molecules 2018, 23, 3075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Schweizer, H.P. Bacterial resistance to antibiotics: Active efflux and reduced uptake. Adv. Drug Deliv. Rev. 2005, 57, 1486–1513. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, S.K.; Chowdhury, I.; Singh, R. Understanding the mechanism of bacterial biofilms resistance to antimicrobial agents. Open J. Med. Microbiol. 2017, 11, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Acker, H.; Coenye, T. The role of efflux and physiological adaptation in biofilm tolerance and resistance. J. Biol. Chem. 2016, 291, 12565–12572. [Google Scholar] [CrossRef]
- Kunz Coyne, A.J.; El Ghali, A.; Holger, D.; Rebold, N.; Rybak, M.J. Therapeutic strategies for emerging multidrug-resistant Pseudomonas aeruginosa. Infect Dis Ther. 2022, 11, 661–682. [Google Scholar] [CrossRef]
- Zhang, L.; Mah, T.-F. Involvement of a novel efflux system in biofilm-specific resistance to antibiotics. J. Bacteriol. 2008, 190, 4447–4452. [Google Scholar] [CrossRef] [Green Version]
- Buroni, S.; Matthijs, N.; Spadaro, F.; Van Acker, H.; Scoffone, V.C.; Pasca, M.R.; Riccardi, G.; Coenye, T. Differential roles of RND efflux pumps in antimicrobial drug resistance of sessile and planktonic Burkholderia cenocepacia cells. Antimicrob. Agents Chemother. 2014, 58, 7424–7429. [Google Scholar] [CrossRef] [Green Version]
- Shoji, M.M.; Chen, A.F. Biofilms in periprosthetic joint infections: A review of diagnostic modalities, current treatments, and future directions. J. Knee Surg. 2020, 33, 119–131. [Google Scholar] [CrossRef]
- Furner-Pardoe, J.; Anonye, B.O.; Cain, R.; Moat, J.; Ortori, C.A.; Lee, C.; Barrett, D.A.; Corre, C.; Harrison, F. Anti-biofilm efficacy of a medieval treatment for bacterial infection requires the combination of multiple ingredients. Sci. Rep. 2020, 10, 12687. [Google Scholar] [CrossRef]
- Vajjala, A.; Biswas, D.; Tay, W.H.; Hanski, E.; Kline, K.A. Streptolysin-induced endoplasmic reticulum stress promotes group A streptococcal host-associated biofilm formation and necrotising fasciitis. Cell. Microbiol. 2019, 21, e12956. [Google Scholar] [CrossRef] [Green Version]
- Colombo, A.P.V.; Magalhães, C.B.; Hartenbach, F.A.R.R.; do Souto, R.M.; da Silva-Boghossian, C.M. Periodontal-disease-associated biofilm: A reservoir for pathogens of medical importance. Microb. Pathog. 2016, 94, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Roldán, S.; Herrera, D.; Sanz, M. Biofilms and the tongue: Therapeutical approaches for the control of halitosis. Clin. Oral Investig. 2003, 7, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Zimmerli, W.; Sendi, P. Orthopaedic biofilm infections. APMIS 2017, 125, 353–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakar, M.B.A.; McKimm, J.; Haque, M. Otitis media and biofilm: An overview. Int. J. Nutr. Pharmacol. Neurol. 2018, 8, 70–78. [Google Scholar]
- Høiby, N.; Ciofu, O.; Bjarnsholt, T. Pseudomonas aeruginosa biofilms in cystic fibrosis. Future Microbiol. 2010, 5, 1663–1674. [Google Scholar] [CrossRef]
- Galie, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef]
- Lindsay, D.; von Holy, A. What food safety professionals should know about bacterial biofilms. Brit. Food J. 2006, 108, 27–37. [Google Scholar] [CrossRef]
- Srey, S.; Jahid, I.K.; Ha, S.-D. Biofilm formation in food industries: A food safety concern. Food Control 2013, 31, 572–585. [Google Scholar] [CrossRef]
- Govaert, M.; Smet, C.; Baka, M.; Janssens, T.; Impe, J.V. Influence of incubation conditions on the formation of model biofilms by Listeria monocytogenes and Salmonella Typhimurium on abiotic surfaces. J. Appl. Microbiol. 2018, 125, 1890–1900. [Google Scholar] [CrossRef]
- Tang, L.; Pillai, S.; Revsbech, N.P.; Schramm, A.; Bischoff, C.; Meyer, R.L. Biofilm retention on surfaces with variable roughness and hydrophobicity. Biofouling 2011, 27, 111–121. [Google Scholar] [CrossRef]
- Makovcova, J.; Babak, V.; Kulich, P.; Masek, J.; Slany, M.; Cincarova, L. Dynamics of mono-and dual-species biofilm formation and interactions between Staphylococcus aureus and Gram-negative bacteria. Microb. Biotechnol. 2017, 10, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Van Houdt, R.; Michiels, C. Biofilm formation and the food industry, a focus on the bacterial outer surface. J. Appl. Microbiol. 2010, 109, 1117–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, E.A.; de Andrade, N.J.; da Silva, L.H.M.; de Carvalho, A.F.; de Sá Silva, C.A.; Ramos, A.M. Control of microbial adhesion as a strategy for food and bioprocess technology. Food Bioproc. Tech. 2010, 3, 321–332. [Google Scholar] [CrossRef]
- Dhowlaghar, N.; Bansal, M.; Schilling, M.W.; Nannapaneni, R. Scanning electron microscopy of Salmonella biofilms on various food-contact surfaces in catfish mucus. Food Microbiol. 2018, 74, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Jindal, S.; Anand, S.; Metzger, L.; Amamcharla, J. A comparison of biofilm development on stainless steel and modified-surface plate heat exchangers during a 17-h milk pasteurization run. Int. J. Dairy Sci. 2018, 101, 2921–2926. [Google Scholar] [CrossRef]
- Dutra, T.V.; Fernandes, M.d.S.; Perdoncini, M.R.F.G.; Anjos, M.M.d.; Abreu Filho, B.A.d. Capacity of Escherichia coli and Staphylococcus aureus to produce biofilm on stainless steel surfaces in the presence of food residues. J. Food Process. Preserv. 2018, 42, e13574. [Google Scholar] [CrossRef]
- Iñiguez-Moreno, M.; Gutiérrez-Lomelí, M.; Avila-Novoa, M.G. Kinetics of biofilm formation by pathogenic and spoilage microorganisms under conditions that mimic the poultry, meat, and egg processing industries. Int. J. Food Microbiol. 2019, 303, 32–41. [Google Scholar] [CrossRef]
- Karaca, B.; Buzrul, S.; Coleri Cihan, A. Anoxybacillus and Geobacillus biofilms in the dairy industry: Effects of surface material, incubation temperature and milk type. Biofouling 2019, 35, 551–560. [Google Scholar] [CrossRef]
- Grigore-Gurgu, L.; Bucur, F.I.; Borda, D.; Alexa, E.-A.; Neagu, C.; Nicolau, A.I. Biofilms formed by pathogens in food and food processing environments. Bacterial Biofilms; Dincer, S., Özdenefe, M., Arkut, A., Eds.; Intech Open: London, UK, 2019; pp. 1–32. [Google Scholar]
- Kwon, M.; Hussain, M.S.; Oh, D.H. Biofilm formation of Bacillus cereus under food-processing-related conditions. Food Sci. Biotechnol. 2017, 26, 1103–1111. [Google Scholar] [CrossRef]
- Klančnik, A.; Šimunović, K.; Sterniša, M.; Ramić, D.; Smole Možina, S.; Bucar, F. Anti-adhesion activity of phytochemicals to prevent Campylobacter jejuni biofilm formation on abiotic surfaces. Phytochem. Rev. 2021, 20, 55–84. [Google Scholar] [CrossRef] [Green Version]
- Carrascosa, C.; Raheem, D.; Ramos, F.; Saraiva, A.; Raposo, A. Microbial biofilms in the food industry—A comprehensive review. Int. J. Environ. Res. Public Health. 2021, 18, 2014. [Google Scholar] [CrossRef]
- Ricci, E.; Schwinghamer, T.; Fan, D.; Smith, D.L.; Gravel, V. Growth promotion of greenhouse tomatoes with Pseudomonas sp. and Bacillus sp. biofilms and planktonic cells. Appl. Soil Ecol. 2019, 138, 61–68. [Google Scholar] [CrossRef]
- Kadariya, J.; Smith, T.C.; Thapaliya, D. Staphylococcus aureus and staphylococcal food-borne disease: An ongoing challenge in public health. Biomed Res. Int. 2014, 2014, 827965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giaouris, E.; Heir, E.; Desvaux, M.; Hébraud, M.; Møretrø, T.; Langsrud, S.; Doulgeraki, A.; Nychas, G.-J.; Kačániová, M.; Czaczyk, K. Intra-and inter-species interactions within biofilms of important foodborne bacterial pathogens. Front. Microbiol. 2015, 6, 841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.I.; Mizan, M.F.R.; Ashrafudoulla, M.; Nahar, S.; Joo, H.-J.; Jahid, I.K.; Park, S.H.; Kim, K.-S.; Ha, S.-D. Inhibitory effects of probiotic potential lactic acid bacteria isolated from kimchi against Listeria monocytogenes biofilm on lettuce, stainless-steel surfaces, and MBEC™ biofilm device. LWT 2020, 118, 108864. [Google Scholar] [CrossRef]
- Rodríguez-Saavedra, M.; de Llano, D.G.; Beltran, G.; Torija, M.-J.; Moreno-Arribas, M.V. Pectinatus spp.- unpleasant and recurrent brewing spoilage bacteria. Int. J. Food Microbiol. 2021, 336, 108900. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Tau, N.P.; Smouse, S.L.; Allam, M.; Ismail, A.; Ramalwa, N.R.; Disenyeng, B.; Ngomane, M.; Thomas, J. Outbreak of Listeria monocytogenes in South Africa, 2017–2018: Laboratory activities and experiences associated with whole-genome sequencing analysis of isolates. Foodborne Pathog. Dis. 2019, 16, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, A.; Shantikumar, S.; Beaufoy, D.; Allman, A.; Fenelon, D.; Reynolds, K.; Normington, A.; Afza, M.; Todkill, D. Outbreak of Clostridium perfringens food poisoning linked to leeks in cheese sauce: An unusual source. Epidemiol. Infect. 2020, 148, e43. [Google Scholar] [CrossRef] [Green Version]
