
The Role of Neutrophils in Pregnancy, Term and Preterm Labour

Abstract
:1. Neutrophils and Their Effector Functions
1.1. Migration and Chemotaxis
1.2. Phagocytosis and Opsonisation
1.3. The Role of the Complement System in Enhancing Neutrophil Phagocytosis
1.4. Neutrophil Degranulation
1.5. Respiratory Burst
1.6. NETosis
1.7. The Role of Neutrophils in Sterile Inflammation
2. Neutrophils during Healthy Pregnancy
2.1. Peripheral Blood Neutrophils in Healthy Pregnancy
2.2. Neutrophils at the Maternal–Foetal Interface in Healthy Pregnancy
3. The Role of Neutrophils in Human Term Labour
3.1. Peripheral Blood Neutrophils and Term Labour
3.2. Neutrophils and the Uterus in Term Labour
3.3. Neutrophils and the Foetal Membranes in Term Labour
3.4. Neutrophils and Cervical Remodelling in Term Labour
4. The Role of Neutrophils in Preterm Labour
4.1. The Role of the Vaginal Microbiome and Ascending Infection/Inflammation
4.2. Peripheral Blood Neutrophils and Preterm Labour and PPROM
4.3. Foetal Membrane and Amniotic Fluid Neutrophils in Preterm Labour and PPROM
4.4. Neutrophils and Cervical Remodelling in Preterm Term Labour
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liew, P.X.; Kubes, P. The Neutrophil’s Role During Health and Disease. Physiol. Rev. 2019, 99, 1223–1248. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Shi, Q.; Wu, P.; Zhang, X.; Kambara, H.; Su, J.; Yu, H.; Park, S.-Y.; Guo, R.; Ren, Q.; et al. Single-Cell Transcriptome Profiling Reveals Neutrophil Heterogeneity in Homeostasis and Infection. Nat. Immunol. 2020, 21, 1119–1133. [Google Scholar] [CrossRef]
- Elghetany, M.T. Surface Antigen Changes during Normal Neutrophilic Development: A Critical Review. Blood Cells. Mol. Dis. 2002, 28, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Elghetany, M.T.; Lacombe, F. Physiologic Variations in Granulocytic Surface Antigen Expression: Impact of Age, Gender, Pregnancy, Race, and Stress. J. Leukoc. Biol. 2004, 75, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Lakschevitz, F.S.; Hassanpour, S.; Rubin, A.; Fine, N.; Sun, C.; Glogauer, M. Identification of Neutrophil Surface Marker Changes in Health and Inflammation Using High-Throughput Screening Flow Cytometry. Exp. Cell Res. 2016, 342, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Cloke, T.; Munder, M.; Taylor, G.; Müller, I.; Kropf, P. Characterization of a Novel Population of Low-Density Granulocytes Associated with Disease Severity in HIV-1 Infection. PLoS ONE 2012, 7, e48939. [Google Scholar] [CrossRef]
- Deng, Y.; Ye, J.; Luo, Q.; Huang, Z.; Peng, Y.; Xiong, G.; Guo, Y.; Jiang, H.; Li, J. Low-Density Granulocytes Are Elevated in Mycobacterial Infection and Associated with the Severity of Tuberculosis. PLoS ONE 2016, 11, e0153567. [Google Scholar] [CrossRef] [PubMed]
- Yizengaw, E.; Getahun, M.; Tajebe, F.; Cruz Cervera, E.; Adem, E.; Mesfin, G.; Hailu, A.; Van der Auwera, G.; Yardley, V.; Lemma, M.; et al. Visceral Leishmaniasis Patients Display Altered Composition and Maturity of Neutrophils as Well as Impaired Neutrophil Effector Functions. Front. Immunol. 2016, 7, 517. [Google Scholar] [CrossRef] [PubMed]
- Denny, M.F.; Yalavarthi, S.; Zhao, W.; Thacker, S.G.; Anderson, M.; Sandy, A.R.; McCune, W.J.; Kaplan, M.J. A Distinct Subset of Proinflammatory Neutrophils Isolated from Patients with Systemic Lupus Erythematosus Induces Vascular Damage and Synthesizes Type I Interferons. J. Immunol. 2010, 184, 3284–3297. [Google Scholar] [CrossRef]
- von Dadelszen, P.; Watson, R.W.G.; Noorwali, F.; Marshall, J.C.; Parodo, J.; Farine, D.; Lye, S.J.; Ritchie, J.W.K.; Rotstein, O.D. Maternal Neutrophil Apoptosis in Normal Pregnancy, Preeclampsia, and Normotensive Intrauterine Growth Restriction. Am. J. Obstet. Gynecol. 1999, 181, 408–414. [Google Scholar] [CrossRef]
- Zhang, J.; Shynlova, O.; Sabra, S.; Bang, A.; Briollais, L.; Lye, S.J. Immunophenotyping and Activation Status of Maternal Peripheral Blood Leukocytes during Pregnancy and Labour, Both Term and Preterm. J. Cell. Mol. Med. 2017, 21, 2386–2402. [Google Scholar] [CrossRef]
- Yuan, M.; Jordan, F.; McInnes, I.B.; Harnett, M.M.; Norman, J.E. Leukocytes Are Primed in Peripheral Blood for Activation during Term and Preterm Labour†. Mol. Hum. Reprod. 2009, 15, 713–724. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Kubes, P. Neutrophil Recruitment and Function in Health and Inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Metzemaekers, M.; Gouwy, M.; Proost, P. Neutrophil Chemoattractant Receptors in Health and Disease: Double-Edged Swords. Cell. Mol. Immunol. 2020, 17, 433–450. [Google Scholar] [CrossRef]
- Peiser, L. Phagocytosis: Enhancement. In eLS; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 1–12. ISBN 978-0-470-01590-2. [Google Scholar]
