

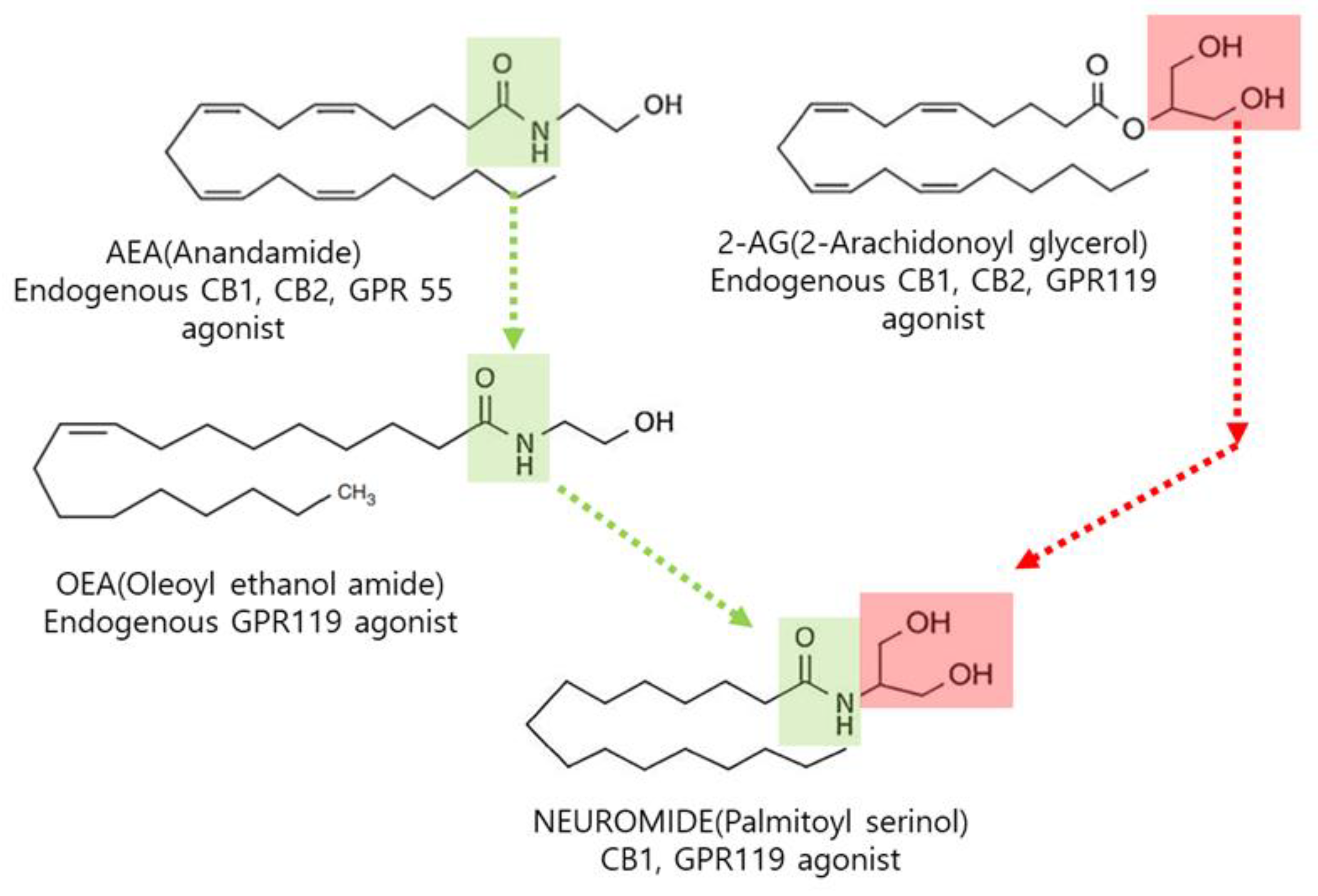
The Neuroprotective Effect of NEUROMIDE, a Compound Bioidentical to Commensal Bacteria Metabolites
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Viability Test
2.3. Induction of Cell Damage
2.4. Cell Viability Test
2.5. Measurement of Acetylcholine (Ach) in PC-12 Cells
2.6. RNA Preparation and qRT-PCR
2.7. Animal
2.8. Morris Water Maze
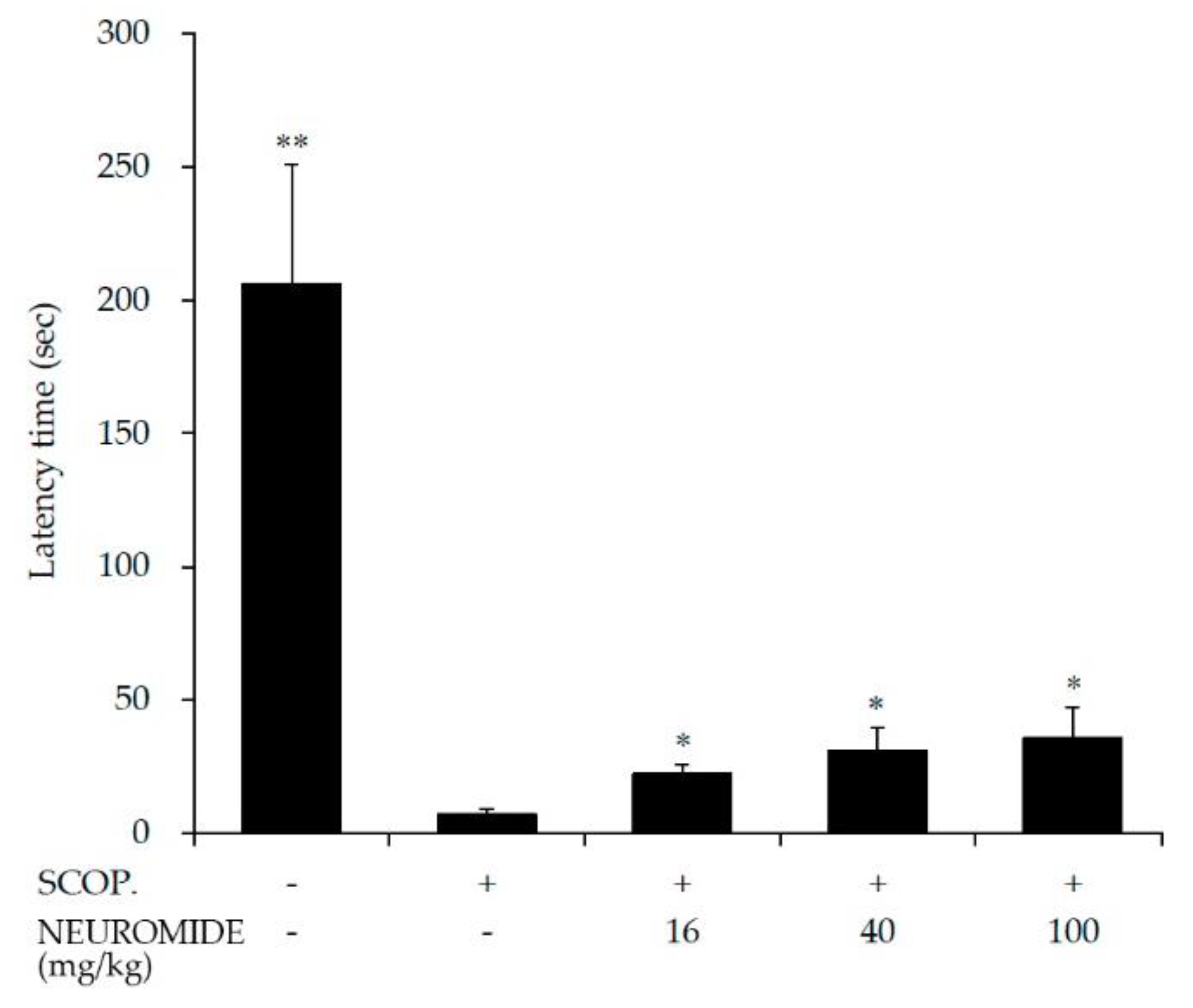