- Mikhail, A.; Jenkins, C.; Dallman, T.; Inns, T.; Douglas, A.; Martín, A.; Fox, A.; Cleary, P.; Elson, R.; Hawker, J. An outbreak of Shiga toxin-producing Escherichia coli O157: H7 associated with contaminated salad leaves: Epidemiological, genomic and food trace back investigations. Epidemiol. Infect. 2018, 146, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Vaughn, E.L.; Vo, Q.T.; Vostok, J.; Stiles, T.; Lang, A.; Brown, C.M.; Klevens, R.M.; Madoff, L. Linking epidemiology and whole-genome sequencing to investigate Salmonella outbreak, Massachusetts, USA, 2018. Emerg. Infect. Dis. 2020, 26, 1538. [Google Scholar] [CrossRef]
- Kenyon, J.; Inns, T.; Aird, H.; Swift, C.; Astbury, J.; Forester, E.; Decraene, V. Campylobacter outbreak associated with raw drinking milk, North West England, 2016. Epidemiol. Infect. 2020, 148, e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denayer, S.; Delbrassinne, L.; Nia, Y.; Botteldoorn, N. Food-borne outbreak investigation and molecular typing: High diversity of Staphylococcus aureus strains and importance of toxin detection. Toxins 2017, 9, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Zhang, R.; Qi, X.; Zhou, B.; Wang, J.; Chen, Y.; Zhang, H. Epidemiology of foodborne disease outbreaks caused by Vibrio parahaemolyticus during 2010–2014 in Zhejiang Province, China. Food Control 2017, 77, 110–115. [Google Scholar] [CrossRef]
- Griffiths, M.; Schraft, H. Bacillus cereus food poisoning. In Foodborne diseases; Elsevier: Amsterdam, The Netherlands, 2017; pp. 395–405. [Google Scholar]
- Wu, Y.; Wen, J.; Ma, Y.; Ma, X.; Chen, Y. Epidemiology of foodborne disease outbreaks caused by Vibrio parahaemolyticus, China, 2003–2008. Food Control 2014, 46, 197–202. [Google Scholar] [CrossRef]
- Popovic, I.; Heron, B.; Covacin, C. Listeria: An Australian perspective (2001–2010). Foodborne Pathog. Dis. 2014, 11, 425–432. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Jensen, P.Ø.; Rasmussen, T.B.; Christophersen, L.; Calum, H.; Hentzer, M.; Hougen, H.-P.; Rygaard, J.; Moser, C.; Eberl, L. Garlic blocks quorum sensing and promotes rapid clearing of pulmonary Pseudomonas aeruginosa infections. Microbiology 2005, 151, 3873–3880. [Google Scholar] [CrossRef] [Green Version]
- Persson, T.; Hansen, T.H.; Rasmussen, T.B.; Skindersø, M.E.; Givskov, M.; Nielsen, J. Rational design and synthesis of new quorum-sensing inhibitors derived from acylated homoserine lactones and natural products from garlic. Org. Biomol. Chem. 2005, 3, 253–262. [Google Scholar] [CrossRef]
- Kazemian, H.; Ghafourian, S.; Heidari, H.; Amiri, P.; Yamchi, J.K.; Shavalipour, A.; Houri, H.; Maleki, A.; Sadeghifard, N. Antibacterial, anti-swarming and anti-biofilm formation activities of Chamaemelum nobile against Pseudomonas aeruginosa. Rev. Soc. Bras. Med. Trop. 2015, 48, 432–436. [Google Scholar] [CrossRef] [Green Version]
- Howell, A.B. Cranberry proanthocyanidins and the maintenance of urinary tract health. Crit. Rev. Food Sci. Nutr. 2002, 42, 273–278. [Google Scholar] [CrossRef]
- Burger, O.; Ofek, I.; Tabak, M.; Weiss, E.I.; Sharon, N.; Neeman, I. A high molecular mass constituent of cranberry juice inhibits Helicobacter pylori adhesion to human gastric mucus. FEMS Microbiol. Immunol. 2000, 29, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, A.; Kimizuka, R.; Kato, T.; Okuda, K. Inhibitory effects of cranberry juice on attachment of oral streptococci and biofilm formation. Oral Microbiol. Immunol. 2004, 19, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, D.; Feldman, M.; Ofek, I.; Weiss, E.I. Cranberry high molecular weight constituents promote Streptococcus sobrinus desorption from artificial biofilm. Int. J. Antimicrob. Agents 2005, 25, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-W.; Chang, I.-M.; Oh, K.-B. Inhibition of the bacterial surface protein anchoring transpeptidase sortase by medicinal plants. Biosci. Biotechnol. Biochem. 2002, 66, 2751–2754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, L.; Zou, W.; Zheng, K.; Shi, B.; Liu, M. The Herba patriniae (Caprifoliaceae): A review on traditional uses, phytochemistry, pharmacology and quality control. J. Ethnopharmacol. 2021, 265, 113264. [Google Scholar] [CrossRef] [PubMed]
- Gerstmeier, J.; Seegers, J.; Witt, F.; Waltenberger, B.; Temml, V.; Rollinger, J.M.; Stuppner, H.; Koeberle, A.; Schuster, D.; Werz, O. Ginkgolic acid is a multi-target inhibitor of key enzymes in pro-inflammatory lipid mediator biosynthesis. Front. Pharmacol. 2019, 10, 797. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kim, Y.-G.; Ryu, S.Y.; Cho, M.H.; Lee, J. Ginkgolic acids and Ginkgo biloba extract inhibit Escherichia coli O157: H7 and Staphylococcus aureus biofilm formation. Int. J. Food Microbiol. 2014, 174, 47–55. [Google Scholar] [CrossRef]
- Elekhnawy, E.; Negm, W.A.; El-Aasr, M.; Kamer, A.A.; Alqarni, M.; Batiha, G.E.-S.; Obaidullah, A.J.; Fawzy, H.M. Histological assessment, anti-quorum sensing, and anti-biofilm activities of Dioon spinulosum extract: In vitro and in vivo approach. Sci. Rep. 2022, 12, 180. [Google Scholar] [CrossRef]
- Obaid, R.F.; Kadhim Hindi, N.K.; Kadhum, S.A.; Jafaar Alwaeli, L.A.; Jalil, A.T. Antibacterial activity, anti-adherence and anti-biofilm activities of plants extracts against Aggregatibacter actinomycetemcomitans: An in vitro study in Hilla City, Iraq. Casp. J. Environ. Sci. 2022, 20, 367–372. [Google Scholar]
- Negm, W.A.; El-Aasr, M.; Attia, G.; Alqahtani, M.J.; Yassien, R.I.; Abo Kamer, A.; Elekhnawy, E. Promising antifungal activity of Encephalartos laurentianus de wild against Candida albicans clinical isolates: In vitro and in vivo effects on renal cortex of adult albino rats. J. Fungi 2022, 8, 426. [Google Scholar] [CrossRef]
- Olawuwo, O.S.; Famuyide, I.M.; McGaw, L.J. Antibacterial and antibiofilm activity of selected medicinal plant leaf extracts against pathogens implicated in poultry diseases. Front. Vet. Sci. 2022, 9, 820304. [Google Scholar] [CrossRef]
- Fathi, M.; Ghane, M.; Pishkar, L. Phytochemical composition, antibacterial, and antibiofilm activity of Malva sylvestris against human pathogenic bacteria. Jundishapur J. Nat. Pharm. Prod. 2022, 17, e114164. [Google Scholar] [CrossRef]
- Priyanto, J.A.; Prastya, M.E.; Sinarawadi, G.S.; Datu’salamah, W.; Avelina, T.Y.; Yanuar, A.I.A.; Azizah, E.; Tachrim, Z.P.; Mozef, T. The antibacterial and antibiofilm potential of Paederia foetida Linn. leaves extract. J. Appl. Pharm. Sci. 2022, 12, 117–124. [Google Scholar] [CrossRef]
- Panjaitan, C.C.; Widyarman, A.S.; Amtha, R.; Astoeti, T.E. Antimicrobial and antibiofilm activity of cinnamon (Cinnamomum burmanii) extract on periodontal pathogens—An in vitro study. Eur. J. Dent. 2022. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Plescia, F.; Venturella, F.; Lauricella, M.; Catania, V.; Polito, G.; Schillaci, D.; Piccionello, A.P.; Giuseppe, D.; D’Anneo, A.; Raffa, D. Chemical composition, cytotoxic effects, antimicrobial and antibiofilm activity of Artemisia arborescens (Vaill.) L. growing wild in the province of Agrigento, Sicily, Italy. Plant Biosyst. 2022, 1–10. [Google Scholar] [CrossRef]
- Rhimi, W.; Mohammed, M.A.; Zarea, A.A.K.; Greco, G.; Tempesta, M.; Otranto, D.; Cafarchia, C. Antifungal, antioxidant and antibiofilm activities of essential oils of Cymbopogon spp. Antibiotics 2022, 11, 829. [Google Scholar] [CrossRef]
- Nazzaro, F.; Polito, F.; Amato, G.; Caputo, L.; Francolino, R.; D’Acierno, A.; Fratianni, F.; Candido, V.; Coppola, R.; De Feo, V. Chemical composition of essential oils of bulbs and aerial parts of two cultivars of Allium sativum and their antibiofilm activity against food and nosocomial pathogens. Antibiotics 2022, 11, 724. [Google Scholar] [CrossRef]
- Gamal El-Din, M.I.; Youssef, F.S.; Altyar, A.E.; Ashour, M.L. GC/MS Analyses of the essential oils obtained from different jatropha species, their discrimination using chemometric analysis and assessment of their antibacterial and anti-biofilm activities. Plants 2022, 11, 1268. [Google Scholar] [CrossRef]
