

Effect of Salinity and Plant Growth Promoters on Secondary Metabolism and Growth of Milk Thistle Ecotypes
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Details
2.2. Growth and Yield Indicators
2.3. Determination of Secondary Metabolites
2.4. Anthocyanin Determination
2.5. Total Alkaloids Determination
2.6. Saponin Estimation
2.7. Statistical Analysis
3. Results
3.1. Plant Length
3.2. Number of Leaves
3.3. Leaf Area
3.4. Fresh Weight
3.5. Dry Weight
3.6. Number of Roots
3.7. Number of Spines
3.8. Root Diameter
3.9. Relative Water Content
3.10. Saponin Content
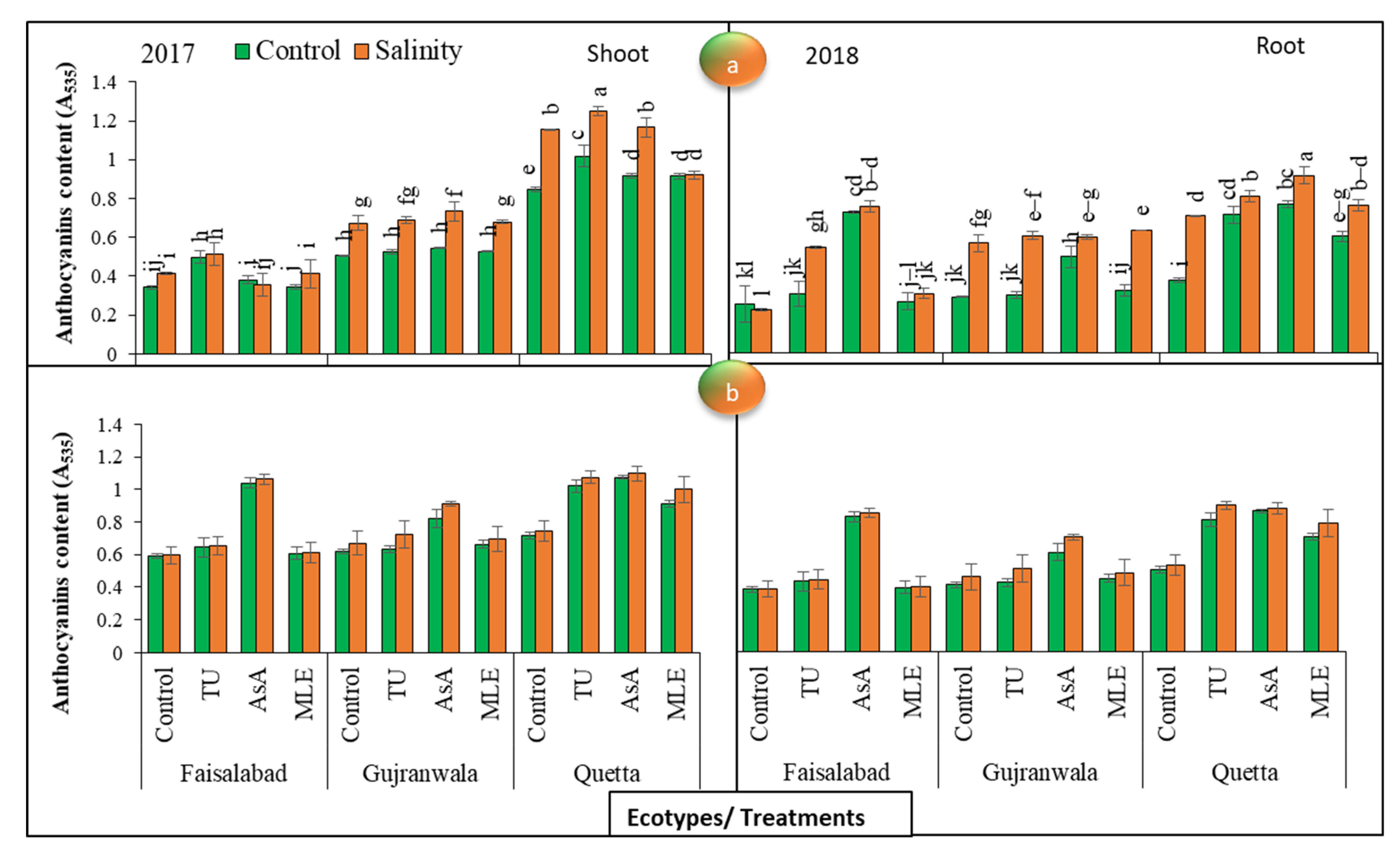
3.11. Anthocyanin Content
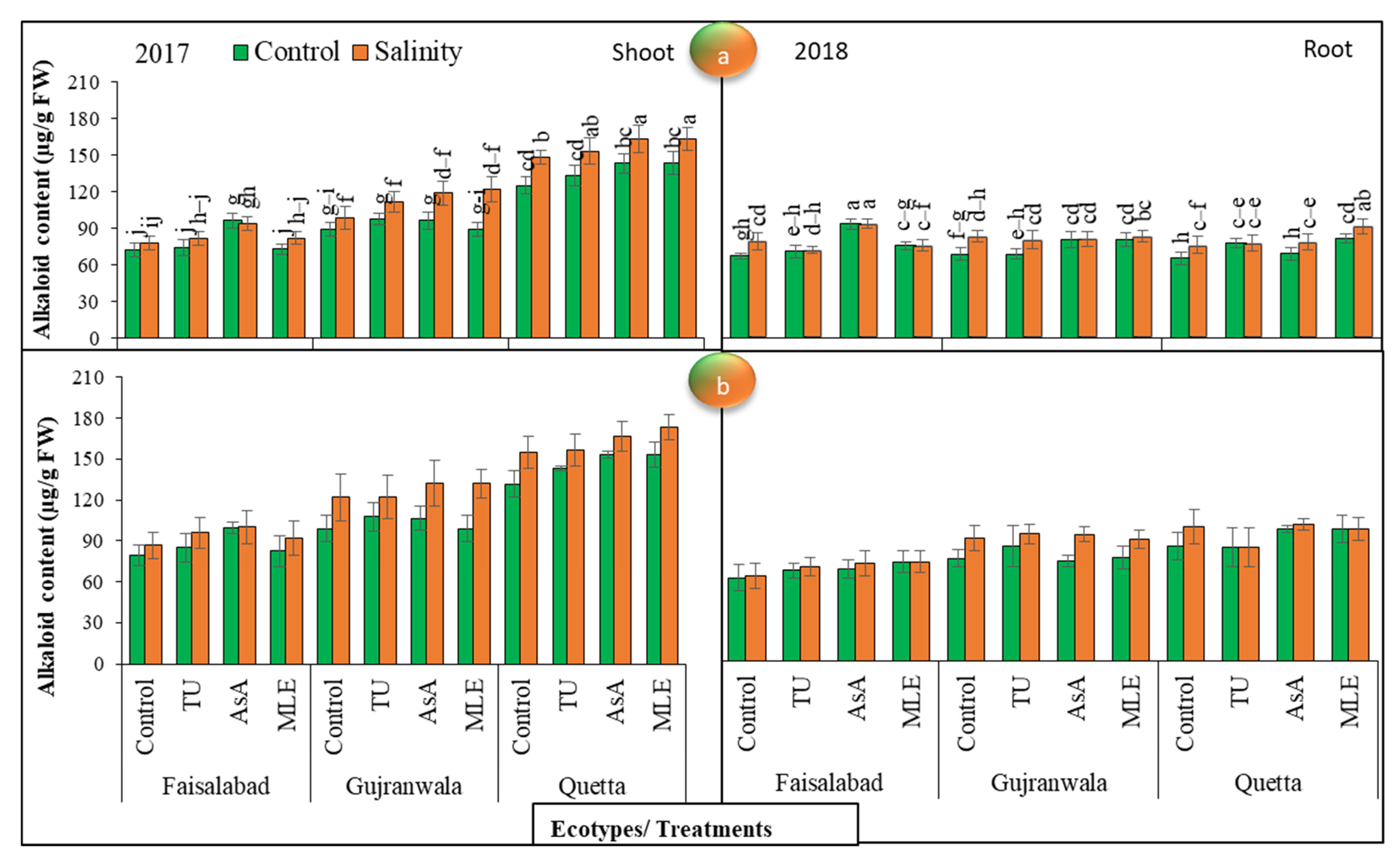
3.12. Alkaloids Content
3.13. Principle Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shah, M.; Jan, H.; Drouet, S.; Tungmunnithum, D.; Shirazi, J.H.; Hano, C.; Abbasi, B.H. Chitosan Elicitation Impacts Flavonolignan Biosynthesis in Silybum marianum (L.) Gaertn Cell Suspension and Enhances Antioxidant and Anti-Inflammatory Activities of Cell Extracts. Molecules 2021, 26, 791. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.-C.; Graf, T.N.; Sparacino, C.M.; Wani, M.C.; Wall, M.E. Complete isolation and characterization of silybins and isosilybins from milk thistle (Silybum marianum). Org. Biomol. Chem. 2003, 1, 1684–1689. [Google Scholar] [CrossRef] [PubMed]
- Denev, P.; Ognyanov, M.; Georgiev, Y.; Teneva, D.; Klisurova, D.; Yanakieva, I.Z. Chemical composition and antioxidant activity of partially defatted milk thistle (Silybum marianum L.) seeds. Bulg. Chem. Commun. 2020, 52, 182–187. [Google Scholar]
- Choe, U.; Whent, M.; Luo, Y.; Yu, L. Total phenolic content, free radical scavenging capacity, and anti-cancer activity of silymarin. J. Food Bioact. 2020, 10, 53–64. [Google Scholar] [CrossRef]
- Abenavoli, L.; Izzo, A.A.; Milić, N.; Cicala, C.; Santini, A.; Capasso, R. Milk thistle (Silybum marianum): A concise overview on its chemistry, pharmacological, and nutraceutical uses in liver diseases. Phytother. Res. 2018, 32, 2202–2213. [Google Scholar] [CrossRef] [PubMed]
- Kogo, B.K.; Kumar, L.; Koech, R. Climate change and variability in Kenya: A review of impacts on agriculture and food security. Environ. Develop. Sustain. 2020, 23, 23–43. [Google Scholar] [CrossRef]