2.9. Passive Avoidance Test
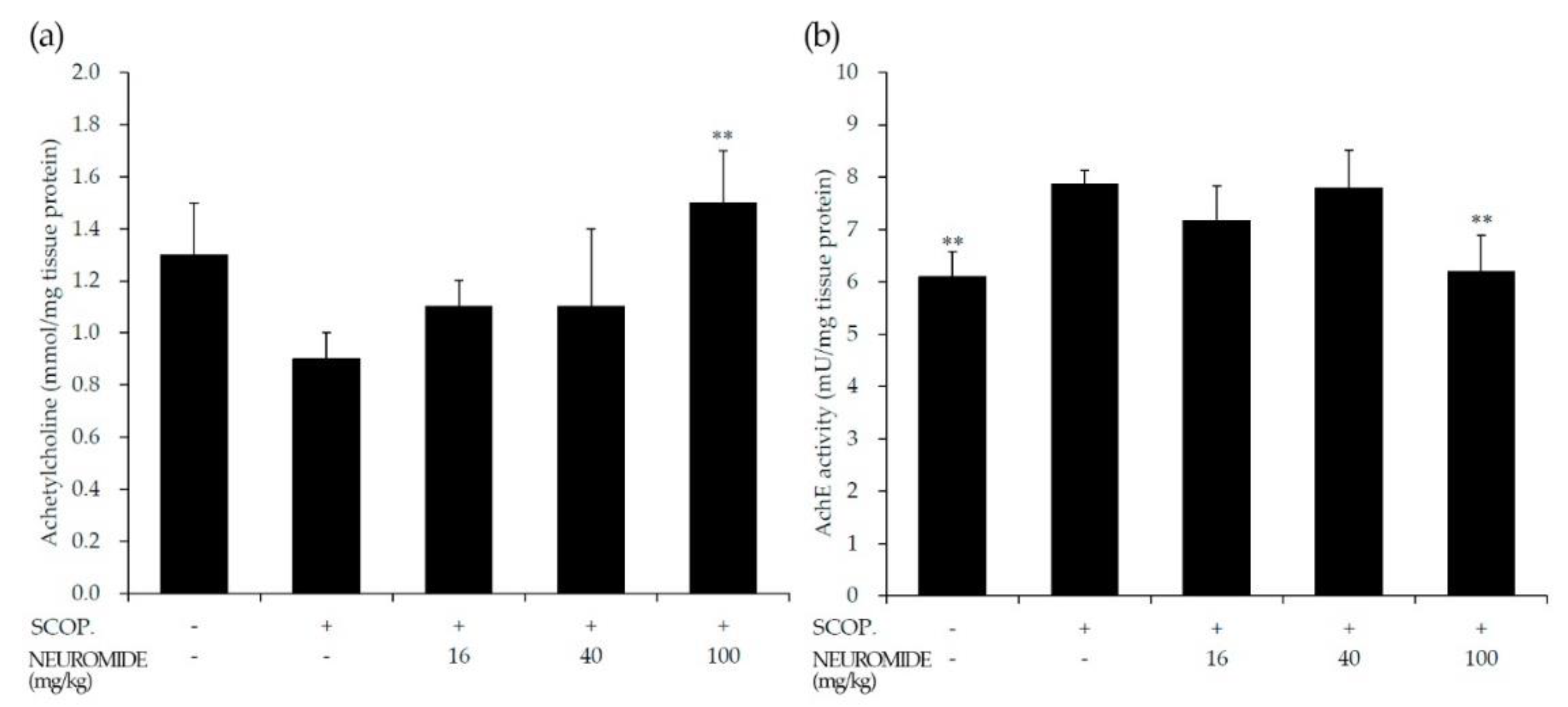
2.10. Measurement of Acetylcholine and Acetylcholine Esterase Activity in Brain Tissue
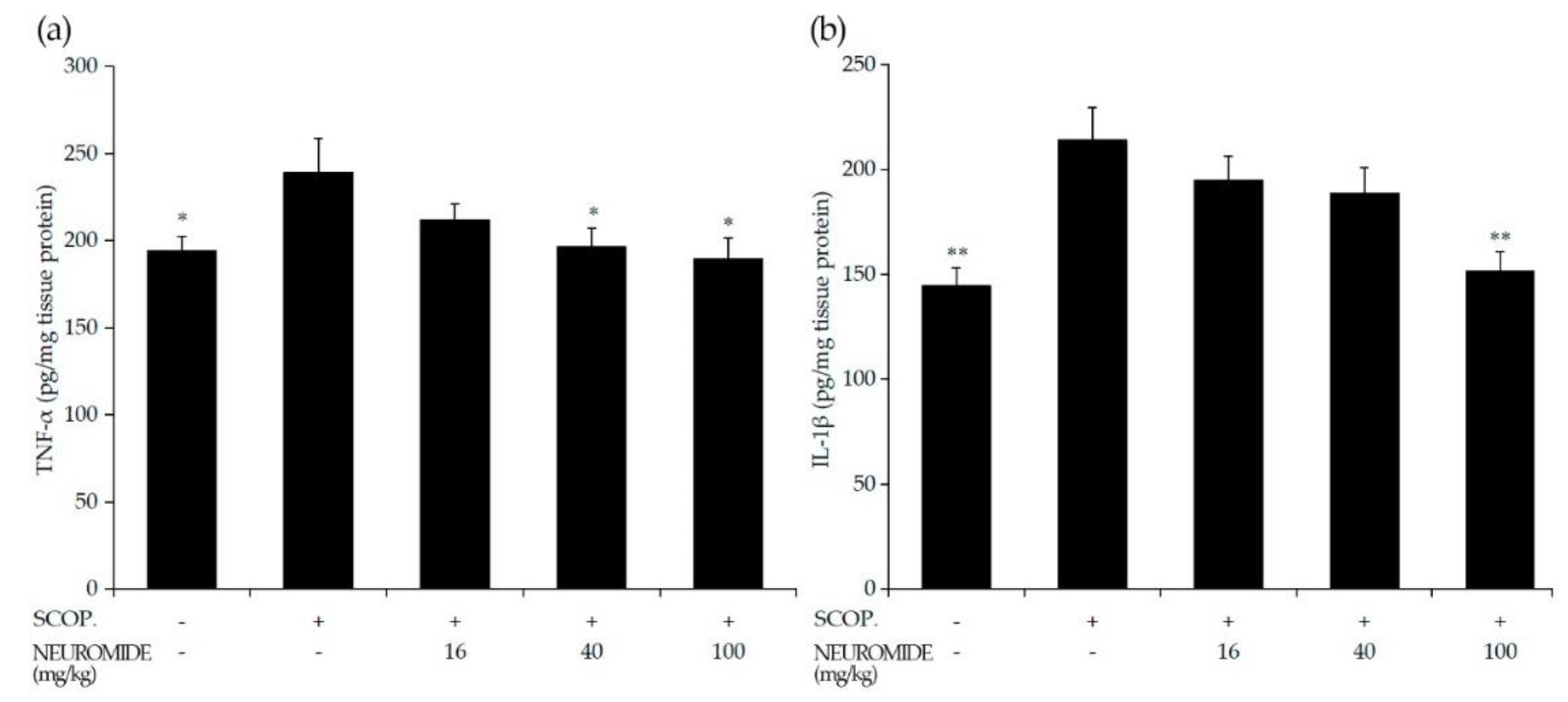
2.11. Measurement of Cytokines in Brain Tissue
2.12. Statistical Analysis
3. Results
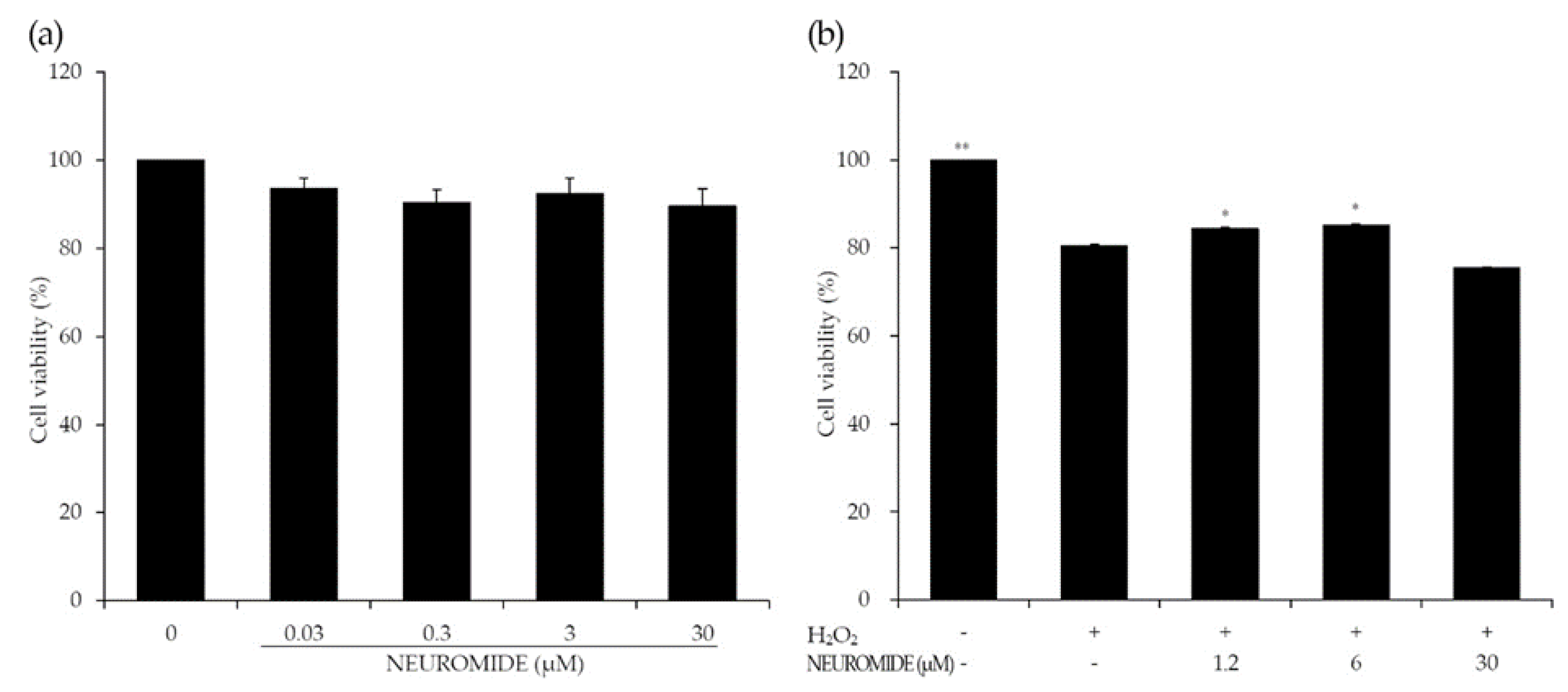
3.1. NEUROMIDE Protects Neuronal Cells from Oxidative Stress
