
Caveolin-1 Down-Regulation Reduces VEGF-A Secretion Induced by IGF-1 in ARPE-19 Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. RNA Silencing
2.3. Immunoblotting Analysis
2.4. RNA Isolation, cDNA and qRT-PCR
2.5. VEGF-A Secretion
2.6. Scratch Assay
2.7. Cell Viability
2.8. Statistical Analysis
3. Results
3.1. Caveolin-1 Expression and Down-Regulation of Cav-1 with siRNA
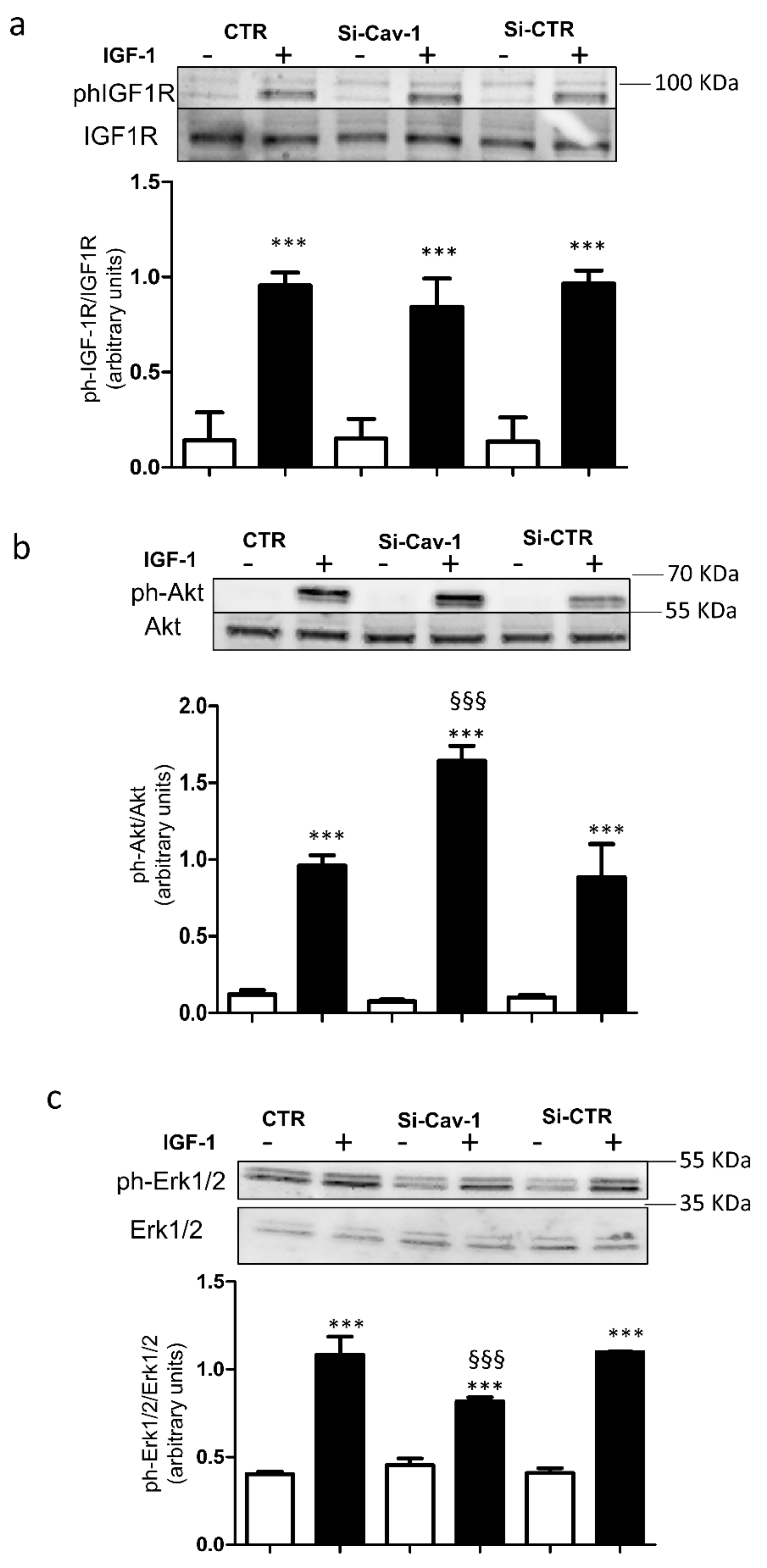
3.2. Effects of Caveolin-1 Down-Regulation on Intracellular Signaling of IGF-1
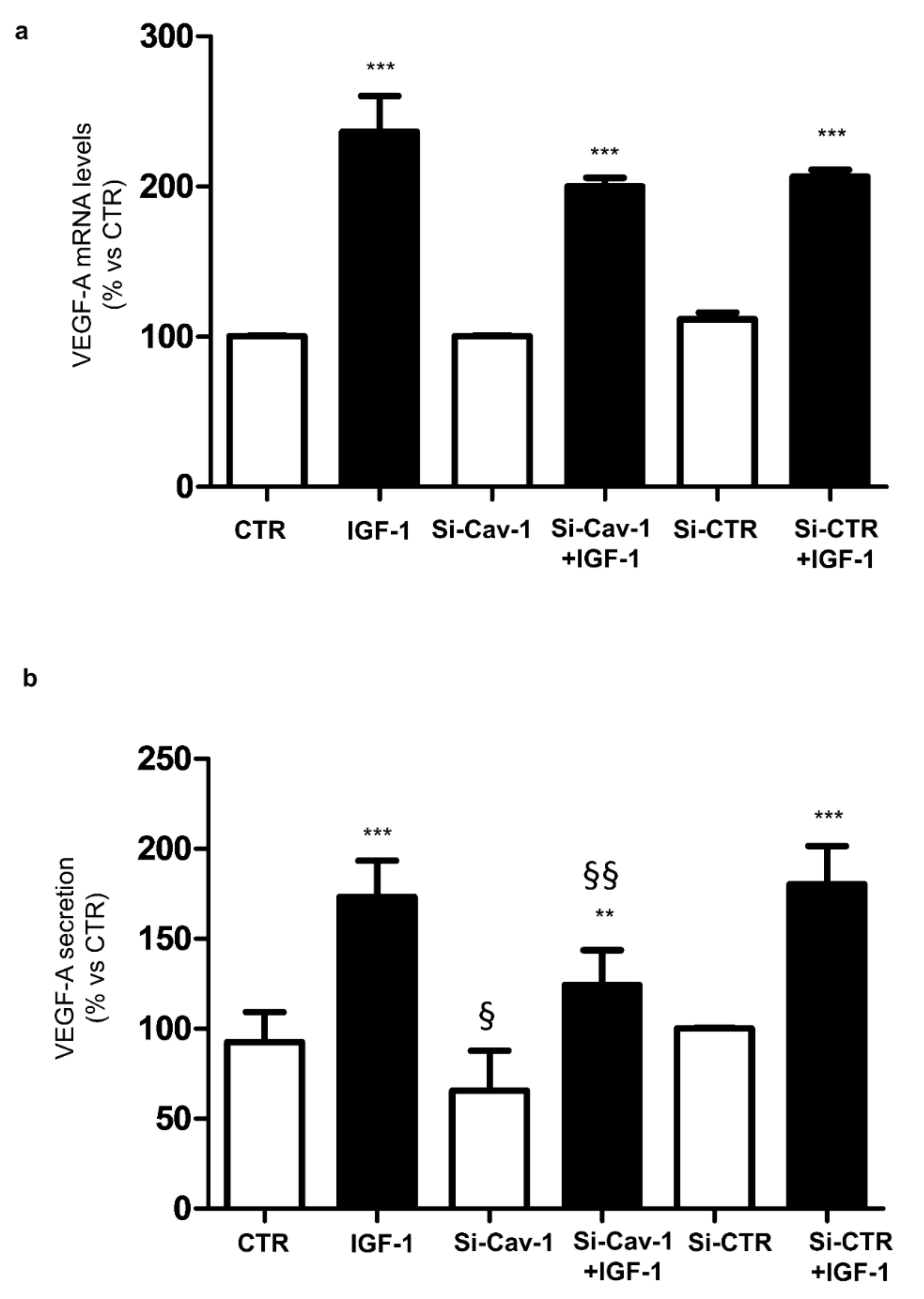