- Djebili, S.; Taş, M.; Bouguerra, A.; Kucukaydin, S.; Ceylan, O.; Duru, M.E.; Barkat, M. Volatile compound profile and essential oil composition of three wild Algerian aromatic plants with their antioxidant and antibiofilm activities. J. Food Meas. Charact. 2022, 16, 987–999. [Google Scholar] [CrossRef]
- De Oliveira Galvão, F.; da Silva Dantas, F.G.; de Lima Santos, C.R.; Marchioro, S.B.; Cardoso, C.A.L.; Wender, H.; Sangalli, A.; de Almeida-Apolonio, A.A.; de Oliveira, K.M.P. Cochlospermum regium (Schrank) pilger leaf extract inhibit methicillin-resistant Staphylococcus aureus biofilm formation. J. Ethnopharmacol. 2020, 261, 113167. [Google Scholar] [CrossRef]
- Vijayakumar, K.; Ramanathan, T. Musa acuminata and its bioactive metabolite 5-Hydroxymethylfurfural mitigates quorum sensing (las and rhl) mediated biofilm and virulence production of nosocomial pathogen Pseudomonas aeruginosa in vitro. J. Ethnopharmacol. 2020, 246, 112242. [Google Scholar] [CrossRef]
- Tang, Y.; Bai, J.; Yang, Y.; Bai, X.; Bello-Onaghise, G.s.; Xu, Y.; Li, Y. Effect of syringopicroside extracted from syringa oblata lindl on the biofilm formation of Streptococcus suis. Molecules 2021, 26, 1295. [Google Scholar] [CrossRef] [PubMed]
- Bocquet, L.; Sahpaz, S.; Bonneau, N.; Beaufay, C.; Mahieux, S.; Samaillie, J.; Roumy, V.; Jacquin, J.; Bordage, S.; Hennebelle, T. Phenolic compounds from Humulus lupulus as natural antimicrobial products: New weapons in the fight against methicillin resistant Staphylococcus aureus, Leishmania mexicana and Trypanosoma brucei strains. Molecules 2019, 24, 1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attallah, N.G.; Al-Fakhrany, O.M.; Elekhnawy, E.; Hussein, I.A.; Shaldam, M.A.; Altwaijry, N.; Alqahtani, M.J.; Negm, W.A. Anti-biofilm and antibacterial activities of Cycas media R. Br secondary metabolites: In silico, in vitro, and in vivo approaches. Antibiotics 2022, 11, 993. [Google Scholar] [CrossRef] [PubMed]
- Viktorová, J.; Stupák, M.; Řehořová, K.; Dobiasová, S.; Hoang, L.; Hajšlová, J.; Van Thanh, T.; Van Tri, L.; Van Tuan, N.; Ruml, T. Lemon grass essential oil does not modulate cancer cells multidrug resistance by citral—Its dominant and strongly antimicrobial compound. Foods 2020, 9, 585. [Google Scholar] [CrossRef]
- Alibi, S.; Selma, W.B.; Ramos-Vivas, J.; Smach, M.A.; Touati, R.; Boukadida, J.; Navas, J.; Mansour, H.B. Anti-oxidant, antibacterial, anti-biofilm, and anti-quorum sensing activities of four essential oils against multidrug-resistant bacterial clinical isolates. Curr. Res. Transl. Med. 2020, 68, 59–66. [Google Scholar] [CrossRef]
- Umamaheswari, A.; SJ, G.F. Investigation on the biofilm eradication potential of selected medicinal plants against methicillin-resistant Staphylococcus aureus. Biotechnol. Rep. 2020, 28, e00523. [Google Scholar]
- Elamary, R.B.; Albarakaty, F.M.; Salem, W.M. Efficacy of Acacia nilotica aqueous extract in treating biofilm-forming and multidrug resistant uropathogens isolated from patients with UTI syndrome. Sci. Rep. 2020, 10, 11125. [Google Scholar] [CrossRef]
- Aumeeruddy-Elalfi, Z.; Ismaël, I.S.; Hosenally, M.; Zengin, G.; Mahomoodally, M.F. Essential oils from tropical medicinal herbs and food plants inhibit biofilm formation in vitro and are non-cytotoxic to human cells. 3 Biotech 2018, 8, 395. [Google Scholar] [CrossRef]
- Bernal-Mercado, A.T.; Vazquez-Armenta, F.J.; Tapia-Rodriguez, M.R.; Islas-Osuna, M.A.; Mata-Haro, V.; Gonzalez-Aguilar, G.A.; Lopez-Zavala, A.A.; Ayala-Zavala, J.F. Comparison of single and combined use of catechin, protocatechuic, and vanillic acids as antioxidant and antibacterial agents against uropathogenic Escherichia coli at planktonic and biofilm levels. Molecules 2018, 23, 2813. [Google Scholar] [CrossRef] [Green Version]
- Dias-Souza, M.V.; Dos Santos, R.M.; Cerávolo, I.P.; Cosenza, G.; Marçal, P.H.F. Euterpe oleracea pulp extract: Chemical analyses, antibiofilm activity against Staphylococcus aureus, cytotoxicity and interference on the activity of antimicrobial drugs. Microb. Pathog. 2018, 114, 29–35. [Google Scholar] [CrossRef]
- Dolatabadi, S.; Nesari, H.; Mahdavi-ourtakand, M. Microbial pathogenesis evaluating the anti-biofilm and antibacterial effects of Juglans regia L. extracts against clinical isolates of Pseudomonas aeruginosa. Microb. Pathog 2018, 118, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Endo, E.H.; Costa, G.M.; Makimori, R.Y.; Ueda-Nakamura, T.; Nakamura, C.V.; Dias Filho, B.P. Anti-biofilm activity of Rosmarinus officinalis, Punica granatum and Tetradenia riparia against methicillin-resistant Staphylococcus aureus (MRSA) and synergic interaction with penicillin. J. Herb. Med. 2018, 14, 48–54. [Google Scholar] [CrossRef]
- Romero, C.M.; Vivacqua, C.G.; Abdulhamid, M.B.; Baigori, M.D.; Slanis, A.C.; Allori, M.C.G.d.; Tereschuk, M.L. Biofilm inhibition activity of traditional medicinal plants from Northwestern Argentina against native pathogen and environmental microorganisms. Rev. Soc. Bras. Med. Trop. 2016, 49, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Cartagena, E.; Colom, O.Á.; Neske, A.; Valdez, J.C.; Bardón, A. Effects of plant lactones on the production of biofilm of Pseudomonas aeruginosa. Chem. Pharm. Bull. 2007, 55, 22–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Combarros-Fuertes, P.; Estevinho, L.M.; Dias, L.G.; Castro, J.M.; Tomás-Barberán, F.A.; Tornadijo, M.E.; Fresno-Baro, J.M. Bioactive components and antioxidant and antibacterial activities of different varieties of honey: A screening prior to clinical application. J. Agric. Food Chem. 2018, 67, 688–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddocks, S.E.; Jenkins, R.E. Honey: A sweet solution to the growing problem of antimicrobial resistance? Future Microbiol. 2013, 8, 1419–1429. [Google Scholar] [CrossRef]
- Cooper, R.; Jenkins, R. Are there feasible prospects for manuka honey as an alternative to conventional antimicrobials? Expert Rev. Anti-Infect. Ther. 2012, 10, 623–625. [Google Scholar] [CrossRef]
- Bouchelaghem, S.; Das, S.; Naorem, R.S.; Czuni, L.; Papp, G.; Kocsis, M. Evaluation of total phenolic and flavonoid contents, antibacterial and antibiofilm activities of Hungarian Propolis ethanolic extract against Staphylococcus aureus. Molecules 2022, 27, 574. [Google Scholar] [CrossRef]
- Alandejani, T.; Marsan, J.; Ferris, W.; Slinger, R.; Chan, F. Effectiveness of honey on Staphylococcus aureus and Pseudomonas aeruginosa biofilms. Otolaryngol. Head Neck Surg. 2009, 141, 114–118. [Google Scholar] [CrossRef]
- Qamar, M.U.; Saleem, S.; Toleman, M.A.; Saqalein, M.; Waseem, M.; Nisar, M.A.; Khurshid, M.; Taj, Z.; Jahan, S. In vitro and in vivo activity of Manuka honey against NDM-1-producing Klebsiella pneumoniae ST11. Future Microbiol. 2018, 13, 13–26. [Google Scholar] [CrossRef]
- Girma, A.; Seo, W.; She, R.C. Antibacterial activity of varying UMF-graded Manuka honeys. PLoS ONE 2019, 14, e0224495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah Pratibha, J.; Williamson Manita, T. Antibacterial activity of honey against ESBL producing Klebsiella pneumoniae from burn wound infections. Int. J. Curr. Pharm. Res. 2015, 7, 32–36. [Google Scholar]
- Idris, A.R.; Afegbua, S.L. Single and joint antibacterial activity of aqueous garlic extract and Manuka honey on extended-spectrum beta-lactamase-producing Escherichia coli. Trans. R. Soc. Trop. Med. Hyg. 2017, 111, 472–478. [Google Scholar] [CrossRef]
- Hillitt, K.; Jenkins, R.; Spiller, O.B.; Beeton, M.L. Antimicrobial activity of Manuka honey against antibiotic-resistant strains of the cell wall-free bacteria Ureaplasma parvum and Ureaplasma urealyticum. Lett. Appl. Microbiol. 2017, 64, 198–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadl, A.E.-W. Antibacterial and antibiofilm effects of bee venom from (Apis mellifera) on multidrug-resistant bacteria (MDRB). Al-Azhar J. Pharm. Sci. 2018, 58, 60–80. [Google Scholar] [CrossRef]
- Jeevanandam, J.; Aing, Y.S.; Chan, Y.S.; Pan, S.; Danquah, M.K. Nanoformulation and application of phytochemicals as antimicrobial agents. In Antimicrobial Nanoarchitectonics; Elsevier: Amsterdam, The Netherlands, 2017; pp. 61–82. [Google Scholar]
- Cartaxo, A.L.P. Nanoparticles types and properties–understanding these promising devices in the biomedical area. Biol. Mater. Sci. 2015, 1–8. Available online: https://www.semanticscholar.org/paper/Nanoparticles-types-and-properties-%E2%80%93-understanding-Cartaxo/9570b62e051cf8cff1398915cf14eafcff21fbdd?sort=is-influential (accessed on 11 October 2022).
