
Surveillance of Pathogenicity of Rhizoctonia solani Japanese Isolates with Varied Anastomosis Groups and Subgroups on Arabidopsis thaliana

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Plant Materials
2.2. Inoculation Tests
2.3. Phytohormones Treatments
3. Results
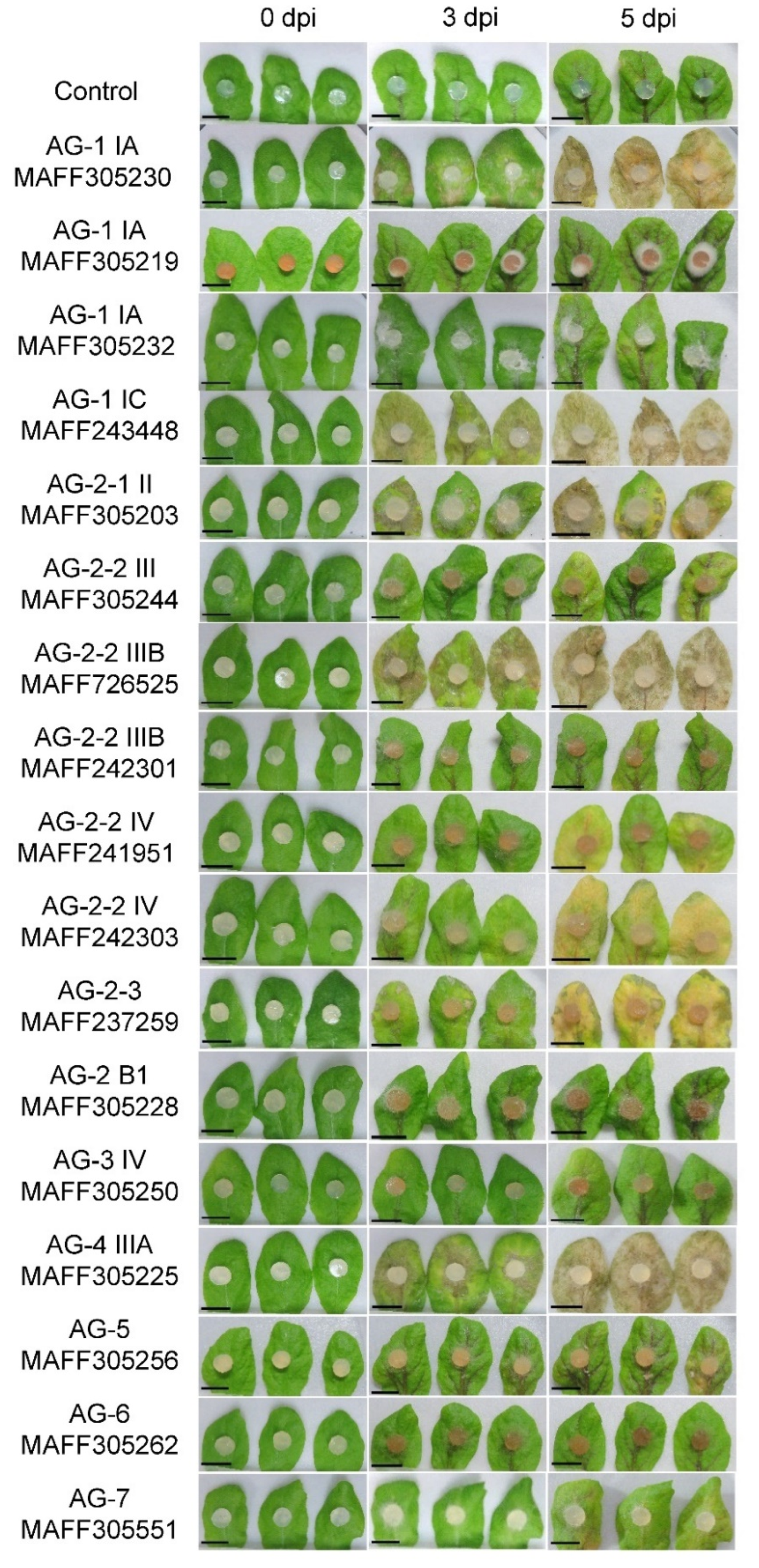
3.1. Pathogenic Behavior of the Rhizoctonia solani Isolates on Arabidopsis Leaves
3.2. Pathogenic Behavior of the Rhizoctonia solani Isolates on Arabidopsis Roots
3.3. The Effects of Exogenously Applied SA, JA, and ET on the Resistance to Rhizoctonia solani in Arabidopsis Leaves
3.4. The Effects of Exogenously Applied SA, JA, and ET on the Resistance to Rhizoctonia solani in Arabidopsis Roots
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adams, G.C. Thanatephorus cucumeris (Rhizoctonia solani) A species complex of wide host range. In Advances in Plant Pathology; Academic Press: New York, NY, USA, 1988; Volume 6, pp. 534–552. [Google Scholar]
- Baker, K.F. Types of Rhizoctonia diseases and their occurrence. In Rhizoctonia solari: Biology and Pathology; Parmeter, J.R., Jr., Ed.; University of California Press: Berkeley, CA, USA, 1970; pp. 125–148. [Google Scholar]
- Anderson, N.A. The genetics and pathology of Rhizoctonia solani. Annu. Rev. Phytopathol. 1982, 20, 329–347. [Google Scholar] [CrossRef]
- Matthew, J.S.; Brooker, J.D. The isolation and characterization of polyclonal and monoclonal antibodies to anastomosis group 8 of Rhizoctonia solani. Plant Pathol. 1991, 40, 67–77. [Google Scholar] [CrossRef]
- Webster, J.; Weber, R.W.S. Introduction to Fungi, 3rd ed.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Carling, D.E. Grouping in Rhizoctonia solani by hyphal anastomosis. In Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology and Fisease Control; Sneh, B., Jabaji-Hare, S., Neate, S., Dijst, G., Eds.; Kluwer Academic: Dordrecht, The Netherlands, 1996; pp. 37–47. [Google Scholar]
- Carling, D.E.; Kuninaga, S.; Brainard, K.A. Hyphal anastomosis reactions, rDNA-internal transcribed spacer sequences, and virulence levels among subsets of Rhizoctonia solani anastomosis group-2 (AG-2) and AG-BI. Phytopathology 2002, 92, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogoshi, A. Ecology and pathogenicity of anastomosis and intraspecific groups of Rhizoctonia solani Kühn. Annu. Rev. Phytopathol. 1987, 25, 125–143. [Google Scholar] [CrossRef]
- Sneh, B.; Burpee, L.; Ogoshi, A. Identification of Rhizoctonia Species; APS Press: St. Paul, MN, USA, 1991. [Google Scholar]
- Hua, G.K.H.; Bertier, L.; Soltaninejad, S.; Höfte, M. Cropping systems and cultural practices determine the Rhizoctonia anastomosis groups associated with Brassica spp. in Vietnam. PLoS ONE 2014, 9, e111750. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.K.; Belmar, S.B. Characterization and pathogenicity of Rhizoctonia spp. isolated from rice, soybean, and other crops grown in rotation with rice in Texas. Plant Dis. 1989, 73, 1004–1010. [Google Scholar] [CrossRef]