- Galindo-Sevilla, N.; Reyes-Arroyo, F.; Mancilla-Ramírez, J. The Role of Complement in Preterm Birth and Prematurity. J. Perinat. Med. 2019, 47, 793–803. [Google Scholar] [CrossRef]
- Futosi, K.; Fodor, S.; Mócsai, A. Neutrophil Cell Surface Receptors and Their Intracellular Signal Transduction Pathways. Int. Immunopharmacol. 2013, 17, 638–650. [Google Scholar] [CrossRef]
- Segal, A.W. How Neutrophils Kill Microbes. Annu. Rev. Immunol. 2005, 23, 197–223. [Google Scholar] [CrossRef] [PubMed]
- Faurschou, M.; Borregaard, N. Neutrophil Granules and Secretory Vesicles in Inflammation. Microbes Infect. 2003, 5, 1317–1327. [Google Scholar] [CrossRef]
- Othman, A.; Sekheri, M.; Filep, J.G. Roles of Neutrophil Granule Proteins in Orchestrating Inflammation and Immunity. FEBS J. 2022, 289, 3932–3953. [Google Scholar] [CrossRef]
- Maloney, C.G.; Kutchera, W.A.; Albertine, K.H.; McIntyre, T.M.; Prescott, S.M.; Zimmerman, G.A. Inflammatory Agonists Induce Cyclooxygenase Type 2 Expression by Human Neutrophils. J. Immunol. 1998, 160, 1402–1410. [Google Scholar]
- St-Onge, M.; Flamand, N.; Biarc, J.; Picard, S.; Bouchard, L.; Dussault, A.-A.; Laflamme, C.; James, M.J.; Caughey, G.E.; Cleland, L.G.; et al. Characterization of Prostaglandin E2 Generation through the Cyclooxygenase (COX)-2 Pathway in Human Neutrophils. Biochim. Biophys. Acta BBA-Mol. Cell Biol. Lipids 2007, 1771, 1235–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH Oxidase Activation and Bacterial Resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Chen, G.Y.; Nuñez, G. Sterile Inflammation: Sensing and Reacting to Damage. Nat. Rev. Immunol. 2010, 10, 826–837. [Google Scholar] [CrossRef]
- Eigenbrod, T.; Park, J.-H.; Harder, J.; Iwakura, Y.; Núñez, G. Cutting Edge: Critical Role for Mesothelial Cells in Necrosis-Induced Inflammation through the Recognition of IL-1α Released from Dying Cells. J. Immunol. 2008, 181, 8194–8198. [Google Scholar] [CrossRef] [PubMed]
- Verri, W.A.; Souto, F.O.; Vieira, S.M.; Almeida, S.C.L.; Fukada, S.Y.; Xu, D.; Alves-Filho, J.C.; Cunha, T.M.; Guerrero, A.T.G.; Mattos-Guimaraes, R.B.; et al. IL-33 Induces Neutrophil Migration in Rheumatoid Arthritis and Is a Target of Anti-TNF Therapy. Ann. Rheum. Dis. 2010, 69, 1697–1703. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.S.; Pulskens, W.P.; Sadler, J.J.; Butter, L.M.; Teske, G.J.; Ulland, T.K.; Eisenbarth, S.C.; Florquin, S.; Flavell, R.A.; Leemans, J.C.; et al. Necrotic Cells Trigger a Sterile Inflammatory Response through the Nlrp3 Inflammasome. Proc. Natl. Acad. Sci. USA 2009, 106, 20388–20393. [Google Scholar] [CrossRef] [PubMed]
- Pittman, K.; Kubes, P. Damage-Associated Molecular Patterns Control Neutrophil Recruitment. J. Innate Immun. 2013, 5, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Mor, G.; Aldo, P.; Alvero, A.B. The Unique Immunological and Microbial Aspects of Pregnancy. Nat. Rev. Immunol. 2017, 17, 469–482. [Google Scholar] [CrossRef]
- Sykes, L.; MacIntyre, D.A.; Yap, X.J.; Teoh, T.G.; Bennett, P.R. The Th1:Th2 Dichotomy of Pregnancy and Preterm Labour. Mediators Inflamm. 2012, 2012, 967629. [Google Scholar] [CrossRef]
- Racicot, K.; Kwon, J.-Y.; Aldo, P.; Silasi, M.; Mor, G. Understanding the Complexity of the Immune System during Pregnancy. Am. J. Reprod. Immunol. 2014, 72, 107–116. [Google Scholar] [CrossRef]
- Aghaeepour, N.; Ganio, E.A.; Mcilwain, D.; Tsai, A.S.; Tingle, M.; Van Gassen, S.; Gaudilliere, D.K.; Baca, Q.; McNeil, L.; Okada, R.; et al. An Immune Clock of Human Pregnancy. Sci. Immunol. 2017, 2, eaan2946. [Google Scholar] [CrossRef]
- Griffin, J.F.T.; Beck, I. A Longitudinal Study of Leucocyte Numbers and Mitogenesis during the Last Ten Weeks of Human Pregnancy. J. Reprod. Immunol. 1983, 5, 239–247. [Google Scholar] [CrossRef]
- Luppi, P.; Haluszczak, C.; Betters, D.; Richard, C.A.H.; Trucco, M.; DeLoia, J.A. Monocytes Are Progressively Activated in the Circulation of Pregnant Women. J. Leukoc. Biol. 2002, 72, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Tripathi, A.K.; Mishra, S.; Amzarul, M.; Vaish, A.K. Physiological Changes in Hematological Parameters During Pregnancy. Indian J. Hematol. Blood Transfus. 2012, 28, 144–146. [Google Scholar] [CrossRef] [PubMed]
- Lampé, R.; Kövér, Á.; Szűcs, S.; Pál, L.; Árnyas, E.; Ádány, R.; Póka, R. Phagocytic Index of Neutrophil Granulocytes and Monocytes in Healthy and Preeclamptic Pregnancy. J. Reprod. Immunol. 2015, 107, 26–30. [Google Scholar] [CrossRef]
- Pramanik, S.S.; Pramanik, T.; Mondal, S.C.; Chanda, R. Number, Maturity and Phagocytic Activity of Neutrophils in the Three Trimesters of Pregnancy. East. Mediterr. Health J. 2007, 13, 862–864. [Google Scholar]