- Wang, L.; Hu, C.; Shao, L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Li, X.; Yu, C.; Wang, Y. Promising Therapeutic Strategies against Microbial Biofilm Challenges. Front. Cell. Infect. Microbiol. 2020, 10, 359. [Google Scholar] [CrossRef]
- Ahmad, N.; Bhatnagar, S.; Ali, S.S.; Dutta, R. Phytofabrication of bioinduced silver nanoparticles for biomedical applications. Int. J. Nanomed. 2015, 10, 7019. [Google Scholar]
- Swidan, N.S.; Hashem, Y.A.; Elkhatib, W.F.; Yassien, M.A. Antibiofilm activity of green synthesized silver nanoparticles against biofilm associated enterococcal urinary pathogens. Sci. Rep. 2022, 12, 3869. [Google Scholar] [CrossRef]
- Muthulakshmi, L.; Suganya, K.; Murugan, M.; Annaraj, J.; Duraipandiyan, V.; Al Farraj, D.A.; Elshikh, M.S.; Juliet, A.; Pasupuleti, M.; Arockiaraj, J. Antibiofilm efficacy of novel biogenic silver nanoparticles from Terminalia catappa against food-borne Listeria monocytogenes ATCC 15313 and mechanisms investigation in-vivo and in-vitro. J. King Saud Univ. Sci. 2022, 34, 102083. [Google Scholar] [CrossRef]
- Salem, S.S.; Badawy, M.S.E.; Al-Askar, A.A.; Arishi, A.A.; Elkady, F.M.; Hashem, A.H. Green biosynthesis of selenium nanoparticles using orange peel waste: Characterization, antibacterial and antibiofilm activities against multidrug-resistant bacteria. Life 2022, 12, 893. [Google Scholar] [CrossRef] [PubMed]
- Rajivgandhi, G.N.; Maruthupandy, M.; Li, J.-L.; Dong, L.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Alanzi, K.F.; Li, W.-J. Photocatalytic reduction and anti-bacterial activity of biosynthesized silver nanoparticles against multi drug resistant Staphylococcus saprophyticus BDUMS 5 (MN310601). Mater. Sci. Eng. C. 2020, 114, 111024. [Google Scholar] [CrossRef] [PubMed]
- Mohanta, Y.K.; Biswas, K.; Jena, S.K.; Hashem, A.; Abd_Allah, E.F.; Mohanta, T.K. Anti-biofilm and antibacterial activities of silver nanoparticles synthesized by the reducing activity of phytoconstituents present in the Indian medicinal plants. Front. Microbiol. 2020, 11, 1143. [Google Scholar] [CrossRef] [PubMed]
- Mariadoss, A.V.A.; Ramachandran, V.; Shalini, V.; Agilan, B.; Franklin, J.H.; Sanjay, K.; Alaa, Y.G.; Tawfiq, M.A.-A.; Ernest, D. Green synthesis, characterization and antibacterial activity of silver nanoparticles by Malus domestica and its cytotoxic effect on (MCF-7) cell line. Microb. Pathog. 2019, 135, 103609. [Google Scholar] [CrossRef]
- Shah, S.; Gaikwad, S.; Nagar, S.; Kulshrestha, S.; Vaidya, V.; Nawani, N.; Pawar, S. Biofilm inhibition and anti-quorum sensing activity of phytosynthesized silver nanoparticles against the nosocomial pathogen Pseudomonas aeruginosa. Biofouling 2019, 35, 34–49. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Wang, P.; Li, T.; Huang, X.; Guo, W.; Yang, Y.; Yan, M.; Zhang, H.; Cai, D.; Jia, X. Self-assembled natural phytochemicals for synergistically antibacterial application from the enlightenment of traditional Chinese medicine combination. Acta Pharm. Sin. B. 2020, 10, 1784–1795. [Google Scholar] [CrossRef]
- Huang, X.; Wang, P.; Li, T.; Tian, X.; Guo, W.; Xu, B.; Huang, G.; Cai, D.; Zhou, F.; Zhang, H. Self-assemblies based on traditional medicine berberine and cinnamic acid for adhesion-induced inhibition multidrug-resistant Staphylococcus aureus. ACS Appl. Mater. Interfaces 2019, 12, 227–237. [Google Scholar] [CrossRef]
- Cherian, T.; Ali, K.; Saquib, Q.; Faisal, M.; Wahab, R.; Musarrat, J. Cymbopogon citratus functionalized green synthesis of CuO-nanoparticles: Novel prospects as antibacterial and antibiofilm agents. Biomolecules 2020, 10, 169. [Google Scholar] [CrossRef] [Green Version]
- Lotha, R.; Shamprasad, B.R.; Sundaramoorthy, N.S.; Nagarajan, S.; Sivasubramanian, A. Biogenic phytochemicals (cassinopin and isoquercetin) capped copper nanoparticles (ISQ/CAS@ CuNPs) inhibits MRSA biofilms. Microb. Pathog. 2019, 132, 178–187. [Google Scholar] [CrossRef]
- Cherian, T.; Ali, K.; Fatima, S.; Saquib, Q.; Ansari, S.M.; Alwathnani, H.A.; Al-Khedhairy, A.A.; Al-Shaeri, M.; Musarrat, J. Myristica fragrans bio-active ester functionalized ZnO nanoparticles exhibit antibacterial and antibiofilm activities in clinical isolates. J. Microbiol. Methods 2019, 166, 105716. [Google Scholar] [CrossRef] [PubMed]
- Scriboni, A.B.; Couto, V.M.; Ribeiro, L.N.D.M.; Freires, I.A.; Groppo, F.C.; De Paula, E.; Franz-Montan, M.; Cogo-Müller, K. Fusogenic liposomes increase the antimicrobial activity of vancomycin against Staphylococcus aureus biofilm. Front. Pharmacol. 2019, 10, 1401. [Google Scholar] [CrossRef] [PubMed]
- Zahra, M.-J.; Hamed, H.; Mohammad, R.-Y.; Nosratollah, Z.; Akbarzadeh, A.; Morteza, M. Evaluation and study of antimicrobial activity of nanoliposomal meropenem against Pseudomonas aeruginosa isolates. Artif. Cells Nanomed. Biotechnol. 2017, 45, 975–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paliwal, R.; Paliwal, S.R.; Kenwat, R.; Kurmi, B.D.; Sahu, M.K. Solid lipid nanoparticles: A review on recent perspectives and patents. Expert Opin. Ther. Pat. 2020, 30, 179–194. [Google Scholar] [CrossRef]
- Singh, B.; Vuddanda, P.R.; Vijayakumar, M.; Kumar, V.; Saxena, P.S.; Singh, S. Cefuroxime axetil loaded solid lipid nanoparticles for enhanced activity against S. aureus biofilm. Colloids Surf. B. 2014, 121, 92–98. [Google Scholar] [CrossRef]
- Nafee, N.; Husari, A.; Maurer, C.K.; Lu, C.; de Rossi, C.; Steinbach, A.; Hartmann, R.W.; Lehr, C.-M.; Schneider, M. Antibiotic-free nanotherapeutics: Ultra-small, mucus-penetrating solid lipid nanoparticles enhance the pulmonary delivery and anti-virulence efficacy of novel quorum sensing inhibitors. J. Control. Release 2014, 192, 131–140. [Google Scholar] [CrossRef]
- Gómez-Gómez, B.; Arregui, L.; Serrano, S.; Santos, A.; Pérez-Corona, T.; Madrid, Y. Selenium and tellurium-based nanoparticles as interfering factors in quorum sensing-regulated processes: Violacein production and bacterial biofilm formation. Metallomics 2019, 11, 1104–1114. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Hensel, A.; Goycoolea, F.M. Chitosan/cyclodextrin surface-adsorbed naringenin-loaded nanocapsules enhance bacterial quorum quenching and anti-biofilm activities. Colloids Surf. B. 2022, 211, 112281. [Google Scholar] [CrossRef]
- Nava-Ortiz, C.A.; Burillo, G.; Concheiro, A.; Bucio, E.; Matthijs, N.; Nelis, H.; Coenye, T.; Alvarez-Lorenzo, C. Cyclodextrin-functionalized biomaterials loaded with miconazole prevent Candida albicans biofilm formation in vitro. Acta Biomater. 2010, 6, 1398–1404. [Google Scholar] [CrossRef]
- Gharbi, A.; Humblot, V.; Turpin, F.; Pradier, C.-M.; Imbert, C.; Berjeaud, J.-M. Elaboration of antibiofilm surfaces functionalized with antifungal-cyclodextrin inclusion complexes. FEMS Microbiol. Immunol. 2012, 65, 257–269. [Google Scholar] [CrossRef]
- Huang, Y.; Bai, L.; Yang, Y.; Yin, Z.; Guo, B. Biodegradable gelatin/silver nanoparticle composite cryogel with excellent antibacterial and antibiofilm activity and hemostasis for Pseudomonas aeruginosa-infected burn wound healing. J. Colloid Interface Sci. 2022, 608, 2278–2289. [Google Scholar] [CrossRef] [PubMed]
- Haidari, H.; Bright, R.; Garg, S.; Vasilev, K.; Cowin, A.J.; Kopecki, Z. Eradication of mature bacterial biofilms with concurrent improvement in chronic wound healing using silver nanoparticle hydrogel treatment. Biomedicines 2021, 9, 1182. [Google Scholar] [CrossRef] [PubMed]
- Hemmingsen, L.M.; Giordani, B.; Pettersen, A.K.; Vitali, B.; Basnet, P.; Škalko-Basnet, N. Liposomes-in-chitosan hydrogel boosts potential of chlorhexidine in biofilm eradication in vitro. Carbohydr. Polym. 2021, 262, 117939. [Google Scholar] [CrossRef]