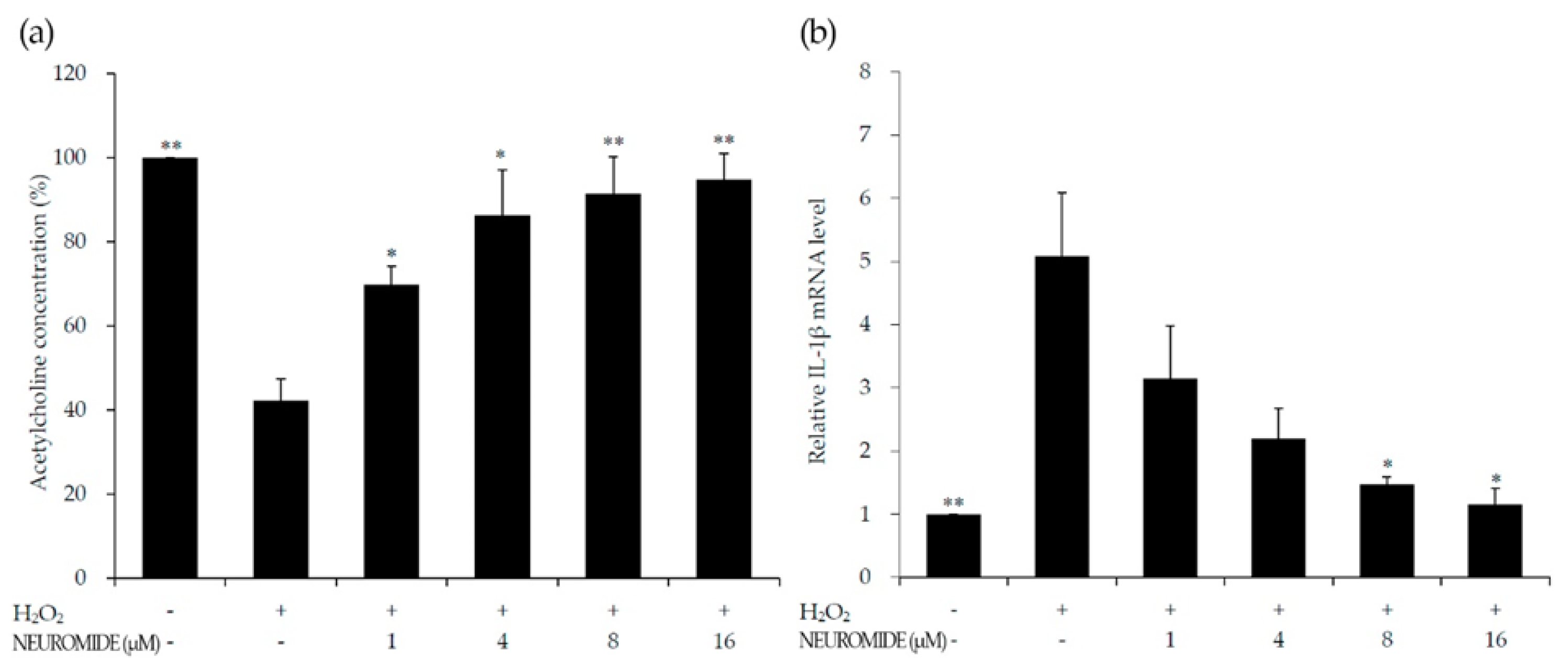
3.2. Effects of NEUROMIDE on Acetylcholine and Cytokines in PC-12 Cells
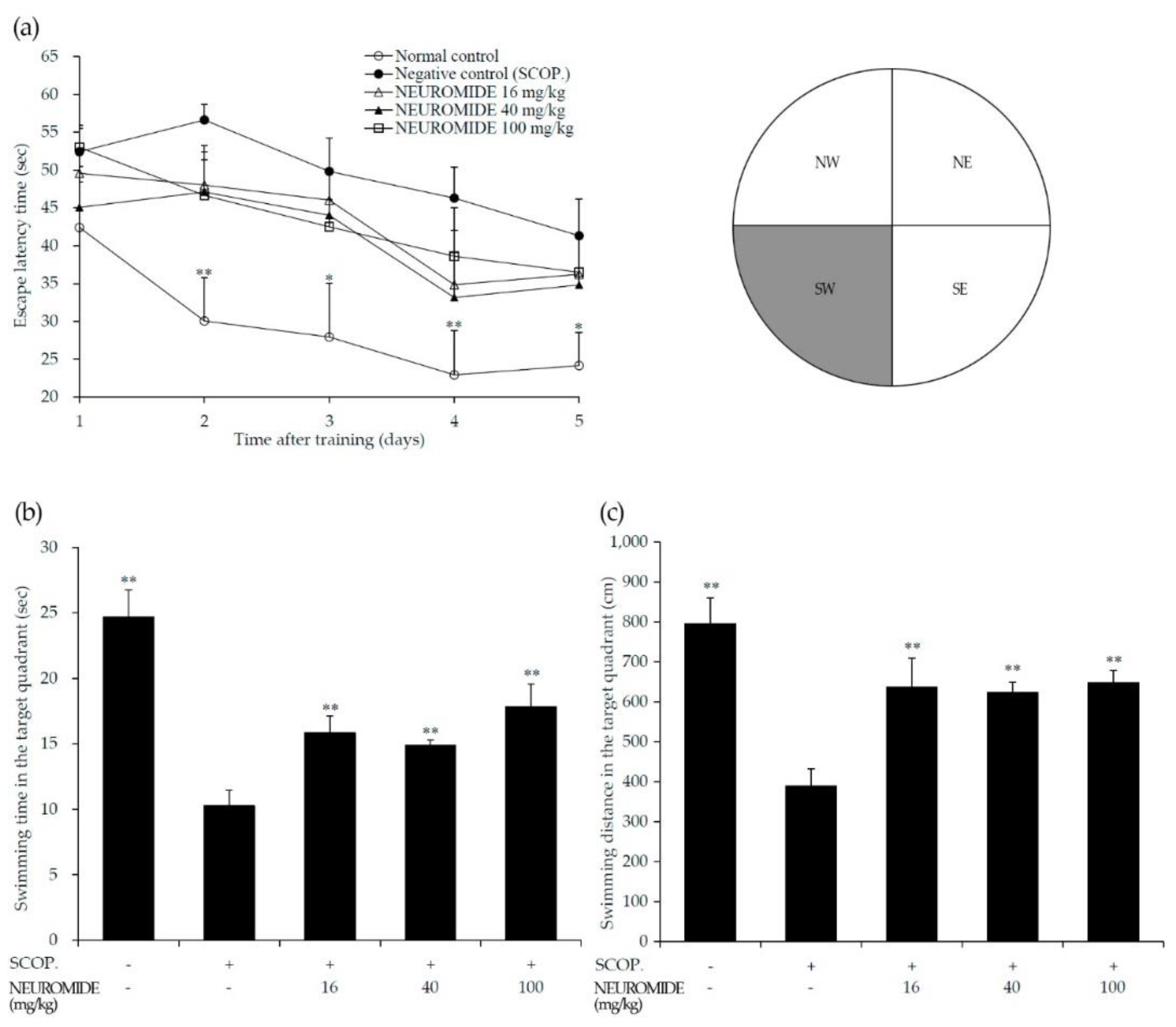
3.3. Effect of NEUROMIDE on Improving Spatial Memory
3.4. Effect of NEUROMIDE on Improvements in Learning and Memory
3.5. Effect of NEUROMIDE on Acetylcholine and Acetylcholine Esterase in the Forebrain
3.6. Effect of NEUROMIDE on Inflammatory Cytokines in the Hippocampus