- Carling, D.E.; Leiner, R.H. Isolation and characterization of Rhizoctonia solani and binucleate R. solani-like fungi from aerial stems and subterranean organs of potato plants. Phytopathology 1986, 76, 725–729. [Google Scholar] [CrossRef]
- Paulitz, T.C.; Okubara, P.A.; Schillinger, W.F. First report of damping-off of canola caused by Rhizoctonia solani AG 2-1 in Washington State. Plant Dis. 2006, 90, 829. [Google Scholar] [CrossRef]
- Stodart, B.J.; Harvey, P.R.; Neate, S.M.; Melanson, D.L.; Scott, E.S. Genetic variation and pathogenicity of anastomosis group 2 isolates of Rhizoctonia solani in Australia. Mycol. Res. 2007, 111, 891–900. [Google Scholar] [CrossRef]
- Ozer, G.; Imren, M.; Bozoglu, T.; Dababat, A.A. First report of Rhizoctonia solani AG2-1 on roots of wheat in Kazakhstan. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Bolkan, H.A.; Ribeiro, W.R.C. Anastomosis groups and pathogenicity of Rhizoctonia solani isolates from Brazil. Plant Dis. 1985, 69, 599–601. [Google Scholar] [CrossRef] [Green Version]
- Keijer, J.; Korsman, M.G.; Dullemans, A.M.; Houterman, P.M.; De Bree, J.; Van Silfhout, C.H. In vitro analysis of host plant specificity in Rhizoctonia solani. Plant Pathol. 1997, 46, 659–669. [Google Scholar] [CrossRef]
- Jaaffar, A.K.M.; Paulitz, T.C.; Schroeder, K.L.; Thomashow, L.S.; Weller, D.M. Molecular characterization, morphological characteristics, virulence, and geographic distribution of Rhizoctonia spp. in Washington State. Phytopathology 2016, 106, 459–473. [Google Scholar] [CrossRef] [Green Version]
- Pannecoucque, J.; Hofte, M. Interactions between cauliflower and Rhizoctonia anastomosis groups with different levels of pathogenicity. BMC Plant Biol. 2009, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Kidd, B.N.; Foley, R.; Singh, K.B.; Anderson, J.P. Foliar resistance to Rhizoctonia solani in Arabidopsis is compromised by simultaneous loss of ethylene, jasmonate and PEN2 mediated defense pathways. Sci. Rep. 2021, 11, 2546. [Google Scholar] [CrossRef] [PubMed]
- Foley, R.C.; Gleason, C.A.; Anderson, J.P.; Hamann, T.; Singh, K.B. Genetic and genomic analysis of Rhizoctonia solani interactions with Arabidopsis; Evidence of resistance mediated through NADPH oxidases. PLoS ONE 2013, 8, e56814. [Google Scholar]
- Kumar, S.; Tripathi, D.; Okubara, P.A.; Tanaka, K. Purinoceptor P2K1/DORN1 enhances plant resistance against a soilborne fungal pathogen, Rhizoctonia solani. Front. Plant Sci. 2020, 11, 572920. [Google Scholar] [CrossRef] [PubMed]
- Khangura, R.K.; Barbetti, M.J.; Sweetingham, M.W. Characterization and pathogenicity of Rhizoctonia species on canola. Plant Dis. 1999, 83, 714–721. [Google Scholar] [CrossRef] [Green Version]