- Hafeez, M.B.; Raza, A.; Zahra, N.; Shaukat, K.; Akram, M.Z.; Iqbal, S.; Basra, S.M.A. Gene regulation in halophytes in conferring salt tolerance. In Handbook of Bioremediation; Elsevier: Amsterdam, The Netherlands, 2021; pp. 341–370. [Google Scholar]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. The role of endogenous nitric oxide in salicylic acid-induced up-regulation of ascorbate-glutathione cycle involved in salinity tolerance of pepper (Capsicum annuum L.) plants. Plant Physiol. Biochem. 2020, 147, 10–20. [Google Scholar] [CrossRef]
- Zahra, N.; Hafeez, M.B.; Shaukat, K.; Wahid, A.; Hussain, S.; Naseer, R.; Raza, A.; Iqbal, S.; Farooq, M. Hypoxia and Anoxia Stress: Plant responses and tolerance mechanisms. J. Agron. Crop Sci. 2021, 207, 249–284. [Google Scholar] [CrossRef]
- Mubarik, M.S.; Khan, S.H.; Sajjad, M.; Raza, A.; Hafeez, M.B.; Yasmeen, T.; Rizwan, M.; Ali, S.; Arif, M.S. A manipulative interplay between positive and negative regulators of phytohormones: A way forward for improving drought tolerance in plants. Physiol. Plant. 2021, 172, 1269–1290. [Google Scholar] [CrossRef]
- Batool, S.; Khan, S.; Basra, S.M.; Hussain, M.; Saddiq, M.S.; Iqbal, S.; Irshad, S.; Hafeez, M. Impact of natural and synthetic plant stimulants on Moringa seedlings grown under low-temperature conditions. Int. Lett. Nat. Sci. 2019, 76, 50–59. [Google Scholar] [CrossRef]
- Akhter, M.S.; Noreen, S.; Mahmood, S.; Ashraf, M.; Alsahli, A.A.; Ahmad, P. Influence of salinity stress on PSII in barley (Hordeum vulgare L.) genotypes, probed by chlorophyll-a fluorescence. J. King Saud Univ. Sci. 2021, 33, 101239. [Google Scholar] [CrossRef]
- Ahmad, P.; Venema, K.; Corpas, F.J. Unravelling salt tolerance mechanisms in plants: From Lab to Field. Plant Physiol. Biochem. 2022, 176, 31–33. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; Li, Y.; Li, X.; Han, W.-Y.; Chen, S. Epigallocatechin-3-gallate alleviates salinity-retarded seed germination and oxidative stress in tomato. J. Plant Growth Regul. 2018, 37, 1349–1356. [Google Scholar] [CrossRef]
- Shah, T.; Latif, S.; Saeed, F.; Ali, I.; Ullah, S.; Alsahli, A.A.; Jan, S.; Ahmad, P. Seed priming with titanium dioxide nanoparticles enhances seed vigor, leaf water status, and antioxidant enzyme activities in maize (Zea mays L.) under salinity stress. J King Saud Uni. Sci. 2020, 33, 101207. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Mir, R.A.; Alyemeni, M.N.; Ahmad, P. Combined effects of brassinosteroid and kinetin mitigates salinity stress in tomato through the modulation of antioxidant and osmolyte metabolism. Plant Physiol. Biochem. 2020, 147, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Zahra, N.; Raza, Z.A.; Mahmood, S. Effect of salinity stress on various growth and physiological attributes of two contrasting maize genotypes. Brazil. Arch. Biol. Technol. 