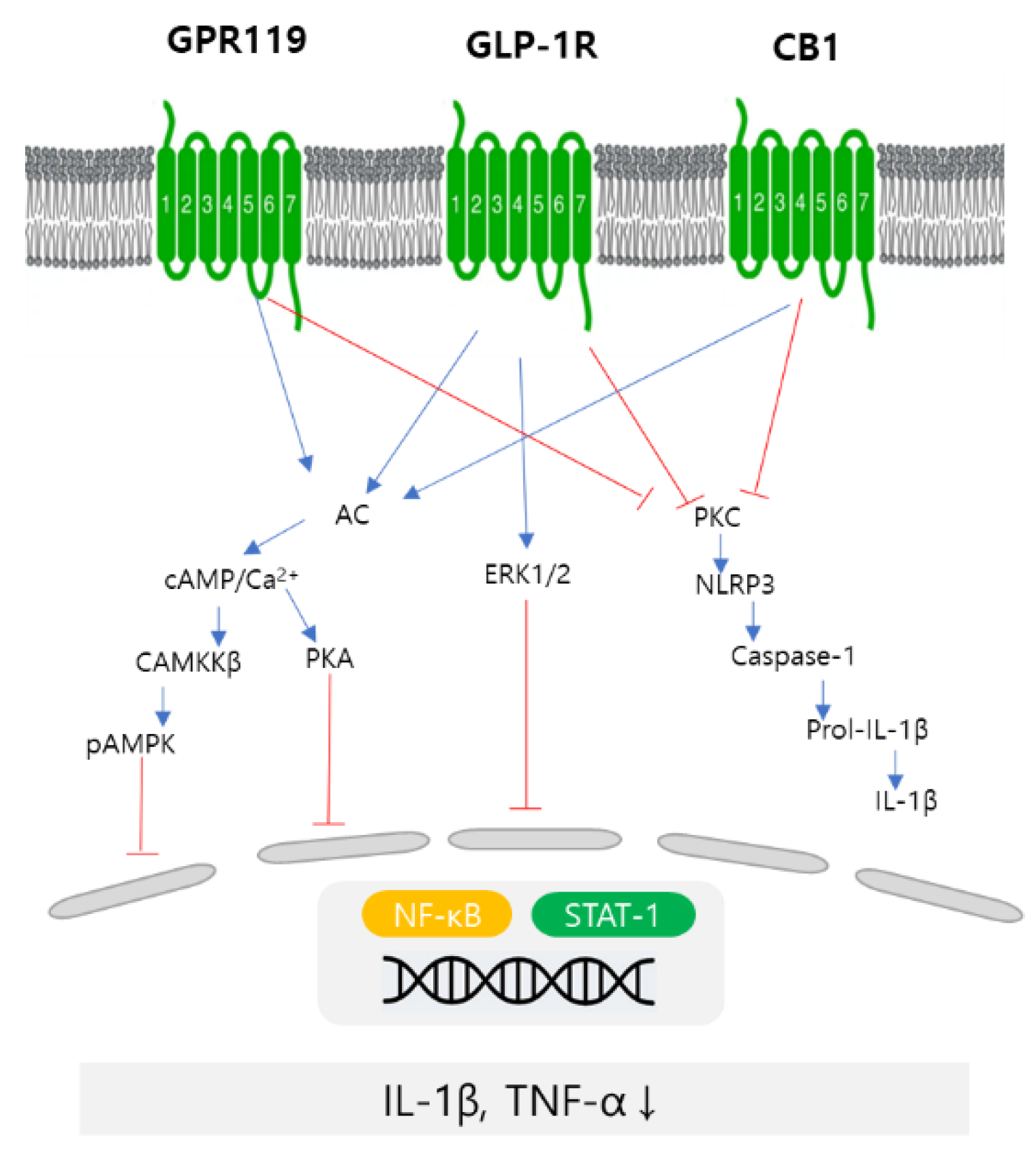
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef]
- Sharir, H.; Console-Bram, L.; Mundy, C.; Popoff, S.N.; Kapur, A.; Abood, M.E. The Endocannabinoids Anandamide and Vi-rodhamine Modulate the Activity of the Candidate Cannabinoid Receptor GPR55. J. Neuroimmune Pharmacol. 2012, 7, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Lauffer, L.M.; Iakoubov, R.; Brubaker, P.L. GPR119 Is Essential for Oleoylethanolamide-Induced Glucagon-Like Peptide-1 Secretion from the Intestinal Enteroendocrine L-Cell. Diabetes 2009, 58, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Gonsiorek, W.; Lunn, C.; Fan, X.; Narula, S.; Lundell, D.; Hipkin, R.W. Endocannabinoid 2-arachidonyl glycerol is a full agonist through human type 2 cannabinoid receptor: Antagonism by anandamide. Mol. Pharmacol. 2000, 57, 1045–1050. [Google Scholar] [PubMed]
- Hansen, K.B.; Rosenkilde, M.M.; Knop, F.K.; Wellner, N.; Diep, T.A.; Rehfeld, J.F.; Andersen, U.B.; Holst, J.J.; Hansen, H.S. 2-Oleoyl Glycerol Is a GPR119 Agonist and Signals GLP-1 Release in Humans. J. Clin. Endocrinol. Metab. 2011, 96, E1409–E1417. [Google Scholar] [CrossRef]
- Available online: https://hmdb.ca/metabolites/HMDB0013654 (accessed on 7 January 2022).