- Li, X.; Fu, Y.-n.; Huang, L.; Liu, F.; Moriarty, T.F.; Tao, L.; Wei, Y.; Wang, X. Combating biofilms by a self-adapting drug loading hydrogel. ACS Appl. Bio Mater. 2021, 4, 6219–6226. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.; Bu, Q.; Tao, N.; Chen, M.; Liu, H.; Zhou, J.; Liu, J.; Deng, B.; Kong, N.; Zhang, X. A facile and general method for synthesis of antibiotic-free protein-based hydrogel: Wound dressing for the eradication of drug-resistant bacteria and biofilms. Bioact. Mater. 2022, 18, 446–458. [Google Scholar] [CrossRef]
- Chen, H.; Cheng, J.; Cai, X.; Han, J.; Chen, X.; You, L.; Xiong, C.; Wang, S. pH-Switchable antimicrobial supramolecular hydrogels for synergistically eliminating biofilm and promoting wound healing. ACS Appl. Mater. Interfaces 2022, 14, 18120–18132. [Google Scholar] [CrossRef]
- Alfuraydi, R.T.; Alminderej, F.M.; Mohamed, N.A. Evaluation of antimicrobial and anti-biofilm formation activities of novel poly (vinyl alcohol) hydrogels reinforced with crosslinked chitosan and silver nano-particles. Polymers 2022, 14, 1619. [Google Scholar] [CrossRef]
- Wroe, J.A.; Johnson, C.T.; García, A.J. Bacteriophage delivering hydrogels reduce biofilm formation in vitro and infection in vivo. J. Biomed. Mater. Res. A 2020, 108, 39–49. [Google Scholar] [CrossRef]
- Barros, J.A.R.; de Melo, L.D.R.; da Silva, R.A.R.; Ferraz, M.P.; de Rodrigues Azeredo, J.C.V.; de Carvalho Pinheiro, V.M.; Colaço, B.J.A.; Fernandes, M.H.R.; de Sousa Gomes, P.; Monteiro, F.J. Encapsulated bacteriophages in alginate-nanohydroxyapatite hydrogel as a novel delivery system to prevent orthopedic implant-associated infections. Nanomed. Nanotechnol. Biol. Med. 2020, 24, 102145. [Google Scholar] [CrossRef] [Green Version]
- Tarawneh, O.; Abu Mahfouz, H.; Hamadneh, L.; Deeb, A.A.; Al-Sheikh, I.; Alwahsh, W.; Fadhil Abed, A. Assessment of persistent antimicrobial and anti-biofilm activity of p-HEMA hydrogel loaded with rifampicin and cefixime. Sci. Rep. 2022, 12, 3900. [Google Scholar] [CrossRef]
- Anjum, A.; Sim, C.-H.; Ng, S.-F. Hydrogels containing antibiofilm and antimicrobial agents beneficial for biofilm-associated wound infection: Formulation characterizations and In vitro study. AAPS PharmSciTech 2018, 19, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, L.; Tessier, J.; Douyere, G.; Nardello-Rataj, V.; Schmitzer, A.R. Phytochemical-and cyclodextrin-based pickering emulsions: Natural potentiators of antibacterial, antifungal, and antibiofilm activity. Langmuir 2020, 36, 4317–4323. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, G.; Tiwari, R.; Rai, A.K. Cyclodextrins in delivery systems: Applications. J. Pharm. Bioallied Sci. 2010, 2, 72. [Google Scholar] [CrossRef]
- Agrahari, A.K.; Singh, A.K.; Singh, A.S.; Singh, M.; Maji, P.; Yadav, S.; Rajkhowa, S.; Prakash, P.; Tiwari, V.K. Click inspired synthesis of p-tert-butyl calix [4] arene tethered benzotriazolyl dendrimers and their evaluation as anti-bacterial and anti-biofilm agents. New J. Chem. 2020, 44, 19300–19313. [Google Scholar] [CrossRef]
- Barik, S.K.; Singh, B.N. Nanoemulsion-loaded hydrogel coatings for inhibition of bacterial virulence and biofilm formation on solid surfaces. Sci. Rep. 2019, 9, 6520. [Google Scholar]
- Liang, Y.; He, J.; Guo, B. Functional hydrogels as wound dressing to enhance wound healing. ACS Nano 2021, 15, 12687–12722. [Google Scholar] [CrossRef]
- Dong, D.; Thomas, N.; Thierry, B.; Vreugde, S.; Clive, A.; Prestidge, C.A.; Wormald, P.-J. Distribution and inhibition of liposomes on Staphylococcus aureus and Pseudomonas aeruginosa biofilm. PLoS ONE 2015, 10, e0131806. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y. Liposome as a delivery system for the treatment of biofilm-mediated infections. J. Appl. Microbiol. 2021, 131, 2626–2639. [Google Scholar] [CrossRef]
- Wang, C.; Chen, P.; Qiao, Y.; Kang, Y.; Yan, C.; Yu, Z.; Wang, J.; He, X.; Wu, H. pH responsive superporogen combined with PDT based on poly Ce6 ionic liquid grafted on SiO2 for combating MRSA biofilm infection. Theranostics 2020, 10, 4795. [Google Scholar] [CrossRef]
- Zhao, Z.; Ding, C.; Wang, Y.; Tan, H.; Li, J. pH-Responsive polymeric nanocarriers for efficient killing of cariogenic bacteria in biofilms. Biomater. Sci. 2019, 7, 1643–1651. [Google Scholar] [CrossRef]
- Singh, P.; Pandit, S.; Beshay, M.; Mokkapati, V.; Garnaes, J.; Olsson, M.E.; Sultan, A.; Mackevica, A.; Mateiu, R.V.; Lütken, H. Anti-biofilm effects of gold and silver nanoparticles synthesized by the Rhodiola rosea rhizome extracts. Artif. Cells Nanomed. Biotechnol. 2018, 46, S886–S899. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, L.; Su, L.; van der Mei, H.C.; Jutte, P.C.; Ren, Y.; Busscher, H.J. Nanotechnology-based antimicrobials and delivery systems for biofilm-infection control. Chem. Soc. Rev. 2019, 48, 428–446. [Google Scholar] [CrossRef] [PubMed]
- Szerencsés, B.; Igaz, N.; Tóbiás, Á.; Prucsi, Z.; Rónavári, A.; Bélteky, P.; Madarász, D.; Papp, C.; Makra, I.; Vágvölgyi, C. Size-dependent activity of silver nanoparticles on the morphological switch and biofilm formation of opportunistic pathogenic yeasts. BMC Microbiol. 2020, 20, 176. [Google Scholar] [CrossRef] [PubMed]
- Pham, P.; Oliver, S.; Wong, E.H.; Boyer, C. Effect of hydrophilic groups on the bioactivity of antimicrobial polymers. Polym. Chem. 2021, 12, 5689–5703. [Google Scholar] [CrossRef]
- Olmo, J.A.-D.; Ruiz-Rubio, L.; Pérez-Alvarez, L.; Sáez-Martínez, V.; Vilas-Vilela, J.L. Antibacterial coatings for improving the performance of biomaterials. Coatings 2020, 10, 139. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Li, J.; Zhang, Y.; Wang, Y.; Liu, L.; Li, M. Inhibition of gold nanoparticles (AuNPs) on pathogenic biofilm formation and invasion to host cells. Sci. Rep. 2016, 6, 26667. [Google Scholar] [CrossRef]
- Giri, K.; Yepes, L.R.; Duncan, B.; Parameswaran, P.K.; Yan, B.; Jiang, Y.; Bilska, M.; Moyano, D.F.; Thompson, M.A.; Rotello, V.M. Targeting bacterial biofilms via surface engineering of gold nanoparticles. RSC Adv. 2015, 5, 105551–105559. [Google Scholar] [CrossRef] [Green Version]
- Sathyanarayanan, M.B.; Balachandranath, R.; Genji Srinivasulu, Y.; Kannaiyan, S.K.; Subbiahdoss, G. The effect of gold and iron-oxide nanoparticles on biofilm-forming pathogens. Int. Sch. Res. Not. 2013, 2013, 272086. [Google Scholar] [CrossRef] [Green Version]
- Van de Walle, A.; Perez, J.E.; Abou-Hassan, A.; Hémadi, M.; Luciani, N.; Wilhelm, C. Magnetic nanoparticles in regenerative medicine: What of their fate and impact in stem cells? Mater. Today Nano 2020, 11, 100084. [Google Scholar] [CrossRef]
- Belkahla, H.; Mazarío, E.; Sangnier, A.P.; Lomas, J.S.; Gharbi, T.; Ammar, S.; Micheau, O.; Wilhelm, C.; Hémadi, M. TRAIL acts synergistically with iron oxide nanocluster-mediated magneto-and photothermia. Theranostics 2019, 9, 5924. [Google Scholar] [CrossRef]
- Di Corato, R.; Béalle, G.; Kolosnjaj-Tabi, J.; Espinosa, A.; Clement, O.; Silva, A.K.; Menager, C.; Wilhelm, C. Combining magnetic hyperthermia and photodynamic therapy for tumor ablation with photoresponsive magnetic liposomes. ACS Nano 2015, 9, 2904–2916. [Google Scholar] [CrossRef] [PubMed]
- Hasanzadeh, M.; Shadjou, N.; de la Guardia, M. Iron and iron-oxide magnetic nanoparticles as signal-amplification elements in electrochemical biosensing. TrAC Trends Anal. Chem. 2015, 72, 1–9. [Google Scholar] [CrossRef]