2020, 63, e20200072. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Saddiq, M.S.; Iqbal, S.; Hafeez, M.B.; Ibrahim, A.M.; Raza, A.; Fatima, E.M.; Baloch, H.; Woodrow, P.; Ciarmiello, L.F. Effect of salinity stress on physiological changes in winter and spring wheat. Agronomy 2021, 11, 1193. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Ahmad, I.; Basit, A.; El-Lateef, A.; Hany, M.; Yasir, M.; Tanveer Shah, S.; Ullah, I.; Elsayed Mohamed Mohamed, M.; Ali, I. Effect of azospirillum and azotobacter species on the performance of cherry tomato under different salinity levels. Gesunde Pflanz. 2022, 74, 487–499. [Google Scholar] [CrossRef]
- Ismail, L.M.; Soliman, M.I.; Abd El-Aziz, M.H.; Abdel-Aziz, H.M. Impact of silica ions and nano silica on growth and productivity of pea plants under salinity stress. Plants 2022, 11, 494. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, X.; Yan, J.; Yuan, Z.; Gu, M. Effects of salt stress on growth, photosynthesis, and mineral nutrients of 18 pomegranate (Punica granatum) cultivars. Agronomy 2020, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Khademian, R.; Asghari, B.; Sedaghati, B.; Yaghoubian, Y. Plant beneficial rhizospheric microorganisms (PBRMs) mitigate deleterious effects of salinity in sesame (Sesamum indicum L.): Physio-biochemical properties, fatty acids composition and secondary metabolites content. Ind. Crops Prod. 2019, 136, 129–139. [Google Scholar] [CrossRef]
- MacNeill, G.J.; Mehrpouyan, S.; Minow, M.A.; Patterson, J.A.; Tetlow, I.J.; Emes, M.J.; Raines, C. Starch as a source, starch as a sink: The bifunctional role of starch in carbon allocation. J. Exp. Bot. 2017, 68, 4433–4453. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Khaliq, A.; Zia-ul-Haq, M.; Ali, F.; Aslam, F.; Matloob, A.; Navab, A.; Hussain, S. Salinity tolerance in wheat cultivars is related to enhanced activities of enzymatic antioxidants and reduced lipid peroxidation. CLEAN–Soil Air Water 2015, 43, 1248–1258. [Google Scholar] [CrossRef]
- Kamiab, F. Exogenous melatonin mitigates the salinity damages and improves the growth of pistachio under salinity stress. J. Plant Nutr. 2020, 43, 1468–1484. [Google Scholar] [CrossRef]
- Ishaq, H.; Nawaz, M.; Azeem, M.; Mehwish, M.; Naseem, M.B.B. Ascorbic Acid (Asa) improves Salinity Tolerance in Wheat (Triticum Aestivum L.) by Modulating Growth and Physiological Attributes. J. Bioresour. Manag. 2021, 8, 1. [Google Scholar] [CrossRef]