- Kouzai, Y.; Kimura, M.; Watanabe, M.; Kusunoki, K.; Osaka, D.; Suzuki, T.; Matsui, H.; Yamamoto, M.; Ichinose, Y.; Toyoda, K.; et al. Salicylic acid-dependent immunity contributes to resistance against Rhizoctonia solani, a necrotrophic fungal agent of sheath blight, in rice and Brachypodium distachyon. New Phytol. 2018, 217, 771–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-Microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Bostock, R.M. Signal crosstalk and induced resistance: Straddling the line between cost and benefit. Annu. Rev. Phytopathol. 2005, 43, 545–580. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Muthamilarasan, M.; Prasad, M. Plant innate immunity: An updated insight into defense mechanism. J. Biosci. 2013, 38, 433–449. [Google Scholar] [CrossRef]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.-J. Signaling crosstalk between salicylic acid and ethylene/jasmonate in plant defense: Do we understand what they are whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef] [Green Version]
- Parween, D.; Sultan, E.; Dalei, K.; Sahu, B.B. Arabidopsis nonhost resistance gene PENETRATION 2 is involved in disruption of cushion formation by Rhizoctonia solani during early infection process. Australas. Plant Pathol. 2021, 50, 281–292. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Carp, M.-J.; Zuchman, R.; Ziv, T.; Horwitz, B.A.; Gepstein, S. Proteomics of the response of Arabidopsis thaliana to infection with Alternaria brassicicola. J. Proteom. 2010, 73, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Bowyer, P.; Clarke, B.R.; Lunness, P.; Daniels, M.J.; Osbourn, A.E. Host range of a plant pathogenic fungus determined by a saponin detoxifying enzyme. Science 1995, 267, 371–374. [Google Scholar] [CrossRef]
- Maeda, S.; Dubouzet, J.G.; Kondou, Y.; Jikumaru, Y.; Seo, S.; Oda, K.; Matsui, M.; Hirochika, H.; Mori, M. The rice CYP78A gene BSR2 confers resistance to Rhizoctonia solani and affects seed size and growth in Arabidopsis and rice. Sci. Rep. 2019, 9, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouzai, Y.; Shimizu, M.; Inoue, K.; Uehara-Yamaguchi, Y.; Takahagi, K.; Nakayama, R.; Matsuura, T.; Mori, I.C.; Hirayama, T.; Abdelsalam, S.S.H.; et al. BdWRKY38 is required for the incompatible interaction of Brachypodium distachyon with the necrotrophic fungus Rhizoctonia solani. Plant J. 2020, 104, 995–1008. [Google Scholar] [CrossRef] [PubMed]
- Abdelsalam, S.S.H.; Kouzai, Y.; Watanabe, M.; Inoue, K.; Matsui, H.; Yamamoto, M.; Ichinose, Y.; Toyoda, K.; Tsuge, S.; Mochida, K.; et al. Identification of effector candidate genes of Rhizoctonia solani AG-1 IA expressed during infection in Brachypodium distachyon. Sci. Rep. 2020, 10, 14889. [Google Scholar] [CrossRef] [PubMed]