- Available online: https://foodb.ca/compounds/FDB029624 (accessed on 7 January 2022).
- Bieberich, E.; Kawaguchi, T.; Yu, R.K. N-Acylated Serinol Is a Novel Ceramide Mimic Inducing Apoptosis in Neuroblastoma Cells. J. Biol. Chem. 2000, 275, 177–181. [Google Scholar] [CrossRef]
- Cohen, L.J.; Esterházy, D.; Kim, S.-H.; Lemetre, C.; Aguilar, R.R.; Gordon, E.A.; Pickard, A.J.; Cross, J.R.; Emiliano, A.B.; Han, S.M.; et al. Corrigendum: Commensal bacteria make GPCR ligands that mimic human signalling molecules. Nature 2018, 556, 135. [Google Scholar] [CrossRef]
- Zhao, J.; Zhao, Y.; Hu, Y.; Peng, J. Targeting the GPR119/incretin axis: A promising new therapy for metabolic-associated fatty liver disease. Cell. Mol. Biol. Lett. 2021, 26, 32. [Google Scholar] [CrossRef]
- Im, D.-S. GPR119 and GPR55 as Receptors for Fatty Acid Ethanolamides, Oleoylethanolamide and Palmitoylethanolamide. Int. J. Mol. Sci. 2021, 22, 1034. [Google Scholar] [CrossRef]
- Suardíaz, M.; Estivill-Torrús, G.; Goicoechea, C.; Bilbao, A.; Rodríguez de Fonseca, F. Analgesic properties of oleoylethanola-mide (OEA) in visceral and inflammatory pain. Pain 2007, 133, 99–110. [Google Scholar] [CrossRef]
- Tough, I.R.; Forbes, S.; Herzog, H.; Jones, R.M.; Schwartz, T.W.; Cox, H.M. Bidirectional GPR119 Agonism Requires Peptide YY and Glucose for Activity in Mouse and Human Colon Mucosa. Endocrinology 2018, 159, 1704–1717. [Google Scholar] [CrossRef] [PubMed]
- Cully, M. Gut feeling on bacterial GPCR agonists. Nat. Rev. Drug Discov. 2017, 16, 754. [Google Scholar] [CrossRef] [PubMed]
- Markovics, A.; Angyal, Á.; Tóth, K.F.; Ádám, D.; Pénzes, Z.; Magi, J.; Pór, Á.; Kovács, I.; Törőcsik, D.; Zouboulis, C.C.; et al. GPR119 Is a Potent Regulator of Human Sebocyte Biology. J. Investig. Dermatol. 2020, 140, 1909–1918.e8. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.A.; Jacobs, S.E.; Pentland, A.P. sPLA2-X Stimulates Cutaneous Melanocyte Dendricity and Pigmentation Through a Lysophosphatidylcholine-Dependent Mechanism. J. Investig. Dermatol. 2006, 126, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Kim, M.S.; Lee, S.H.; Park, B.D. Epidermal Endocannabinoid System (EES) and its Cosmetic Application. Cosmetics 2019, 6, 33. [Google Scholar] [CrossRef]
- Wen, S.; Ye, L.; Liu, D.; Yang, B.; Man, M.Q. Topical N-palmitoyl serinol, a commensal bacterial metabolite, prevents the de-velopment of epidermal permeability barrier dysfunction in a murine model of atopic dermatitis-like skin. Can. J. Vet. Res. 2021, 85, 201–204. [Google Scholar]
- Shin, K.-O.; Kim, S.; Park, B.; Uchida, Y.; Park, K. N-Palmitoyl Serinol Stimulates Ceramide Production through a CB1-Dependent Mechanism in In Vitro Model of Skin Inflammation. Int. J. Mol. Sci. 2021, 22, 8302. [Google Scholar] [CrossRef]
- Grill, M.; Högenauer, C.; Blesl, A.; Haybaeck, J.; Golob-Schwarzl, N.; Ferreirós, N.; Thomas, D.; Gurke, R.; Trötzmüller, M.; Köfeler, H.C.; et al. Members of the endocannabinoid system are distinctly regulated in inflammatory bowel disease and col-orectal cancer. Sci. Rep. 2019, 9, 2358. [Google Scholar] [CrossRef]
- Houser, M.; Tansey, M.G. The gut-brain axis: Is intestinal inflammation a silent driver of Parkinson’s disease pathogenesis? NPJ Parkinson’s Dis. 2017, 3, 3. [Google Scholar] [CrossRef]
- Meseguer Henarejos, A.B.; Popović, N.; Bokonjić, D.; Morales-Delgado, N.; Alonso, A.; Caballero Bleda, M.; Popović, M. Sex and Time-of-Day Impact on Anxiety and Passive Avoidance Memory Strategies in Mice. Front. Behav. Neurosci. 2020, 14, 68. [Google Scholar] [CrossRef]