- Ahmadi, F.; Karimi, K.; Struik, P. Effect of exogenous application of methyl jasmonate on physiological and biochemical characteristics of Brassica napus L. cv. Talaye under salinity stress. S. Afr. J. Bot. 2018, 115, 5–11. [Google Scholar] [CrossRef]
- Alam, P.; Albalawi, T.H.; Altalayan, F.H.; Bakht, M.A.; Ahanger, M.A.; Raja, V.; Ashraf, M.; Ahmad, P. 24-Epibrassinolide (EBR) confers tolerance against NaCl stress in soybean plants by up-regulating antioxidant system, ascorbate-glutathione cycle, and glyoxalase system. Biomolecules 2019, 9, 640. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Aziz, U.; Alsahli, A.A.; Alyemeni, M.N.; Ahmad, P. Influence of exogenous salicylic acid and nitric oxide on growth, photosynthesis, and ascorbate-glutathione cycle in salt stressed Vigna angularis. Biomolecules 2020, 10, 42. [Google Scholar] [CrossRef]
- Zheng, Y.; Xu, B.; Ren, K.; Zhang, Y.; Wu, J. Impact of soil drench and foliar spray of 24-epibrassinolide on the growth, yield, and quality of field-grown Moringa oleifera in Southwest China. J. Plant Growth Regul. 2017, 36, 931–941. [Google Scholar] [CrossRef]
- Waqas, M.; Yaning, C.; Iqbal, H.; Shareef, M.; ur Rehman, H.; Iqbal, S.; Mahmood, S. Soil drenching of paclobutrazol: An efficient way to improve quinoa performance under salinity. Physiol. Plant. 2019, 165, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.; Bakheta, M.; Zaki, S. Influence of uniconazole on growth characters, photosynthetic pigments, total carbohydrates and total soluble sugars of Hordium vulgare L. plants grown under salinity stress. Int. J. Sci. Res. 2014, 3, 2208–2213. [Google Scholar]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic acid-a potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Zhang, G.; Chen, Y.; Gao, J.; Sun, Y.-R.; Sun, M.-F.; Chen, J.-P. Exogenous application of gibberellic acid and ascorbic acid improved tolerance of okra seedlings to NaCl stress. Acta Physiol. Plant. 2019, 41, 93. [Google Scholar] [CrossRef]
- de Sousa Basílio, A.G.; Vieira de Sousa, L.; da Silva, T.I.; de Moura, J.G.; de Melo Gonçalves, A.C.; de Melo Filho, J.S.; Leal, Y.H.; Jardelino Dias, T. Radish (Raphanus sativus L.) morphophysiology under salinity stress and ascorbic acid treatments. Agron. Colomb. 2018, 36, 257–265. [Google Scholar] [CrossRef]
- Waqas, M.A.; Kaya, C.; Riaz, A.; Farooq, M.; Nawaz, I.; Wilkes, A.; Li, Y. Potential mechanisms of abiotic stress tolerance in crop plants induced by thiourea. Front. Plant Sci. 2019, 10, 1336. [Google Scholar] [CrossRef]