- Sacks, G.P.; Studena, K.; Sargent, I.L.; Redman, C.W.G. Normal Pregnancy and Preeclampsia Both Produce Inflammatory Changes in Peripheral Blood Leukocytes Akin to Those of Sepsis. Am. J. Obstet. Gynecol. 1998, 179, 80–86. [Google Scholar] [CrossRef]
- Naccasha, N.; Gervasi, M.-T.; Chaiworapongsa, T.; Berman, S.; Yoon, B.H.; Maymon, E.; Romero, R. Phenotypic and Metabolic Characteristics of Monocytes and Granulocytes in Normal Pregnancy and Maternal Infection. Am. J. Obstet. Gynecol. 2001, 185, 1118–1123. [Google Scholar] [CrossRef]
- Köstlin, N.; Kugel, H.; Spring, B.; Leiber, A.; Marmé, A.; Henes, M.; Rieber, N.; Hartl, D.; Poets, C.F.; Gille, C. Granulocytic Myeloid Derived Suppressor Cells Expand in Human Pregnancy and Modulate T-Cell Responses. Eur. J. Immunol. 2014, 44, 2582–2591. [Google Scholar] [CrossRef]
- Farias-Jofre, M.; Romero, R.; Galaz, J.; Xu, Y.; Tao, L.; Demery-Poulos, C.; Arenas-Hernandez, M.; Bhatti, G.; Liu, Z.; Kawahara, N.; et al. Pregnancy Tailors Endotoxin-Induced Monocyte and Neutrophil Responses in the Maternal Circulation. Inflamm. Res. 2022, 71, 653–668. [Google Scholar] [CrossRef] [PubMed]
- Kropf, P.; Baud, D.; Marshall, S.E.; Munder, M.; Mosley, A.; Fuentes, J.M.; Bangham, C.R.M.; Taylor, G.P.; Herath, S.; Choi, B.-S.; et al. Arginase Activity Mediates Reversible T Cell Hyporesponsiveness in Human Pregnancy. Eur. J. Immunol. 2007, 37, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Alqurashi, M.; Thurairajah, S.; Adan, M.; Chan, D.; Teoh, T.G.; Bennett, P.R.; Muller, I.; Kropf, P.; Sykes, L. Characterisation of Normal and Low Density Granulocytes in Pregnancy, Term Labour, and Women at High Risk of Preterm Labour. Reprod. Sci. 2019, 26, 122A–123A. [Google Scholar]
- Ssemaganda, A.; Kindinger, L.; Bergin, P.; Nielsen, L.; Mpendo, J.; Ssetaala, A.; Kiwanuka, N.; Munder, M.; Teoh, T.G.; Kropf, P.; et al. Characterization of Neutrophil Subsets in Healthy Human Pregnancies. PLoS ONE 2014, 9, e85696. [Google Scholar] [CrossRef]
- Roehrl, M.H.A.; Wang, J.Y. Immature Granulocytes in Pregnancy: A Story of Virchow, Anxious Fathers, and Expectant Mothers. Am. J. Hematol. 2011, 86, 307–308. [Google Scholar] [CrossRef]
- Luppi, P. How Immune Mechanisms Are Affected by Pregnancy. Vaccine 2003, 21, 3352–3357. [Google Scholar] [CrossRef]
- Thornton, C.A. Immunology of Pregnancy. Proc. Nutr. Soc. 2010, 69, 357–365. [Google Scholar] [CrossRef]
- Bert, S.; Ward, E.J.; Nadkarni, S. Neutrophils in Pregnancy: New Insights into Innate and Adaptive Immune Regulation. Immunology 2021, 164, 665–676. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; Romero, R.; Xu, Y.; Miller, D.; Leng, Y.; Panaitescu, B.; Silva, P.; Faro, J.; Alhousseini, A.; Gill, N.; et al. The Immunophenotype of Amniotic Fluid Leukocytes in Normal and Complicated Pregnancies. Am. J. Reprod. Immunol. 2018, 79, e12827. [Google Scholar] [CrossRef]
- Thomson, A.J.; Telfer, J.F.; Young, A.; Campbell, S.; Stewart, C.J.; Cameron, I.T.; Greer, I.A.; Norman, J.E. Leukocytes Infiltrate the Myometrium during Human Parturition: Further Evidence That Labour Is an Inflammatory Process. Hum. Reprod. Oxf. Engl. 1999, 14, 229–236. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Moran, P.; Bulmer, J.N.; Searle, R.F.; Robson, S.C. Macrophages and Not Granulocytes Are Involved in Cervical Ripening. J. Reprod. Immunol. 2005, 66, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Word, R.A.; Li, X.-H.; Hnat, M.; Carrick, K. Dynamics of Cervical Remodeling during Pregnancy and Parturition: Mechanisms and Current Concepts. Semin. Reprod. Med. 2007, 25, 069–079. [Google Scholar] [CrossRef] [PubMed]
- Myers, D.A. The Recruitment and Activation of Leukocytes into the Immune Cervix: Further Support That Cervical Remodeling Involves an Immune and Inflammatory Mechanism. Biol. Reprod. 2012, 87, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golightly, E.; Jabbour, H.N.; Norman, J.E. Endocrine Immune Interactions in Human Parturition. Mol. Cell. Endocrinol. 2011, 335, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.B.; Buhimschi, C.S.; Norwitz, E.R. Normal Labor: Mechanism and Duration. Obstet. Gynecol. Clin. N. Am. 2005, 32, 145–164. [Google Scholar] [CrossRef]
- Terzidou, V. Biochemical and Endocrinological Preparation for Parturition. Best Pract. Res. Clin. Obstet. Gynaecol. 2007, 21, 729–756. [Google Scholar] [CrossRef]
- Sennström, M.B.; Ekman, G.; Westergren-Thorsson, G.; Malmström, A.; Byström, B.; Endrésen, U.; Mlambo, N.; Norman, M.; Ståbi, B.; Brauner, A. Human Cervical Ripening, an Inflammatory Process Mediated by Cytokines. Mol. Hum. Reprod. 2000, 6, 375–381. [Google Scholar] [CrossRef]
- Challis, J.R.; Lockwood, C.J.; Myatt, L.; Norman, J.E.; Strauss, J.F.; Petraglia, F. Inflammation and Pregnancy. Reprod. Sci. 2009, 16, 206–215. [Google Scholar] [CrossRef]