- Kim, S.H.; Kwak, S.-Y.; Sohn, B.-H.; Park, T.H. Design of TiO2 nanoparticle self-assembled aromatic polyamide thin-film-composite (TFC) membrane as an approach to solve biofouling problem. J. Membr. Sci. 2003, 211, 157–165. [Google Scholar] [CrossRef]
- Scioli Montoto, S.; Muraca, G.; Ruiz, M.E. Solid lipid nanoparticles for drug delivery: Pharmacological and biopharmaceutical aspects. Front. Mol. Biosci. 2020, 7, 587997. [Google Scholar] [CrossRef]
- Liu, Y.; Naha, P.C.; Hwang, G.; Kim, D.; Huang, Y.; Simon-Soro, A.; Jung, H.-I.; Ren, Z.; Li, Y.; Gubara, S. Topical ferumoxytol nanoparticles disrupt biofilms and prevent tooth decay in vivo via intrinsic catalytic activity. Nat. Commun. 2018, 9, 2920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qayyum, S.; Oves, M.; Khan, A.U. Obliteration of bacterial growth and biofilm through ROS generation by facilely synthesized green silver nanoparticles. PLoS ONE 2017, 12, e0181363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddique, M.H.; Aslam, B.; Imran, M.; Ashraf, A.; Nadeem, H.; Hayat, S.; Khurshid, M.; Afzal, M.; Malik, I.R.; Shahzad, M. Effect of silver nanoparticles on biofilm formation and EPS production of multidrug-resistant Klebsiella pneumoniae. Biomed Res. Int. 2020, 2020, 6398165. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.P.; Wang, L.; Chen, Y.-P.; Pellechia, P.J.; Benicewicz, B.C.; Decho, A.W. Engineering nanoparticles to silence bacterial communication. Front. Microbiol. 2015, 6, 189. [Google Scholar] [CrossRef] [Green Version]
- Mangzira Kemung, H.; Tan, L.T.-H.; Chan, K.-G.; Ser, H.-L.; Law, J.W.-F.; Lee, L.-H.; Goh, B.-H. Streptomyces sp. strain MUSC 125 from mangrove soil in Malaysia with anti-MRSA, anti-biofilm and antioxidant activities. Molecules 2020, 25, 3545. [Google Scholar] [CrossRef] [PubMed]
- Younis, K.M.; Usup, G.; Ahmad, A. Secondary metabolites produced by marine streptomyces as antibiofilm and quorum-sensing inhibitor of uropathogen Proteus mirabilis. Environ. Sci. Pollut. Res. 2016, 23, 4756–4767. [Google Scholar] [CrossRef]
- Peach, K.C.; Cheng, A.T.; Oliver, A.G.; Yildiz, F.H.; Linington, R.G. Discovery and biological characterization of the auromomycin chromophore as an inhibitor of biofilm formation in Vibrio cholerae. Chembiochem 2013, 14, 2209–2215. [Google Scholar] [CrossRef] [PubMed]
- Manefield, M.; de Nys, R.; Naresh, K.; Roger, R.; Givskov, M.; Peter, S.; Kjelleberg, S. Evidence that halogenated furanones from Delisea pulchra inhibit acylated homoserine lactone (AHL)-mediated gene expression by displacing the AHL signal from its receptor protein. Microbiology 1999, 145, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.; Bedzyk, L.A.; Setlow, P.; England, D.F.; Kjelleberg, S.; Thomas, S.M.; Ye, R.W.; Wood, T.K. Differential gene expression to investigate the effect of (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2 (5H)-furanone on Bacillus subtilis. Appl. Environ. Microbiol. 2004, 70, 4941–4949. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.; Sims, J.; Wood, T. Inhibition of biofilm formation and swarming of Bacillus subtilis by (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2 (5H)-furanone. Lett. Appl. Microbiol. 2002, 34, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Manefield, M.; Rasmussen, T.B.; Henzter, M.; Andersen, J.B.; Steinberg, P.; Kjelleberg, S.; Givskov, M. Halogenated furanones inhibit quorum sensing through accelerated LuxR turnover. Microbiology 2002, 148, 1119–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahreen, S.; Mukhtar, H.; Imre, K.; Morar, A.; Herman, V.; Sharif, S. Exploring the function of quorum sensing regulated biofilms in biological wastewater treatment: A review. Int. J. Mol. Sci. 2022, 23, 9751. [Google Scholar] [CrossRef] [PubMed]
- Pereira, U.A.; Barbosa, L.C.; Maltha, C.R.; Demuner, A.J.; Masood, M.A.; Pimenta, A.L. γ-Alkylidene-γ-lactones and isobutylpyrrol-2 (5H)-ones analogues to rubrolides as inhibitors of biofilm formation by gram-positive and gram-negative bacteria. Bioorg. Med. Chem. Lett. 2014, 24, 1052–1056. [Google Scholar] [CrossRef] [PubMed]
- Tello, E.; Castellanos, L.; Arevalo-Ferro, C.; Rodríguez, J.; Jiménez, C.; Duque, C. Absolute stereochemistry of antifouling cembranoid epimers at C-8 from the Caribbean octocoral Pseudoplexaura flagellosa. Revised structures of plexaurolones. Tetrahedron 2011, 67, 9112–9121. [Google Scholar] [CrossRef]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, physiology, and natural products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Cai, Z.H.; Lao, Y.M.; Jin, H.; Ying, K.Z.; Lin, G.H.; Zhou, J. Antibiofilm activity substances derived from coral symbiotic bacterial extract inhibit biofouling by the model strain Pseudomonas aeruginosa PAO 1. Microb. Biotechnol. 2018, 11, 1090–1105. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Chen, J.; Yan, Y.; Chen, S.; Xu, X.; Zhang, H.; Wang, H. Quorum sensing inhibitors from marine bacteria Oceanobacillus sp. XC22919. Nat. Prod. Res. 2019, 33, 1819–1823. [Google Scholar] [CrossRef] [PubMed]
- Goel, N.; Fatima, S.W.; Kumar, S.; Sinha, R.; Khare, S.K. Antimicrobial resistance in biofilms: Exploring marine actinobacteria as a potential source of antibiotics and biofilm inhibitors. Biotechnol. Rep. 2021, 30, e00613. [Google Scholar] [CrossRef] [PubMed]
- Marappa, N.; Dharumadurai, D.; Nooruddin, T.; Abdulkader, A.M. Morphological, molecular characterization and biofilm inhibition effect of endophytic Frankia sp. from root nodules of Actinorhizal plant Casuarina sp. S. Afr. J. Bot. 2020, 134, 72–83. [Google Scholar] [CrossRef]
- Singh, R.; Dubey, A.K. Isolation and characterization of a new endophytic actinobacterium Streptomyces californicus strain ADR1 as a promising source of anti-bacterial, anti-biofilm and antioxidant metabolites. Microorganisms 2020, 8, 929. [Google Scholar] [CrossRef] [PubMed]
- Kamarudheen, N.; Naushad, T.; Rao, K.V.B. Biosynthesis, characterization and antagonistic applications of extracellular melanin pigment from marine Nocardiopsis Sps. Ind. J. Pharm. Educ. Res 2019, 53, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Ramalingam, V.; Mahamuni, D.; Rajaram, R. In vitro and in silico approaches of antibiofilm activity of 1-hydroxy-1-norresistomycin against human clinical pathogens. Microb. Pathog. 2019, 132, 343–354. [Google Scholar] [CrossRef]
- Rajivgandhi, G.; Vijayan, R.; Maruthupandy, M.; Vaseeharan, B.; Manoharan, N. Antibiofilm effect of Nocardiopsis sp. GRG 1 (KT235640) compound against biofilm forming Gram negative bacteria on UTIs. Microb. Pathog. 2018, 118, 190–198. [Google Scholar] [CrossRef]
- Miao, L.; Xu, J.; Yao, Z.; Jiang, Y.; Zhou, H.; Jiang, W.; Dong, K. The anti-quorum sensing activity and bioactive substance of a marine derived Streptomyces. Biotechnol. Biotechnol. Equip. 2017, 31, 1007–1015. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.; Zhou, J.; Zhu, X.; Yu, S.; Chen, L.; Jin, H.; Cai, Z. Strain identification and quorum sensing inhibition characterization of marine-derived Rhizobium sp. NAO1. R. Soc. Open Sci. 2017, 4, 170025. [Google Scholar] [CrossRef] [Green Version]
- Stumpp, N.; Premnath, P.; Schmidt, T.; Ammermann, J.; Draeger, G.; Reck, M.; Jansen, R.; Stiesch, M.; Wagner-Döbler, I.; Kirschning, A. Synthesis of new carolacton derivatives and their activity against biofilms of oral bacteria. Org. Biomol. Chem. 2015, 13, 5765–5774. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.-S.; Eom, Y.-B. Antimicrobial and antibiofilm activities of Clostridium butyricum supernatant against Acinetobacter baumannii. Arch. Microbiol. 2020, 202, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Shokri, D.; Khorasgani, M.R.; Mohkam, M.; Fatemi, S.M.; Ghasemi, Y.; Taheri-Kafrani, A. The inhibition effect of lactobacilli against growth and biofilm formation of Pseudomonas aeruginosa. Probiotics Antimicrob. 2018, 10, 34–42. [Google Scholar] [CrossRef]