- Aziz, U.; Qadir, I.; Yasin, G.; Azhar, M.F.; Javed, A.; Akhtar, A. Potential of priming in improving germination, seedling growth and nutrient status of Calotropis procera under salinity. Pak. J. Bot. 2021, 53, 1953–1958. [Google Scholar] [CrossRef]
- Yaseen, A.; Takacsne Hajos, M. Study on moringa tree (Moringa oleifera Lam.) leaf extract in organic vegetable production: A review. Res. Crops 2020, 21, 402–414. [Google Scholar]
- Faisal, M.; Iqbal, S.; Basra, S.; Afzal, I.; Saddiq, M.; Bakhtavar, M.; Hafeez, M.; Rehman, H.; Basit, A.; Habib-ur-Rahman, M. Moringa landraces of Pakistan are potential source of premium quality oil. S. Afr. J. Bot. 2020, 129, 397–403. [Google Scholar] [CrossRef]
- Aslam, M.F.; Basra, S.M.; Hafeez, M.B.; Khan, S.; Irshad, S.; Iqbal, S.; Saqqid, M.S.; Akram, M.Z. Inorganic fertilization improves quality and biomass of Moringa oleifera L. Agrofor. Syst. 2020, 94, 975–983. [Google Scholar] [CrossRef]
- Ahmed, T.; Abou Elezz, A.; Khalid, M.F. Hydropriming with moringa leaf extract mitigates salt stress in Wheat seedlings. Agriculture 2021, 11, 1254. [Google Scholar] [CrossRef]
- Yaghoubian, I.; Antar, M.; Ghassemi, S.; Modarres-Sanavy, S.A.M.; Smith, D.L. The Effects of Hydro-Priming and Colonization with Piriformospora indica and Azotobacter chroococcum on Physio-Biochemical Traits, Flavonolignans and Fatty Acids Composition of Milk Thistle (Silybum marianum) under Saline Conditions. Plants 2022, 11, 1281. [Google Scholar] [CrossRef] [PubMed]
- Perveen, A.; Wahid, A.; Mahmood, S.; Hussain, I.; Rasheed, R. Possible mechanism of medium-supplemented thiourea in improving growth, gas exchange, and photosynthetic pigments in cadmium-stressed maize (Zea mays). Brazil. J. Bot. 2015, 38, 71–79. [Google Scholar] [CrossRef]
- Rashid, N.; Basra, S.M.; Shahbaz, M.; Iqbal, S.; Hafeez, M.B. Foliar applied moringa leaf extract induces terminal heat tolerance in quinoa. Int. J. Agric. Biol. 2018, 20, 157–164. [Google Scholar]
- Peñas, E.; Limón, R.I.; Martínez-Villaluenga, C.; Restani, P.; Pihlanto, A.; Frias, J. Impact of elicitation on antioxidant and potential antihypertensive properties of lentil sprouts. Plant Foods Human Nutr. 2015, 70, 401–407. [Google Scholar] [CrossRef]
- STRACK, D.; WRAY, V. Anthocyanins. In Methods in Plant Biochemistry; Elsevier: Amsterdam, The Netherlands, 1989; Volume 1, pp. 325–356. [Google Scholar]