- Wright, H.L.; Moots, R.J.; Bucknall, R.C.; Edwards, S.W. Neutrophil Function in Inflammation and Inflammatory Diseases. Rheumatology 2010, 49, 1618–1631. [Google Scholar] [CrossRef]
- Osman, I.; Young, A.; Ledingham, M.A.; Thomson, A.J.; Jordan, F.; Greer, I.A.; Norman, J.E. Leukocyte Density and Pro-inflammatory Cytokine Expression in Human Fetal Membranes, Decidua, Cervix and Myometrium before and during Labour at Term. Mol. Hum. Reprod. 2003, 9, 41–45. [Google Scholar] [CrossRef]
- Young, A.; Thomson, A.J.; Ledingham, M.; Jordan, F.; Greer, I.A.; Norman, J.E. Immunolocalization of Proinflammatory Cytokines in Myometrium, Cervix, and Fetal Membranes During Human Parturition at Term1. Biol. Reprod. 2002, 66, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, Y.; Moran, P.; Searle, R.F.; Bulmer, J.N.; Robson, S.C. Interleukin-8 Is Involved in Cervical Dilatation but Not in Prelabour Cervical Ripening. Clin. Exp. Immunol. 2004, 138, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Delgado, I.; Neubert, R.; Dudenhausen, J.W. Changes in White Blood Cells during Parturition in Mothers and Newborn. Gynecol. Obstet. Investig 1994, 38, 227–235. [Google Scholar] [CrossRef]
- Bollopragada, S.; Youssef, R.; Jordan, F.; Greer, I.; Norman, J.; Nelson, S. Term Labor Is Associated with a Core Inflammatory Response in Human Fetal Membranes, Myometrium, and Cervix. Am. J. Obstet. Gynecol. 2009, 200, 104.e1–104.e11. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.; Fischer, D.-C.; Ruck, P.; Marx, T.; Kaiserling, E.; Oberpichler, A.; Tschesche, H.; Rath, W. Parturition at Term: Parallel Increases in Interleukin-8 and Proteinase Concentrations and Neutrophil Count in the Lower Uterine Segment. Hum. Reprod. 1999, 14, 1096–1100. [Google Scholar] [CrossRef]
- Osman, I.; Young, A.; Jordan, F.; Greer, I.A.; Norman, J.E. Leukocyte Density and Proinflammatory Mediator Expression in Regional Human Fetal Membranes and Decidua Before and During Labot at Term. J. Soc. Gynecol. Investig. 2006, 13, 97–103. [Google Scholar] [CrossRef]
- Takahashi, N.; Okuno, T.; Fujii, H.; Makino, S.; Takahashi, M.; Ohba, M.; Saeki, K.; Itakura, A.; Takeda, S.; Yokomizo, T. Up-Regulation of Cytosolic Prostaglandin E Synthase in Fetal-Membrane and Amniotic Prostaglandin E2 Accumulation in Labor. PLoS ONE 2021, 16, e0250638. [Google Scholar] [CrossRef]
- Norman, J.E. Cervical Function and Prematurity. Best Pract. Res. Clin. Obstet. Gynaecol. 2007, 21, 791–806. [Google Scholar] [CrossRef]
- Bokström, H.; Brännström, M.; Alexandersson, M.; Norström, A. Leukocyte Subpopulations in the Human Uterine Cervical Stroma at Early and Term Pregnancy. Hum. Reprod. 1997, 12, 586–590. [Google Scholar] [CrossRef]
- Hein, M.; Petersen, A.C.; Helmig, R.B.; Uldbjerg, N.; Reinholdt, J. Immunoglobulin Levels and Phagocytes in the Cervical Mucus Plug at Term of Pregnancy. Acta Obstet. Gynecol. Scand. 2005, 84, 734–742. [Google Scholar] [CrossRef]
- Timmons, B.C.; Mahendroo, M.S. Timing of Neutrophil Activation and Expression of Proinflammatory Markers Do Not Support a Role for Neutrophils in Cervical Ripening in the Mouse1. Biol. Reprod. 2006, 74, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Sennström, M.B.; Brauner, A.; Byström, B.; Malmström, A.; Ekman, G. Matrix Metalloproteinase-8 Correlates with the Cervical Ripening Process in Humans. Acta Obstet. Gynecol. Scand. 2003, 82, 904–911. [Google Scholar] [CrossRef] [PubMed]
- Stygar, D.; Wang, H.; Vladic, Y.S.; Ekman, G.; Eriksson, H.; Sahlin, L. Increased Level of Matrix Metalloproteinases 2 and 9 in the Ripening Process of the Human Cervix1. Biol. Reprod. 2002, 67, 889–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osmers, R.; Rath, W.; Adelmann-Grill, B.C.; Fittkow, C.; Kuloczik, M.; Szeverényi, M.; Tschesche, H.; Kuhn, W. Origin of Cervical Collagenase during Parturition. Am. J. Obstet. Gynecol. 1992, 166, 1455–1460. [Google Scholar] [CrossRef]
- Blencowe, H.; Cousens, S.; Oestergaard, M.Z.; Chou, D.; Moller, A.-B.; Narwal, R.; Adler, A.; Vera Garcia, C.; Rohde, S.; Say, L.; et al. National, Regional, and Worldwide Estimates of Preterm Birth Rates in the Year 2010 with Time Trends since 1990 for Selected Countries: A Systematic Analysis and Implications. Lancet 2012, 379, 2162–2172. [Google Scholar] [CrossRef]
- Liu, L.; Oza, S.; Hogan, D.; Perin, J.; Rudan, I.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, Regional, and National Causes of Child Mortality in 2000–13, with Projections to Inform Post-2015 Priorities: An Updated Systematic Analysis. Lancet 2015, 385, 430–440. [Google Scholar] [CrossRef]
- Agrawal, V.; Hirsch, E. Intrauterine Infection and Preterm Labor. Semin. Fetal. Neonatal Med. 2012, 17, 12–19. [Google Scholar] [CrossRef]