- Song, H.; Zhang, J.; Qu, J.; Liu, J.; Yin, P.; Zhang, G.; Shang, D. Lactobacillus rhamnosus GG microcapsules inhibit Escherichia coli biofilm formation in coculture. Biotechnol. Lett. 2019, 41, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Roscetto, E.; Masi, M.; Esposito, M.; Di Lecce, R.; Delicato, A.; Maddau, L.; Calabrò, V.; Evidente, A.; Catania, M.R. Anti-biofilm activity of the fungal phytotoxin sphaeropsidin A against clinical isolates of antibiotic-resistant bacteria. Toxins 2020, 12, 444. [Google Scholar] [CrossRef]
- Petersen, L.-E.; Marner, M.; Labes, A.; Tasdemir, D. Rapid metabolome and bioactivity profiling of fungi associated with the leaf and rhizosphere of the Baltic seagrass Zostera marina. Mar. Drugs 2019, 17, 419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narmani, A.; Teponno, R.B.; Helaly, S.E.; Arzanlou, M.; Stadler, M. Cytotoxic, anti-biofilm and antimicrobial polyketides from the plant associated fungus Chaetosphaeronema achilleae. Fitoterapia 2019, 139, 104390. [Google Scholar] [CrossRef]
- Arora, P.; Wani, Z.A.; Nalli, Y.; Ali, A.; Riyaz-Ul-Hassan, S. Antimicrobial potential of thiodiketopiperazine derivatives produced by Phoma sp., an endophyte of Glycyrrhiza glabra Linn. Microb. Ecol. 2016, 72, 802–812. [Google Scholar] [CrossRef]
- Rashmi, M.; Meena, H.; Meena, C.; Kushveer, J.; Busi, S.; Murali, A.; Sarma, V. Anti-quorum sensing and antibiofilm potential of Alternaria alternata, a foliar endophyte of Carica papaya, evidenced by QS assays and in-silico analysis. Fungal Biol. 2018, 122, 998–1012. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, M.; Zhu, X.; Yu, W.; Gong, Q. Equisetin as potential quorum sensing inhibitor of Pseudomonas aeruginosa. Biotechnol. Lett. 2018, 40, 865–870. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polysaccharides | Microbes | Role | Ref. |
|---|---|---|---|
| PGA (Poly-β-1,6-N-acetyl-D-glucosamine) | Actinobacillus actinomycetemcomitans | Intercellular adhesion Cellular detachment Dispersion | [28] |
| Colanic acid | E. coli | Antidesiccative | [29] |
| Galactopyranosyl-glycerol-phosphate | Bacillus licheniformis | Antibiofilm | [30] |
| Alginate, Pel, Psl | P. aeruginosa | Biofilm structural stability maintenance Cell communication and differentiation | [31,32] |
| Capsular polysaccharide, cellulose | Salmonella Typhimurium | Adhesion Environmental survival | [33] |
| Targets | sRNAs | Bacteria | Regulatory Effect | Refs. |
|---|---|---|---|---|
| AphA, HapR | Qrr1-4, Qrr1-5 | V. cholera, V. harveyi | Activation | [72] |
| Crc | CrcZ | Pseudomonas spp. | Repression | [73] |
| CsgD | GcvB | E. coli | Repression | [74] |
| McaS | E. coli | Repression | [75] | |
| OmrA, OmrB | E. coli | Repression | [76] | |
| SdsR | S. Typhimurium | Activation | [77] | |
| CsgD, YdaM | RprA | E. coli | Repression | [78] |
| CsgD, RpoS | ArcZ | S. Typhimurium, E. coli | Activation | [77,79,80] |
| CsrA | CsrB, CsrC | Yersinia pseudotuberculosis, E. coli | Repression | [81,82] |
| PgaA | McaS | E. coli | Activation | [74] |
| PqsR | PhrS | P. aeruginosa | Activation | [83] |
| RpoS | DsrA | E. coli | Activation | [80] |
| OxyS | E. coli | Repression | [80] | |
| RprA | E. coli | Activation | [84] |
| Pathogenic Microbes | Targeted Area | Consequences | Refs. |
|---|---|---|---|
| Group A streptococci | Skin | Necrotizing fasciitis, tissue necrosis | [108] |
| Actinobacillus, Actinomycetemcomitans, Eikenella corrodens, Streptococcus mutans, Prevotella intermedia, Porphyromonas gingivalis, Oral spirochetes. | Oral cavity | Periodontal infections, acute inflammation, teeth loosening due to periodontal tissue breakdown, halitosis | [109,110] |
| Streptococcus spp. Staphylococci (coagulase-negative), S. aureus, Enterococcus spp. | Musculoskeletal system | Bacterial accumulation on implants and dead bones cause biofilm infections. | [111] |
| Haemophilus influenza, Streptococcus pneumoniae, Moraxella catarrhalis. | Middle ear | Otitis media | [112] |
| S. aureus, P. aeruginosa. | Lungs (in patients with cystic fibrosis) | Mucoviscidosis, lung infections | [113] |
| Organism | Contaminated Food Items | Consequences | Refs. |
|---|---|---|---|
| Anoxybacillus flavithermus | Milk powder | Reduced acceptability of powdered milk | [126] |
| B. cereus | Meat, dairy products, vegetables, and rice | Vomiting, diarrhea | [127,128] |
| E. coli | Meat, vegetables, fruits, and milk | Hemolytic uremic syndrome, diarrhea | [114] |
| Campylobacter jejuni | Unpasteurized milk, animals, poultry | Vomiting, bloody diarrhea, nausea, fever, and stomach cramps | [129] |
| S. enterica | Porcine, bovine, fish, ovine, and poultry meat | Septicemia, gastroenteritis | [130] |
| Geobacillus stearothermophilus | Dairy dried products | Enzymes or acids production resulting in off-flavors | [130] |
| Pseudomonas spp. | Meat, vegetables, fruits, and dairy products | Blue discoloration occurrence on fresh cheese | [131] |
| S. aureus | Dairy products, poultry, eggs, meat, salads, cakes, and pastries | Diarrhea, vomiting | [132,133] |
| L. monocytogenes | Ready-to-eat products, raw milk, dairy products, | Listeriosis in immune-compromised, elderly, and pregnant patients | [134] |
| Pectinatus spp. | Brewery environment and beer | Produces beer turbid due to the production of sulfur compounds | [135] |
| Region and Year | Reported Cases | Responsible Organisms | Food Type | Ref. |
|---|---|---|---|---|
| South Africa (2017–2018) | 1060 | L. monocytogenes | Ready-to-eat meat products | [136] |
| England (2018) | 34 | Clostridium perfringens | Cheese sauce | [137] |
| England (2015) | NA | E. coli O157:H7 | Prepacked salad leaves | [138] |
| Massachusetts (2014–2018) | 1200 per year | Salmonella | NA | [139] |
| England (2016) | 69 | Campylobacter | Raw milk | [140] |
| Belgium (2013) | 52 | S. aureus | Several foods | [141] |
| China (2010–2014) | 1040 | Vibrio parahaemolyticus | NA | [142] |
| Europe (2007–2014) | 6657 | B. cereus | NA | [143] |
| China (2003–2008) | 9041 | V. parahaemolyticus | Meat and aquatic products | [144] |
| Australia (2001–2010) | 667 | L. monocytogenes | NA | [145] |
| Plant Extracts, Compounds or Essential Oils (EOs) | Plant Source | Bacterial Species | Inhibition Concentration | Biofilm Inhibition (%) | Ref. |
|---|---|---|---|---|---|
| Leaf extract | Cochlospermum regium | MRSA | 2000 μg/mL | 100 | [169] |
| 5-Hydroxymethylfurfural | Musa acuminata | P. aeruginosa | 10 μg/mL | 83 | [170] |
| Syringopicroside | Syringa oblata | Streptococcus suis | 1.28 μg/mL | 92 | [171] |
| Xanthohumol | Humulus lupulus | S. aureus | 9.8 μg/mL | 100 | [172] |
| Sotetsuflavone | Cycas media R. Br | E. faecalis | - | 60.87–21.74 | [173] |
| Lemon grass EO | Cymbopogon citratus | S. aureus | 250 μg/mL | 50 | [174] |
| P. aeruginosa | - | ||||
| Citral | P. aeruginosa | 0.40 μg/mL | |||
| S. aureus | >107 μg/mL | ||||
| EO | Cinnamomum verum | Acinetobacter baumanii, Citrobacter freundii Corynebacterium striatum, E. coli, Klebsiella spp., S. aureus, Salmonella spp., P. aeruginosa | 10 μg/mL | 97 | [175] |
| Thymus vulgaris | 88 | ||||
| Eugenia caryophyllata | 91 | ||||