- Singh, D.; Sahu, A. Spectrophotometric determination of caffeine and theophylline in pure alkaloids and its application in pharmaceutical formulations. Anal. Biochem. 2006, 349, 176–180. [Google Scholar] [CrossRef]
- Marino, D.; González, E.M.; Arrese-Igor, C. Drought effects on carbon and nitrogen metabolism of pea nodules can be mimicked by paraquat: Evidence for the occurrence of two regulation pathways under oxidative stresses. J. Exp. Bot. 2006, 57, 665–673. [Google Scholar] [CrossRef]
- Su, Y.; Guo, A.; Huang, Y.; Wang, Y.; Hua, J. GhCIPK6a increases salt tolerance in transgenic upland cotton by involving in ROS scavenging and MAPK signaling pathways. BMC Plant Biol. 2020, 20, 421. [Google Scholar] [CrossRef]
- Chartzoulakis, K.; Klapaki, G. Response of two greenhouse pepper hybrids to NaCl salinity during different growth stages. Sci. Horti. 2000, 86, 247–260. [Google Scholar] [CrossRef]
- Kurban, H.; Saneoka, H.; Nehira, K.; Adilla, R.; Premachandra, G.S.; Fujita, K. Effect of salinity on growth, photosynthesis and mineral composition in leguminous plant Alhagi pseudoalhagi (Bieb.). Soil Sci. Plant Nutr. 1999, 45, 851–862. [Google Scholar] [CrossRef]
- Ghavami, N.; Ramin, A. Grain yield and active substances of milk thistle as affected by soil salinity. Comm. Soil Sci. Plant Anal. 2008, 39, 2608–2618. [Google Scholar] [CrossRef]
- Arif, M.R.; Islam, M.T.; Robin, A.H.K. Salinity stress alters root morphology and root hair traits in Brassica napus. Plants 2019, 8, 192. [Google Scholar] [CrossRef] [PubMed]
- Robin, A.H.K.; Matthew, C.; Uddin, M.J.; Bayazid, K.N. Salinity-induced reduction in root surface area and changes in major root and shoot traits at the phytomer level in wheat. J. Exp. Bot. 2016, 67, 3719–3729. [Google Scholar] [CrossRef] [PubMed]
- Sahu, M.; Solanki, N.; Dashora, L. Effects of thiourea, thiamine and ascorbic acid on growth and yield of maize (Zea mays L.). J. Agron. Crop Sci. 1993, 171, 65–69. [Google Scholar] [CrossRef]
- Waqas, M.A.; Khan, I.; Akhter, M.J.; Noor, M.A.; Ashraf, U. Exogenous application of plant growth regulators (PGRs) induces chilling tolerance in short-duration hybrid maize. Environ. Sci. Pollut. Res. 2017, 24, 11459–11471. [Google Scholar] [CrossRef]
- Khan, A.; Ashraf, M. Exogenously applied ascorbic acid alleviates salt-induced oxidative stress in wheat. Environ. Exp. Bot. 2008, 63, 224–231. [Google Scholar]