- Kim, C.J.; Romero, R.; Chaemsaithong, P.; Chaiyasit, N.; Yoon, B.H.; Kim, Y.M. Acute Chorioamnionitis and Funisitis: Definition, Pathologic Features, and Clinical Significance. Am. J. Obstet. Gynecol. 2015, 213, S29–S52. [Google Scholar] [CrossRef]
- Steel, J.H.; O’Donoghue, K.; Kennea, N.L.; Sullivan, M.H.F.; Edwards, A.D. Maternal Origin of Inflammatory Leukocytes in Preterm Fetal Membranes, Shown by Fluorescence in Situ Hybridisation. Placenta 2005, 26, 672–677. [Google Scholar] [CrossRef]
- Yamada, T.; Minakami, H.; Matsubara, S.; Yatsuda, T.; Sato, I. Changes in Polymorphonuclear Leukocytes in the Vagina of Patients with Preterm Labor. Gynecol. Obstet. Investig 1998, 45, 32–34. [Google Scholar] [CrossRef]
- Hezelgrave, N.L.; Seed, P.T.; Chin-Smith, E.C.; Ridout, A.E.; Shennan, A.H.; Tribe, R.M. Cervicovaginal Natural Antimicrobial Expression in Pregnancy and Association with Spontaneous Preterm Birth. Sci. Rep. 2020, 10, 12018. [Google Scholar] [CrossRef] [PubMed]
- Whitworth, M.K.; Pafilis, I.; Vince, G.; Quenby, S. Cervical Leukocyte Sub-Populations in Idiopathic Preterm Labour. J. Reprod. Immunol. 2007, 75, 48–55. [Google Scholar] [CrossRef]
- Stranik, J.; Kacerovsky, M.; Andrys, C.; Soucek, O.; Bolehovska, R.; Holeckova, M.; Matulova, J.; Jacobsson, B.; Musilova, I. Intra-Amniotic Infection and Sterile Intra-Amniotic Inflammation Are Associated with Elevated Concentrations of Cervical Fluid Interleukin-6 in Women with Spontaneous Preterm Labor with Intact Membranes. J. Matern. Fetal Neonatal Med. 2021, 1–9. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; Romero, R.; Varrey, A.; Leng, Y.; Miller, D.; Done, B.; Xu, Y.; Bhatti, G.; Motomura, K.; Gershater, M.; et al. RNA Sequencing Reveals Diverse Functions of Amniotic Fluid Neutrophils and Monocytes/Macrophages in Intra-Amniotic Infection. J. Innate Immun. 2021, 13, 63–82. [Google Scholar] [CrossRef]
- Nadeau-Vallée, M.; Obari, D.; Palacios, J.; Brien, M.-È.; Duval, C.; Chemtob, S.; Girard, S. Sterile Inflammation and Pregnancy Complications: A Review. Reproduction 2016, 152, R277–R292. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Miranda, J.; Chaiworapongsa, T.; Korzeniewski, S.J.; Chaemsaithong, P.; Gotsch, F.; Dong, Z.; Ahmed, A.I.; Yoon, B.H.; Hassan, S.S.; et al. Prevalence and Clinical Significance of Sterile Intra-Amniotic Inflammation in Patients with Preterm Labor and Intact Membranes. Am. J. Reprod. Immunol. 2014, 72, 458–474. [Google Scholar] [CrossRef]
- Baumbusch, M.A.; Buhimschi, C.S.; Oliver, E.A.; Zhao, G.; Thung, S.; Rood, K.; Buhimschi, I.A. High Mobility Group-Box 1 (HMGB1) Levels Are Increased in Amniotic Fluid of Women with Intra-Amniotic Inflammation-Determined Preterm Birth, and the Source May Be the Damaged Fetal Membranes. Cytokine 2016, 81, 82–87. [Google Scholar] [CrossRef]
- Figueroa, R.; Garry, D.; Elimian, A.; Patel, K.; Sehgal, P.B.; Tejani, N. Evaluation of Amniotic Fluid Cytokines in Preterm Labor and Intact Membranes. J. Matern. Fetal Neonatal Med. 2005, 18, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Brody, D.T.; Oyarzun, E.; Mazor, M.; King Wu, Y.; Hobbins, J.C.; Durum, S.K. Infection and Labor: III. Interleukin-1: A Signal for the Onset of Parturition. Am. J. Obstet. Gynecol. 1989, 160, 1117–1123. [Google Scholar] [CrossRef]
- Thaxton, J.E.; Romero, R.; Sharma, S. TLR9 Activation Coupled to IL-10 Deficiency Induces Adverse Pregnancy Outcomes. J. Immunol. 2009, 183, 1144–1154. [Google Scholar] [CrossRef]
- Scharfe-Nugent, A.; Corr, S.C.; Carpenter, S.B.; Keogh, L.; Doyle, B.; Martin, C.; Fitzgerald, K.A.; Daly, S.; O’Leary, J.J.; O’Neill, L.A.J. TLR9 Provokes Inflammation in Response to Fetal DNA: Mechanism for Fetal Loss in Preterm Birth and Preeclampsia. J. Immunol. 2012, 188, 5706–5712. [Google Scholar] [CrossRef]
- Aagaard, K.; Riehle, K.; Ma, J.; Segata, N.; Mistretta, T.-A.; Coarfa, C.; Raza, S.; Rosenbaum, S.; den Veyver, I.V.; Milosavljevic, A.; et al. A Metagenomic Approach to Characterization of the Vaginal Microbiome Signature in Pregnancy. PLoS ONE 2012, 7, e36466. [Google Scholar] [CrossRef]
- DiGiulio, D.B.; Callahan, B.J.; McMurdie, P.J.; Costello, E.K.; Lyell, D.J.; Robaczewska, A.; Sun, C.L.; Goltsman, D.S.A.; Wong, R.J.; Shaw, G.; et al. Temporal and Spatial Variation of the Human Microbiota during Pregnancy. Proc. Natl. Acad. Sci. USA 2015, 112, 11060–11065. [Google Scholar] [CrossRef] [Green Version]
- Farage, M.; Maibach, H. Lifetime Changes in the Vulva and Vagina. Arch. Gynecol. Obstet. 2006, 273, 195–202. [Google Scholar] [CrossRef]
- Bayar, E.; Bennett, P.R.; Chan, D.; Sykes, L.; MacIntyre, D.A. The Pregnancy Microbiome and Preterm Birth. Semin. Immunopathol. 2020, 42, 487–499. [Google Scholar] [CrossRef]
- Grewal, K.; MacIntyre, D.A.; Bennett, P.R. The Reproductive Tract Microbiota in Pregnancy. Biosci. Rep. 2021, 41, BSR20203908. [Google Scholar] [CrossRef]