- Kouzai, Y.; Noutoshi, Y.; Inoue, K.; Shimizu, M.; Onda, Y.; Mochida, K. Benzothiadiazole, a plant defense inducer, negatively regulates sheath blight resistance in Brachypodium distachyon. Sci. Rep. 2018, 8, 17358. [Google Scholar] [CrossRef] [PubMed]
- Elad, Y. Regulators of ethylene biosynthesis or activity as a tool for reducing susceptibility of host plant tissues to infection by Botrytis cinerea. Neth. J. Plant Pathol. 1993, 99, 105–113. [Google Scholar] [CrossRef]
- Panter, S.N.; Jones, D.A. Age-Related resistance to plant pathogens. Adv. Bot. Res. 2002, 38, 251–280. [Google Scholar]
- van Loon, L.C.; Geraats, B.P.J.; Linthorst, H.J.M. Ethylene as a modulator of disease resistance in plants. Trends Plant Sci. 2006, 11, 184–191. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AGs | MAFF Number 1 | Name | Growth (Days) 2 | Source | Infection Severity 3 | |
|---|---|---|---|---|---|---|
| Leaves | Roots | |||||
| AG-1 IA | 305230 | C-325 | 3 | Rice | +++ | ++ |
| AG-1 IA | 305219 | C-54 | 6 | Rice | − | − |
| AG-1 IA | 305232 | C-501 | 3 | Sudangrass | − | − |
| AG-1 IC | 243448 | RD1 | 3 | Carrot | +++ | +++ |
| AG-2-1 II | 305203 | 6 | 7 | Six-rowed barley | ++ | − |
| AG-2-2 IIIB | 305244 | C-329 | 7 | Rice | − | ++ |
| AG-2-2 IIIB | 726525 | RS-B | 3 | Broccoli | +++ | +++ |
| AG-2-2 IIIB | 242301 | WLS81 | 4 | Welch onion | − | +++ |
| AG-2-2 IV | 241951 | SBF1 | 4 | Broccoli | ++ | +++ |
| AG-2-2 IV | 242303 | SD1 | 4 | Soybean | ++ | +++ |
| AG-2-3 | 237259 | H5-210 | 7 | Wheat | ++ | − |
| AG-2 BI | 305228 | SH-1-2 | 7 | Soil | − | − |
| AG-3 IV | 305250 | C564 | 14 | Potato | − | − |
| AG-4 IIIA | 305225 | BO-3 | 3 | Cauliflower | +++ | +++ |
| AG-5 | 305256 | SH-30 | 7 | Soil | + | − |
| AG-6 | 305262 | UB-7-1-A | 4 | Soil | − | − |
| AG-7 | 305551 | 1529 | 3 | Radish field soil | + | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelghany, M.M.A.; Kurikawa, M.; Watanabe, M.; Matsui, H.; Yamamoto, M.; Ichinose, Y.; Toyoda, K.; Kouzai, Y.; Noutoshi, Y. Surveillance of Pathogenicity of Rhizoctonia solani Japanese Isolates with Varied Anastomosis Groups and Subgroups on Arabidopsis thaliana. Life 2022, 12, 76. https://doi.org/10.3390/life12010076
Abdelghany MMA, Kurikawa M, Watanabe M, Matsui H, Yamamoto M, Ichinose Y, Toyoda K, Kouzai Y, Noutoshi Y. Surveillance of Pathogenicity of Rhizoctonia solani Japanese Isolates with Varied Anastomosis Groups and Subgroups on Arabidopsis thaliana. Life. 2022; 12(1):76. https://doi.org/10.3390/life12010076
Chicago/Turabian StyleAbdelghany, Mai Mohsen Ahmed, Maria Kurikawa, Megumi Watanabe, Hidenori Matsui, Mikihiro Yamamoto, Yuki Ichinose, Kazuhiro Toyoda, Yusuke Kouzai, and Yoshiteru Noutoshi. 2022. "Surveillance of Pathogenicity of Rhizoctonia solani Japanese Isolates with Varied Anastomosis Groups and Subgroups on Arabidopsis thaliana" Life 12, no. 1: 76. https://doi.org/10.3390/life12010076