- Gengmao, Z.; Yu, H.; Xing, S.; Shihui, L.; Quanmei, S.; Changhai, W. Salinity stress increases secondary metabolites and enzyme activity in safflower. Ind. Crops Prod. 2015, 64, 175–181. [Google Scholar] [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Comm. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Mbarki, S.; Sytar, O.; Zivcak, M.; Abdelly, C.; Cerda, A.; Brestic, M. Anthocyanins of Coloured wheat genotypes in specific response to SalStress. Molecules 2018, 23, 1518. [Google Scholar] [CrossRef]
- Eryılmaz, F. The relationships between salt stress and anthocyanin content in higher plants. Biotechnol. Biotechnol. Equip. 2006, 20, 47–52. [Google Scholar] [CrossRef]
- Yahyazadeh, M.; Meinen, R.; Hänsch, R.; Abouzeid, S.; Selmar, D. Impact of drought and salt stress on the biosynthesis of alkaloids in Chelidonium majus L. Phytochemistry 2018, 152, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Idrees, M.; Naeem, M.; Aftab, T.; Khan, M.M.A. Salicylic acid mitigates salinity stress by improving antioxidant defence system and enhances vincristine and vinblastine alkaloids production in periwinkle [Catharanthus roseus (L.) G. Don]. Acta Physiol. Plant. 2011, 33, 987–999. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Leelapriya, T.; Kumari, B.D.R. Effects of pulsed magnetic field treatment of soybean seeds on calli growth, cell damage, and biochemical changes under salt stress. Bioelectromagnetics 2012, 33, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Gaafar, A.A.; Ali, S.I.; El-Shawadfy, M.A.; Salama, Z.A.; Sekara, A.; Ulrichs, C.; Abdelhamid, M.T. Ascorbic acid induces the increase of secondary metabolites, antioxidant activity, growth, and productivity of the common bean under water stress conditions. Plants 2020, 9, 627. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zahra, N.; Wahid, A.; Hafeez, M.B.; Lalarukh, I.; Batool, A.; Uzair, M.; El-Sheikh, M.A.; Alansi, S.; Kaushik, P. Effect of Salinity and Plant Growth Promoters on Secondary Metabolism and Growth of Milk Thistle Ecotypes. Life 2022, 12, 1530. https://doi.org/10.3390/life12101530
Zahra N, Wahid A, Hafeez MB, Lalarukh I, Batool A, Uzair M, El-Sheikh MA, Alansi S, Kaushik P. Effect of Salinity and Plant Growth Promoters on Secondary Metabolism and Growth of Milk Thistle Ecotypes. Life. 2022; 12(10):1530. https://doi.org/10.3390/life12101530
Chicago/Turabian StyleZahra, Noreen, Abdul Wahid, Muhammad Bilal Hafeez, Irfana Lalarukh, Aaliya Batool, Muhammad Uzair, Mohamed A. El-Sheikh, Saleh Alansi, and Prashant Kaushik. 2022. "Effect of Salinity and Plant Growth Promoters on Secondary Metabolism and Growth of Milk Thistle Ecotypes" Life 12, no. 10: 1530. https://doi.org/10.3390/life12101530