- Jayaprakash, T.P.; Wagner, E.C.; van Schalkwyk, J.; Albert, A.Y.K.; Hill, J.E.; Money, D.M.; Group, P.S. High Diversity and Variability in the Vaginal Microbiome in Women Following Preterm Premature Rupture of Membranes (PPROM): A Prospective Cohort Study. PLoS ONE 2016, 11, e0166794. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, E.A.; Walther-Antonio, M.; MacLean, A.M.; Gohl, D.M.; Beckman, K.B.; Chen, J.; White, B.; Creedon, D.J.; Chia, N. Persistent Microbial Dysbiosis in Preterm Premature Rupture of Membranes from Onset until Delivery. PeerJ 2015, 3, e1398. [Google Scholar] [CrossRef]
- Kacerovsky, M.; Vrbacky, F.; Kutova, R.; Pliskova, L.; Andrys, C.; Musilova, I.; Menon, R.; Lamont, R.; Nekvindova, J. Cervical Microbiota in Women with Preterm Prelabor Rupture of Membranes. PLoS ONE 2015, 10, e0126884. [Google Scholar] [CrossRef]
- Brown, R.G.; Marchesi, J.R.; Lee, Y.S.; Smith, A.; Lehne, B.; Kindinger, L.M.; Terzidou, V.; Holmes, E.; Nicholson, J.K.; Bennett, P.R.; et al. Vaginal Dysbiosis Increases Risk of Preterm Fetal Membrane Rupture, Neonatal Sepsis and Is Exacerbated by Erythromycin. BMC Med. 2018, 16, 9. [Google Scholar] [CrossRef]
- Kindinger, L.M.; MacIntyre, D.A.; Lee, Y.S.; Marchesi, J.R.; Smith, A.; McDonald, J.A.K.; Terzidou, V.; Cook, J.R.; Lees, C.; Israfil-Bayli, F.; et al. Relationship between Vaginal Microbial Dysbiosis, Inflammation, and Pregnancy Outcomes in Cervical Cerclage. Sci. Transl. Med. 2016, 8, 350ra102. [Google Scholar] [CrossRef] [PubMed]
- Fettweis, J.M.; Serrano, M.G.; Brooks, J.P.; Edwards, D.J.; Girerd, P.H.; Parikh, H.I.; Huang, B.; Arodz, T.J.; Edupuganti, L.; Glascock, A.L.; et al. The Vaginal Microbiome and Preterm Birth. Nat. Med. 2019, 25, 1012–1021. [Google Scholar] [CrossRef]
- Chan, D.; Bennett, P.R.; Lee, Y.S.; Kundu, S.; Teoh, T.G.; Adan, M.; Ahmed, S.; Brown, R.G.; David, A.L.; Lewis, H.V.; et al. Microbial-Driven Preterm Labour Involves Crosstalk between the Innate and Adaptive Immune Response. Nat. Commun. 2022, 13, 975. [Google Scholar] [CrossRef] [PubMed]
- Mohd Zaki, A.; Hadingham, A.; Flaviani, F.; Haque, Y.; Mi, J.D.; Finucane, D.; Dalla Valle, G.; Mason, A.J.; Saqi, M.; Gibbons, D.L.; et al. Neutrophils Dominate the Cervical Immune Cell Population in Pregnancy and Their Transcriptome Correlates with the Microbial Vaginal Environment. Front. Microbiol. 2022, 13, 904451. [Google Scholar] [CrossRef]
- Hunter, P.J.; Sheikh, S.; David, A.L.; Peebles, D.M.; Klein, N. Cervical Leukocytes and Spontaneous Preterm Birth. J. Reprod. Immunol. 2016, 113, 42–49. [Google Scholar] [CrossRef]
- Molina, B.; Bayar, E.; Lee, Y.S.; Muller, I.; Botto, M.; MacIntyre, D.; Bennett, P.R.; Kropf, P.; Sykes, L. Cervicovaginal Inflammation and Neutrophils Infiltration/Activation in Women at High-Risk of Prematurity. BJOG Int. J. Obstet. Gynaecol. 2022, 129, 47–62. [Google Scholar] [CrossRef]
- Clarke, T.B.; Davis, K.M.; Lysenko, E.S.; Zhou, A.Y.; Yu, Y.; Weiser, J.N. Recognition of Peptidoglycan from the Microbiota by Nod1 Enhances Systemic Innate Immunity. Nat. Med. 2010, 16, 228–231. [Google Scholar] [CrossRef]
- Ozel, A.; Alici Davutoglu, E.; Yurtkal, A.; Madazli, R. How Do Platelet-to-Lymphocyte Ratio and Neutrophil-to-Lymphocyte Ratio Change in Women with Preterm Premature Rupture of Membranes, and Threaten Preterm Labour? J. Obstet. Gynaecol. 2020, 40, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, M.P.A.S.; Sravani, V.L. Role of Neutrophil-Lymphocyte Ratio in Determining the Outcomes of Preterm Premature Rupture of Membranes. Int. J. Reprod. Contracept. Obstet. Gynecol. 2021, 10, 1617–1620. [Google Scholar] [CrossRef]
- Vakili, S.; Torabinavid, P.; Tabrizi, R.; Shojazadeh, A.; Asadi, N.; Hessami, K. The Association of Inflammatory Biomarker of Neutrophil-to-Lymphocyte Ratio with Spontaneous Preterm Delivery: A Systematic Review and Meta-Analysis. Mediators Inflamm. 2021, 2021, 6668381. [Google Scholar] [CrossRef]
- Gervasi, M.-T.; Chaiworapongsa, T.; Naccasha, N.; Blackwell, S.; Yoon, B.H.; Maymon, E.; Romero, R. Phenotypic and Metabolic Characteristics of Maternal Monocytes and Granulocytes in Preterm Labor with Intact Membranes. Am. J. Obstet. Gynecol. 2001, 185, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Takeda, J.; Fang, X.; Olson, D.M. Pregnant Human Peripheral Leukocyte Migration during Several Late Pregnancy Clinical Conditions: A Cross-Sectional Observational Study. BMC Pregnancy Childbirth 2017, 17, 16. [Google Scholar] [CrossRef] [PubMed]
- Andrews, W.W.; Goldenberg, R.L.; Faye-Petersen, O.; Cliver, S.; Goepfert, A.R.; Hauth, J.C. The Alabama Preterm Birth Study: Polymorphonuclear and Mononuclear Cell Placental Infiltrations, Other Markers of Inflammation, and Outcomes in 23- to 32-Week Preterm Newborn Infants. Am. J. Obstet. Gynecol. 2006, 195, 803–808. [Google Scholar] [CrossRef]