- Lee, J.-S.; Kim, H.-G.; Lee, H.W.; Han, J.-M.; Lee, S.-K.; Kim, D.W.; Saravanakumar, A.; Son, C.-G. Hippocampal memory en-hancing activity of pine needle extract against scopolamine-induced amnesia in a mouse model. Sci. Rep. 2015, 5, 9651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, J.B.; Agostinho, P.; Oliveira, C. Involvement of oxidative stress in the enhancement of acetylcholinesterase activity induced by amyloid beta-peptide. Neurosci. Res. 2003, 45, 117–127. [Google Scholar] [CrossRef]
- Kaneko, N.; Kurata, M.; Yamamoto, T.; Morikawa, S.; Masumoto, J. The role of interleukin-1 in general pathology. Inflamm. Regen. 2019, 39, 12. [Google Scholar] [CrossRef] [PubMed]
- Howlett, A.C.; Blume, L.C.; Dalton, G.D. CB1 Cannabinoid Receptors and their Associated Proteins. Curr. Med. Chem. 2010, 17, 1382–1393. [Google Scholar] [CrossRef]
- Brown, A.J. Novel cannabinoid receptors. Br. J. Pharmacol. 2007, 152, 567–575. [Google Scholar] [CrossRef]
- Lan, H.; Lin, H.; Wang, C.; Wright, M.; Xu, S.; Kang, L.; Juhl, K.; Hedrick, J.; Kowalski, T. Agonists at GPR119 mediate secretion of GLP-1 from mouse enteroendocrine cells through glucose-independent pathways. Br. J. Pharmacol. 2011, 165, 2799–2807. [Google Scholar] [CrossRef]
- Overton, H.A.; Fyfe, M.C.T.; Reynet, C. GPR119, a novel G protein-coupled receptor target for the treatment of type 2 diabetes and obesity. Br. J. Pharmacol. 2008, 153 (Suppl. S1), S76–S81. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Jun, H.-S. Anti-Inflammatory Effects of GLP-1-Based Therapies beyond Glucose Control. Mediat. Inflamm. 2016, 2016, 3094642. [Google Scholar] [CrossRef]
- Borlongan, C.V.; Esparza-Salazar, F.D.J.; Lezama-Toledo, A.R.; Rivera-Monroy, G. Exendin-4 for Parkinson’s disease. Brain Circ. 2021, 7, 41–43. [Google Scholar] [CrossRef]
- Abrahan, C.E.; Miranda, G.E.; Agnolazza, D.L.; Politi, L.E.; Rotstein, N.P. Synthesis of Sphingosine Is Essential for Oxidative Stress-Induced Apoptosis of Photoreceptors. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1171–1180. [Google Scholar] [CrossRef]
- Li, P.; Li, P. Neuroprotective effect of paeoniflorin on H2O2-induced apoptosis in PC12 cells by modulation of reactive oxygen species and the inflammatory response. Exp. Ther. Med. 2015, 9, 1768–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’agostino, G.; Russo, R.; Avagliano, C.; Cristiano, C.; Meli, R.; Calignano, A. Palmitoylethanolamide Protects against the Amyloid-β25-35-Induced Learning and Memory Impairment in Mice, an Experimental Model of Alzheimer Disease. Neuropsychopharmacology 2012, 37, 1784–1792. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.Y.; Koo, B.-N.; Kim, S.Y.; Kam, E.H.; Nam, J.; Kim, E.J. Scopolamine promotes neuroinflammation and delirium-like neuropsychiatric disorder in mice. Sci. Rep. 2021, 11, 8376. [Google Scholar] [CrossRef] [PubMed]
- Reale, M.; De Angelis, F.; Di Nicola, M.; Capello, E.; Di Ioia, M.; de Luca, G.; Lugaresi, A.; Tata, A.M. Relation between pro-inflammatory cytokines and acetylcholine levels in relapsing-remitting multiple sclerosis patients. Int. J. Mol. Sci. 2012, 13, 12656–12664. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, Y.; Tak, H.; Park, D.; Song, H.; Choe, S.; Park, C.; Park, B. The Neuroprotective Effect of NEUROMIDE, a Compound Bioidentical to Commensal Bacteria Metabolites. Life 2022, 12, 1529. https://doi.org/10.3390/life12101529
Seo Y, Tak H, Park D, Song H, Choe S, Park C, Park B. The Neuroprotective Effect of NEUROMIDE, a Compound Bioidentical to Commensal Bacteria Metabolites. Life. 2022; 12(10):1529. https://doi.org/10.3390/life12101529
Chicago/Turabian StyleSeo, Yoonhee, Hyunji Tak, Dohee Park, Hyejin Song, Sooyoung Choe, Chaehyeong Park, and Byeongdeog Park. 2022. "The Neuroprotective Effect of NEUROMIDE, a Compound Bioidentical to Commensal Bacteria Metabolites" Life 12, no. 10: 1529. https://doi.org/10.3390/life12101529