| Ethanol extract | Azadirachta indica | MRSA | 2000 μg/mL | 43.0 | [176] |
| Moringa oleifera | MRSA | 2000 μg/mL | 51.4 | ||
| Murraya koenigii | MRSA | 2000 μg/mL | 44.9 | ||
| Psidium guajava | MRSA | 2000 μg/mL | 80 | ||
| Petroleum ether extract | Azadirachta indica | MRSA | 2000 μg/mL | 83.8 | |
| Moringa oleifera | MRSA | 2000 μg/mL | 59.9 | ||
| Murraya koenigii | MRSA | 2000 μg/mL | 63.7 | ||
| Psidium guajava | MRSA | 2000 μg/mL | 62.9 | ||
| Aqueous extract | Acacia nilotica | K. pneumoniae | 13,300 μg/mL | 59 | [177] |
| E.coli | 13,300 μg/mL | 63 | |||
| P. aeruginosa | 15,000 μg/mL | 39 | |||
| Proteus mirabilis | 16,700 μg/mL | 49 | |||
| EO | Cinnamomum zeylanicum | E. coli | 2000 μg/mL | 82.76 | [178] |
| S. epidermidis | 83.33 | ||||
| Citrus grandis | E. coli | 2000 μg/mL | 58.62 | ||
| S. epidermidis | 46.67 | ||||
| Citrus hystrix | E. coli | 2000 μg/mL | 75.86 | ||
| S. epidermidis | 83.33 | ||||
| Citrus reticulata | E. coli | 2000 μg/mL | 82.76 | ||
| S. epidermidis | 83.33 | ||||
| Psiadia argute | E. coli | 2000 μg/mL | 90 | ||
| S. epidermidis | |||||
| Psiadia terebinthina | E. coli | 2000 μg/mL | 93.67 | ||
| S. epidermidis | 90 | ||||
| Vanilic acid | Vaccinium macrocarpon Aiton (Cranberry) | E. coli | 23.78 mM | 100 | [179] |
| Protocaterchuic | 25.95 mM | ||||
| Catechin | 55.12, 68.9 mM | ||||
| Pulp extract | Euterpe oleracea | S. aureus | 250 μg/mL | 100 | [180] |
| Leaf extract | Juglans regia | P. aeruginosa | 16,000 μg/mL | 60 | [181] |
| Leaf extract | Tetradenia riparia | MRSA | - | 50 | [182] |
| Rosmarinus officinalis | MRSA | 30 μg/mL | 50 | ||
| Extract | Tagetes minuta | Bacillus sp. Mcn4 | 100 μg/mL | 50 | [183] |
| Extract | Tessaria absinthioides | Bacillus spp. | 100 μg/mL | 66 | |
| Staphylococcus sp. Mcr1 | 10–50 μg/mL | 55–62 | |||
| Sesquiterpene lactones | Acanthospermum hispidum | P. aeruginosa | 0.25–2.5 μg/mL | 69–77 | [184] |
| Nanoparticles | Plant Species | Bacteria | Inhibition Concentration | Biofilm Inhibition (%) | Ref. |
|---|---|---|---|---|---|
| Ag NPs | Morinda citrifolia | S. aureus | 60 μg/mL | 96 | [204] |
| Glochidion lanceolarium | P. aeruginosa, E. coli, S. aureus | 68.9, 12.9, 23.4 μg/mL | 99 | [205] | |
| Semecarpus anacardium | P. aeruginosa, E. coli, S. aureus | 45.5, 23.4, 64.1 μg/mL | >99 | ||
| Bridelia retusa | P. aeruginosa, E. coli, S. aureus | 52.5, 33.8, 32.7 μg/mL | |||
| Malus domestica | K. pneumoniae | 24.6 μg/mL | 34 | [206] | |
| Enterobacter aerogenes | 35.6 μg/mL | 72 | |||
| Piper betle | P. aeruginosa | 8 μg/mL | 78 | [207] | |
| BER-RHE NPs | Coptis chinensis (Berberine), Rheum palmatum L. (Rhein) | S. aureus | 0.1 mmol/mL | 96 | [208] |
| CA-BBR NPs | Coptidis rhizome (Berberine) Cinnamomi cortex (Cinnamic acid) | MRSA | 0.1 μmol/mL | 64 | [209] |
| Cu NPs | Cymbopogon citratus | E. coli | 2000 μg/mL | 49 | [210] |
| MRSA | 33 | ||||
| Crotalaria candicans | MRSA | 1 μg/mL | >75 | [211] | |
| ZnO NPs | Myristica fragrans | E. coli | 1000 μg/mL | 24 | [212] |
| MRSA | 1500 μg/mL | 51 |
| Nanomaterials | Target Organism | Impact on Biofilm | Refs. |
|---|---|---|---|
| Cyclodextrins | S. aureus, MRSA, C. albicans, P. aeruginosa, P. vulgaris, E. faecalis, E. coli | Adhesion inhibition, biofilm eradication | [221,233,234] |
| Dendrimers | MRSA, MSSA, E. coli, K. pneumoniae, P. aeruginosa | Biofilm inhibition | [235] |
| Hydrogels | P. aeruginosa, MRSA, S. aureus, A. baumanii | Biofilm eradication, wound healing | [225,236,237] |
| Liposomes | S. aureus, P. gingivalis | Growth inhibition, biofilm formation reduction | [238,239] |
| Polymeric NPs | E. coli, S. aureus, S. mutans, En. Cloacae, P. aeruginosa | Growth inhibition, matrix disruption, and eradication. | [240,241] |
| Stimuli-responsive NPs: Ag NPs | S. aureus, E. coli, P. aeruginosa, S. flexneri, K. pneumoniae, S. mutans C. albicans, S. aureus, E. coli, P. aeruginosa, P. aeruginosa, H. pylori, S. aureus, S. mutans, M. tuberculosis | Structural alternation, inhibition, oxidative stress. | [236,242,243,244] |
| Au NPs | Growth inhibition, matrix disruption. | [245,246,247,248,249] | |
| SPIONs | Colonization prevention, cell lysis, oxidative stress | [250,251,252,253,254] | |
| Solid Lipid NPs | S. aureus | Growth inhibition | [215,255] |
| Other inorganic NPs | S. aureus, P. aeruginosa, E. coli, S. epidermidis | Growth inhibition, matrix disruption | [205,256,257,258,259] |
| Anti-Biofilm Compounds | Source | Target Organism | Biofilm Inhibition | Ref. |
|---|---|---|---|---|
| Actinobacteria | ||||
| Carotenoid pigment | Streptomyces parvulus | C. albicans | >50% | [273] |
| Bioactive metabolites | Frankia casuarinae DDNSF-02 | Candida sp. Pseudomonas sp. | 59–81% 65–80% | [274] |
| Secondary metabolites | Streptomyces californicus Strain ADR1 | S. aureus and MRSA | 90% | [275] |
| Melanin pigments | Nocardiopsis dassonvillei strain JN1, Nocardiopsis sp. JN2 | Staphylococcus sp. | 64.20% (JN1) 65.99% (JN2) | [276] |
| 1-hydroxy-1norresistomycin (HNM) | Streptomyces variabilis | V. cholera E. coli S. aureus | 92% 96% 93% | [277] |
| Pyrrolo (1,2-a) pyrazine-1,4-dione hexahydro-3-(2-methylpropyl) | Actinomycetes Nocardiopsis sp. GRG 1 (KT235640) | E. coli P. mirabilis | 77% 82% | [278] |
| Actinomycin-D | S.parvulus | S. aureus Ruegeria sp. P. aeruginosa Micrococcus luteus | 53.72% 45.98% 37.12% 22.20% | [279] |
| Bacterial compounds | ||||
| Secondary metabolites | Streptomyces (marine sediment) | P. mirabilis | 63–26% | [261] |
| N-acyl homoserine lactone-based QS analogs | Aqueous extract of Rhizobium sp. NAO1 | P. aeruginosa | 77.9% | [280] |
| Carolacton | Extract of Sorangium cellulosum | Streptococcus oralis, Streptococcus gordonii, S. mutans, A. actinomycetemocomitans | Reduced biofilm formation | [281] |
| CFS | Clostridium butyricum | A. baumannii (MDR strain) | Dose-dependent biofilm inhibition | [282] |
| CFS | Lactobacillus strains | P. aeruginosa | 0–64% 100% (with L. fermentum L1 and L2) | [283] |
| CFS | Lacticasebacillus rhamnosus GG | E. coli | Dose and time-dependent biofilm disruption | [284] |
| Fungal compounds | ||||
| Diterpenoid sphaeropsidin A | Diploidia corticola | P. aeruginosa (MDR strain) MRSA | 62% 53% | [285] |
| Organic extracts | Penicillium sp. | P. aeruginosa | Biofilm formation reduction by QS inhibition | [286] |
| Vulculic acid, curvulol | Chaetosphaeronema achilleae | S. aureus DSM 1104 | 91.9–96.8% | [287] |
| Thiodiketopiperazine derivatives | Phoma sp. GG1F1 | S. pyogenes S. aureus | 60.7–86% 28–57% | [288] |
| Crude extract | Alternaria alternate | P. aeruginosa PAO1 | 65.2% | [289] |
| Equisetin | Fusarium sp. Z10 | P. aeruginosa PAO1 | 58.3% | [290] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asma, S.T.; Imre, K.; Morar, A.; Imre, M.; Acaroz, U.; Shah, S.R.A.; Hussain, S.Z.; Arslan-Acaroz, D.; Istanbullugil, F.R.; Madani, K.; et al. Natural Strategies as Potential Weapons against Bacterial Biofilms. Life 2022, 12, 1618. https://doi.org/10.3390/life12101618
Asma ST, Imre K, Morar A, Imre M, Acaroz U, Shah SRA, Hussain SZ, Arslan-Acaroz D, Istanbullugil FR, Madani K, et al. Natural Strategies as Potential Weapons against Bacterial Biofilms. Life. 2022; 12(10):1618. https://doi.org/10.3390/life12101618
Chicago/Turabian StyleAsma, Syeda Tasmia, Kálmán Imre, Adriana Morar, Mirela Imre, Ulas Acaroz, Syed Rizwan Ali Shah, Syed Zajif Hussain, Damla Arslan-Acaroz, Fatih Ramazan Istanbullugil, Khodir Madani, and et al. 2022. "Natural Strategies as Potential Weapons against Bacterial Biofilms" Life 12, no. 10: 1618. https://doi.org/10.3390/life12101618