- Tita, A.T.N.; Andrews, W.W. Diagnosis and Management of Clinical Chorioamnionitis. Clin. Perinatol. 2010, 37, 339–354. [Google Scholar] [CrossRef]
- McNamara, M.F.; Wallis, T.; Qureshi, F.; Jacques, S.M.; Gonik, B. Determining the Maternal and Fetal Cellular Immunologic Contributions in Preterm Deliveries with Clinical or Subclinical Chorioamnionitis. Infect. Dis. Obstet. Gynecol. 1900, 5, 463137. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; Romero, R.; Xu, Y.; Leng, Y.; Garcia-Flores, V.; Miller, D.; Jacques, S.M.; Hassan, S.S.; Faro, J.; Alsamsam, A.; et al. Are Amniotic Fluid Neutrophils in Women with Intraamniotic Infection and/or Inflammation of Fetal or Maternal Origin? Am. J. Obstet. Gynecol. 2017, 217, 693.e1–693.e16. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.; Potter, J.A.; Mor, G.; Abrahams, V.M. Lipopolysaccharide-Stimulated Human Fetal Membranes Induce Neutrophil Activation and Release of Vital Neutrophil Extracellular Traps. J. Immunol. 2019, 203, 500–510. [Google Scholar] [CrossRef]
- Lappas, M.; Woodruff, T.M.; Taylor, S.M.; Permezel, M. Complement C5a Regulates Prolabor Mediators in Human Placenta1. Biol. Reprod. 2012, 86, 190. [Google Scholar] [CrossRef]
- Segura-Cervantes, E.; Mancilla-Ramirez, J.; Zurita, L.; Paredes, Y.; Arredondo, J.L.; Galindo-Sevilla, N. Blood SC5b-9 Complement Levels Increase at Parturition during Term and Preterm Labor. J. Reprod. Immunol. 2015, 109, 24–30. [Google Scholar] [CrossRef]
- Hillier, S.L.; Witkin, S.S.; Krohn, M.A.; Watts, D.H.; Kiviat, N.B.; Eschenbach, D.A. The Relationship of Amniotic Fluid Cytokines and Preterm Delivery, Amniotic Fluid Infection, Histologic Chorioamnionitis, and Chorioamnion Infection. Obstet. Gynecol. 1993, 81, 941–948. [Google Scholar]
- Soto, E.; Romero, R.; Richani, K.; Yoon, B.H.; Chaiworapongsa, T.; Vaisbuch, E.; Mittal, P.; Erez, O.; Gotsch, F.; Mazor, M.; et al. Evidence for Complement Activation in the Amniotic Fluid of Women with Spontaneous Preterm Labor and Intra-Amniotic Infection. J. Matern. Fetal Neonatal Med. 2009, 22, 983–992. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Park, K.H.; Park, H.; Yoo, H.-N.; Kook, S.Y.; Jeon, S.J. Complement C3a, But Not C5a, Levels in Amniotic Fluid Are Associated with Intra-Amniotic Infection and/or Inflammation and Preterm Delivery in Women with Cervical Insufficiency or an Asymptomatic Short Cervix (≤25 Mm). J. Korean Med. Sci. 2018, 33, e220. [Google Scholar] [CrossRef] [PubMed]
- Maymon, E.; Romero, R.; Pacora, P.; Gomez, R.; Athayde, N.; Edwin, S.; Yoon, B.H. Human Neutrophil Collagenase (Matrix Metalloproteinase 8) in Parturition, Premature Rupture of the Membranes, and Intrauterine Infection. Am. J. Obstet. Gynecol. 2000, 183, 94–99. [Google Scholar] [CrossRef]
- Helmig, B.R.; Romero, R.; Espinoza, J.; Chaiworapongsa, T.; Bujold, E.; Gomez, R.; Ohlsson, K.; Uldbjerg, N. Neutrophil Elastase and Secretory Leukocyte Protease Inhibitor in Prelabor Rupture of Membranes, Parturition and Intra-Amniotic Infection. J. Matern. Fetal Neonatal Med. 2002, 12, 237–246. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; Romero, R.; Garcia-Flores, V.; Xu, Y.; Leng, Y.; Alhousseini, A.; Hassan, S.S.; Panaitescu, B. Amniotic Fluid Neutrophils Can Phagocytize Bacteria: A Mechanism for Microbial Killing in the Amniotic Cavity. Am. J. Reprod. Immunol. 2017, 78, e12723. [Google Scholar] [CrossRef]
- Galaz, J.; Romero, R.; Xu, Y.; Miller, D.; Slutsky, R.; Levenson, D.; Hsu, C.-D.; Gomez-Lopez, N. Cellular Immune Responses in Amniotic Fluid of Women with Preterm Clinical Chorioamnionitis. Inflamm. Res. 2020, 69, 203–216. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; Romero, R.; Xu, Y.; Miller, D.; Unkel, R.; Shaman, M.; Jacques, S.M.; Panaitescu, B.; Garcia-Flores, V.; Hassan, S.S. Neutrophil Extracellular Traps in the Amniotic Cavity of Women with Intra-Amniotic Infection: A New Mechanism of Host Defense. Reprod. Sci. Thousand Oaks Calif 2017, 24, 1139–1153. [Google Scholar] [CrossRef]
- Kim, C.J.; Yoon, B.H.; Romero, R.; Moon, J.B.; Kim, M.; Park, S.-S.; Chi, J.G. Umbilical Arteritis and Phlebitis Mark Different Stages of the Fetal Inflammatory Response. Am. J. Obstet. Gynecol. 2001, 185, 496–500. [Google Scholar] [CrossRef]
- Heng, Y.J.; Liong, S.; Permezel, M.; Rice, G.E.; Di Quinzio, M.K.W.; Georgiou, H.M. Human Cervicovaginal Fluid Biomarkers to Predict Term and Preterm Labor. Front. Physiol. 2015, 6, 151. [Google Scholar] [CrossRef]
- Kindinger, L.M.; Bennett, P.R.; Lee, Y.S.; Marchesi, J.R.; Smith, A.; Cacciatore, S.; Holmes, E.; Nicholson, J.K.; Teoh, T.G.; MacIntyre, D.A. The Interaction between Vaginal Microbiota, Cervical Length, and Vaginal Progesterone Treatment for Preterm Birth Risk. Microbiome 2017, 5, 6. [Google Scholar] [CrossRef]
- Tabatabaei, N.; Eren, A.; Barreiro, L.; Yotova, V.; Dumaine, A.; Allard, C.; Fraser, W. Vaginal Microbiome in Early Pregnancy and Subsequent Risk of Spontaneous Preterm Birth: A Case–Control Study. BJOG Int. J. Obstet. Gynaecol. 2019, 126, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.C.; Bocking, A.; Hill, J.E.; Money, D.M.; Money, D.; Bocking, A.; Hemmingsen, S.; Hill, J.; Reid, G.; Dumonceaux, T.; et al. Increased Richness and Diversity of the Vaginal Microbiota and Spontaneous Preterm Birth. Microbiome 2018, 6, 117. [Google Scholar] [CrossRef] [PubMed]
- Florova, V.; Romero, R.; Tarca, A.L.; Galaz, J.; Motomura, K.; Ahmad, M.M.; Hsu, C.-D.; Hsu, R.; Tong, A.; Ravel, J.; et al. Vaginal Host Immune-Microbiome Interactions in a Cohort of Primarily African-American Women Who Ultimately Underwent Spontaneous Preterm Birth or Delivered at Term. Cytokine 2021, 137, 155316. [Google Scholar] [CrossRef] [PubMed]
- Witkin, S.S.; Moron, A.F.; Ridenhour, B.J.; Minis, E.; Hatanaka, A.; Sarmento, S.G.P.; Franca, M.S.; Carvalho, F.H.C.; Hamamoto, T.K.; Mattar, R.; et al. Vaginal Biomarkers That Predict Cervical Length and Dominant Bacteria in the Vaginal Microbiomes of Pregnant Women. mBio 2019, 10, e02242-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.; Jung, Y.M.; Lee, Y.E.; Park, Y.; Kim, Y.M.; Park, K.H. Complement and Other Immune-Related Factors in Cervicovaginal Fluid Associated with Intra-Amniotic Infection/Inflammation and Spontaneous Preterm Delivery in Women with Preterm Labor. Arch. Gynecol. Obstet. 2020, 301, 1431–1439. [Google Scholar] [CrossRef] [PubMed]
- Elovitz, M.A.; Gajer, P.; Riis, V.; Brown, A.G.; Humphrys, M.S.; Holm, J.B.; Ravel, J. Cervicovaginal Microbiota and Local Immune Response Modulate the Risk of Spontaneous Preterm Delivery. Nat. Commun. 2019, 10, 1305. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, P.C.; de Carvalho, J.B.J.; do Amaral, R.L.G.; da Silveira Gonçalves, A.K.; Eleutério, J., Jr.; Guimarães, F. Identification of Immune Cells by Flow Cytometry in Vaginal Lavages from Women with Vulvovaginitis and Normal Microflora. Am. J. Reprod. Immunol. 2012, 67, 198–205. [Google Scholar] [CrossRef]
- Beghini, J.; Giraldo, P.C.; Riboldi, R.; Amaral, R.L.G.; Eleutério, J.; Witkin, S.S.; Guimarães, F. Altered CD16 Expression on Vaginal Neutrophils from Women with Vaginitis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 167, 96–99. [Google Scholar] [CrossRef]
- Luo, L.; Ibaragi, T.; Maeda, M.; Nozawa, M.; Kasahara, T.; Sakai, M.; Sasaki, Y.; Tanebe, K.; Saito, S. Interleukin-8 Levels and Granulocyte Counts in Cervical Mucus During Pregnancy. Am. J. Reprod. Immunol. 2000, 43, 78–84. [Google Scholar] [CrossRef]
- Simhan, H.N.; Caritis, S.N.; Krohn, M.A.; Hillier, S.L. Elevated Vaginal PH and Neutrophils Are Associated Strongly with Early Spontaneous Preterm Birth. Am. J. Obstet. Gynecol. 2003, 189, 1150–1154. [Google Scholar] [CrossRef]




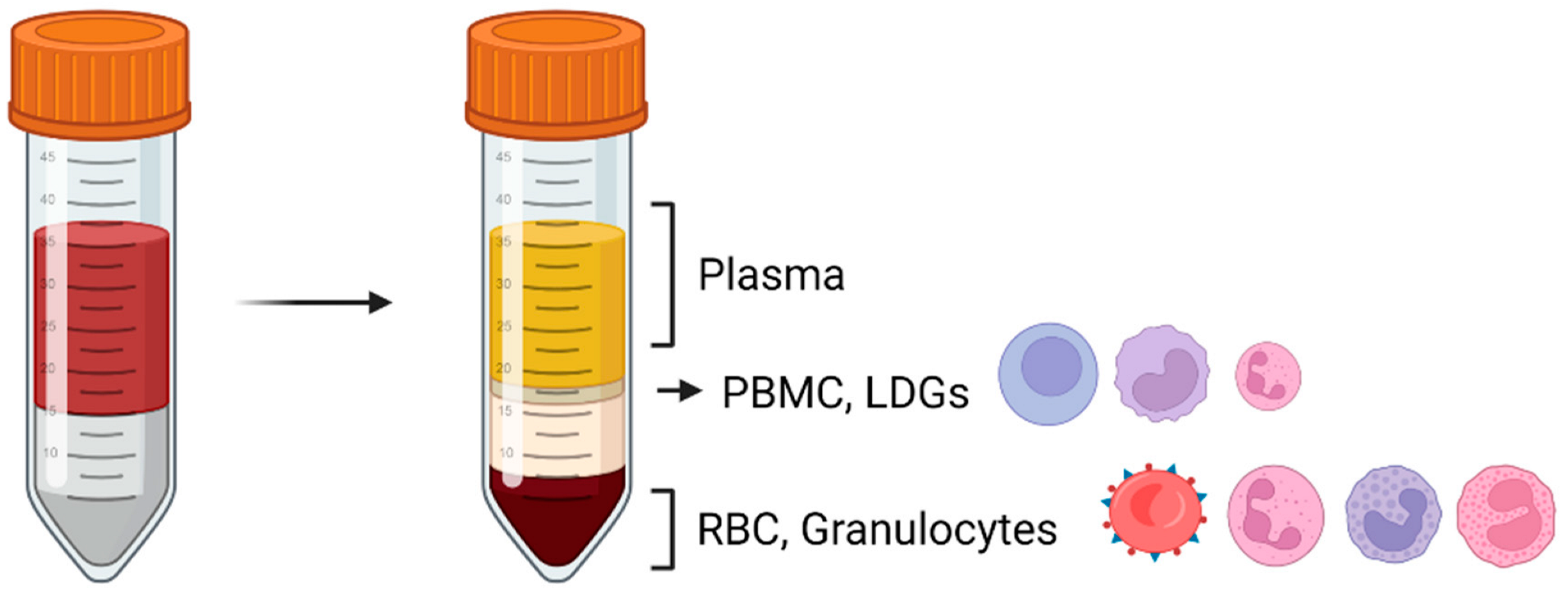{kind=link}
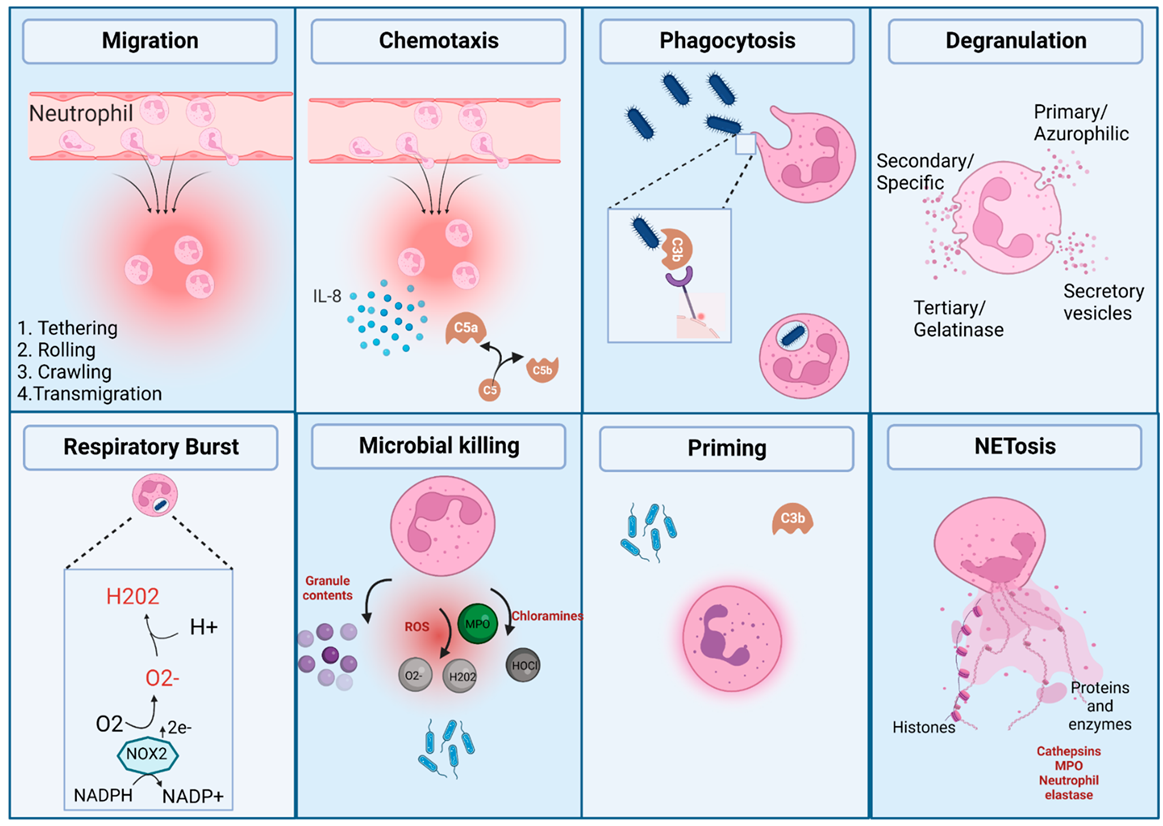{kind=link}
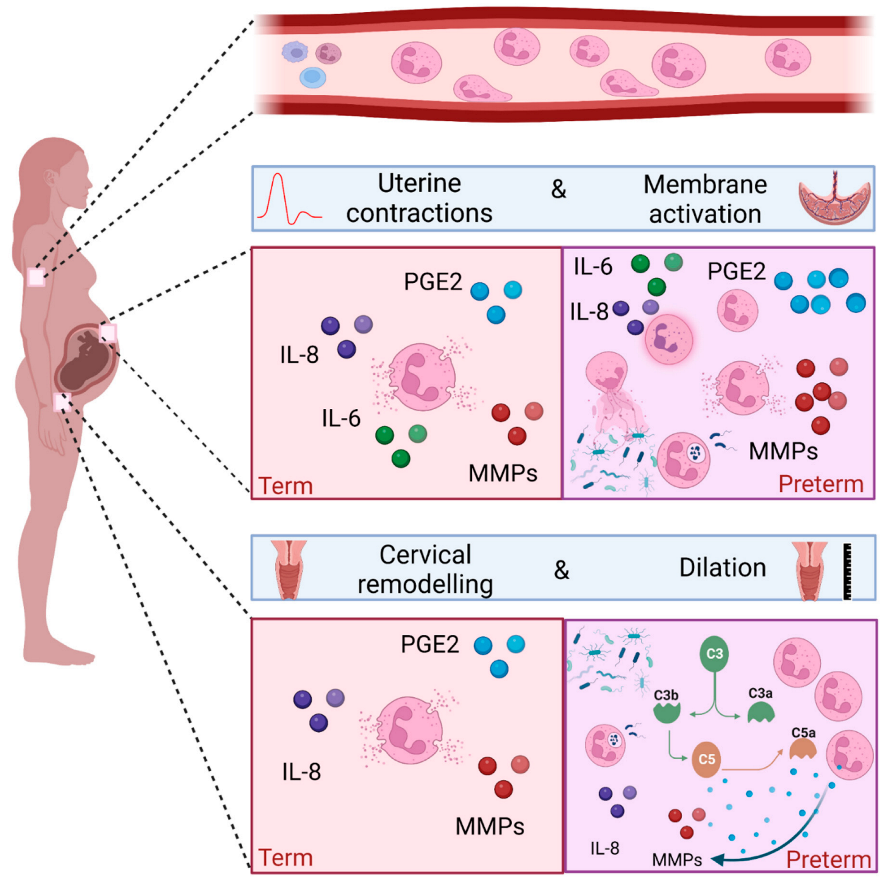{kind=link}
| Function | Primary/ Azurophilic | Secondary/ Specific | Tertiary/ Gelatinase | Secretory Vesicles |
|---|---|---|---|---|
| Microbicidal | Myeloperoxidase Lysozyme Defensins Cationic proteins Azurocidin Cap57 | Lysozyme Lactoferrin Pentraxin 3 Lipocalin 2 Haptoglobin | Cathelicidin Lysozyme | |
| Serine proteases | Neutrophil elastase Cathepsins | |||
| Metalloproteinases | Proteinases | Collagenase (MMP-8) | Gelatinase B (MMP-9) Leukolysin (MMP-25) Collagenase (MMP-8) | Leukolysin (MMP-25) Proteinase 3 (myeloblastin) |
| Acid hydrolases | Cathepsins B β-glucuronidase Glycerophosphatase N-acetylglucosaminidase α-mannosidase | β2-microglobulin | ||
| Others | Heparin binding protein (HBP) Sialidase Presenilin Granulophysin α1-antitrypsin | Histaminase Heparanase Stomatin B12 binding protein Cytochrome b C3b receptor | C3bi receptor | Heparin-binding protein (HBP) Plasma proteins (including albumin) Alkaline phosphatase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gimeno-Molina, B.; Muller, I.; Kropf, P.; Sykes, L. The Role of Neutrophils in Pregnancy, Term and Preterm Labour. Life 2022, 12, 1512. https://doi.org/10.3390/life12101512
Gimeno-Molina B, Muller I, Kropf P, Sykes L. The Role of Neutrophils in Pregnancy, Term and Preterm Labour. Life. 2022; 12(10):1512. https://doi.org/10.3390/life12101512
Chicago/Turabian StyleGimeno-Molina, Belen, Ingrid Muller, Pascale Kropf, and Lynne Sykes. 2022. "The Role of Neutrophils in Pregnancy, Term and Preterm Labour" Life 12, no. 10: 1512. https://doi.org/10.3390/life12101512