3.3. Effects of Caveolin-1 Down-Regulation on VEGF-A Expression and Secretion
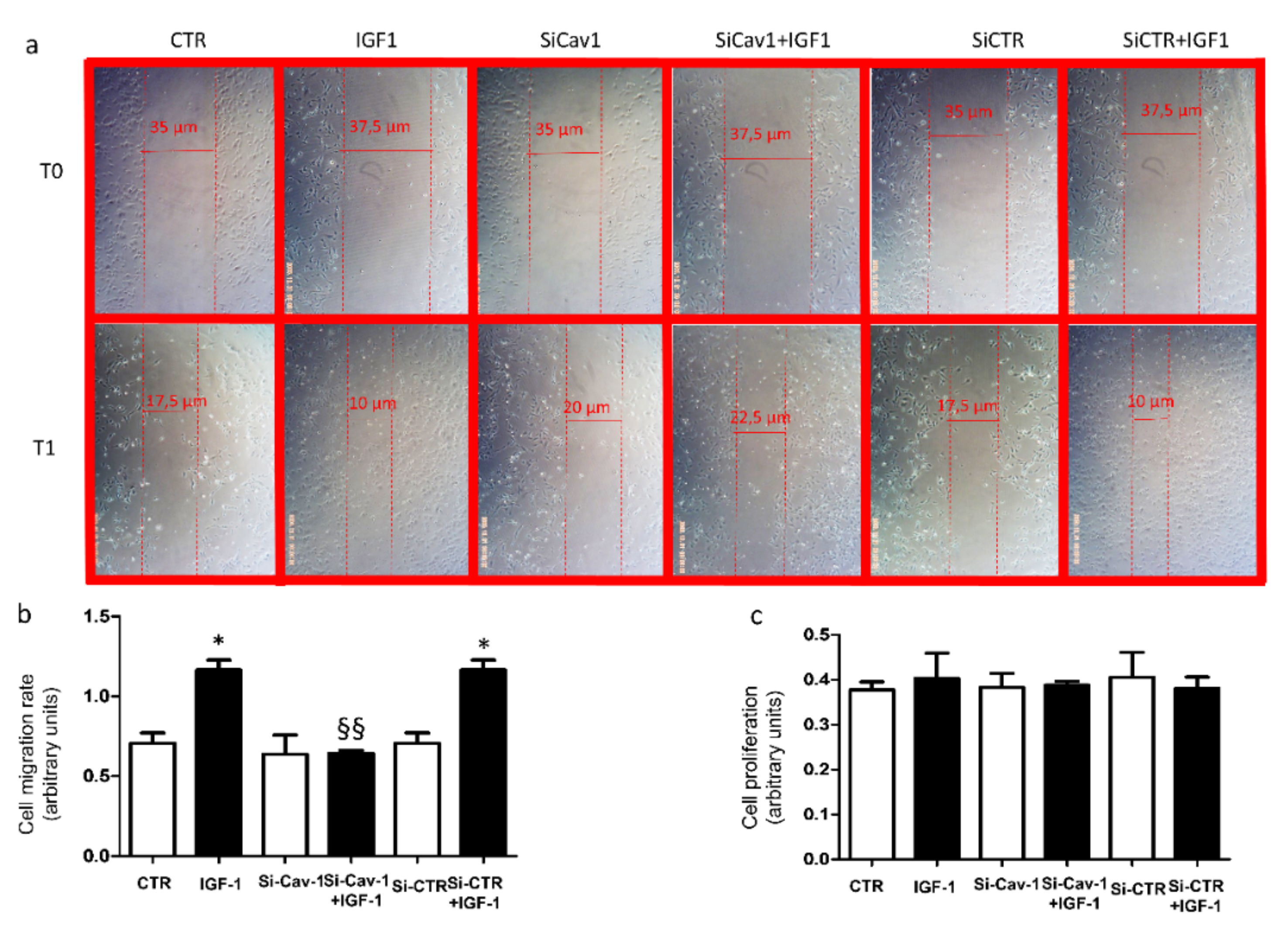
3.4. Effects of ARPE-19 Secretome on Endothelial Cell Activation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simons, K.; Toomre, D. Lipid rafts and signal transduction. Nat. Rev. Mol. Cell Biol. 2000, 1, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Bok, D. The retinal pigment epithelium: A versatile partner in vision. J. Cell Sci. Suppl. 1993, 17, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamis, A.P.; Shima, D.T.; Yeo, K.T.; Yeo, T.K.; Brown, L.F.; Berse, B.; D’Amore, P.A.; Folkman, J. Synthesis and secretion of vascular permeability factor/vascular endothelial growth factor by human retinal pigment epithelial cells. Biochem. Biophys. Res. Commun. 1993, 193, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Strauss, O. The retinal pigment epithelium in visual function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef] [Green Version]
- Sethna, S.; Chamakkala, T.; Gu, X.; Thompson, T.C.; Cao, G.; Elliott, M.H.; Finnemann, S.C. Regulation of Phagolysosomal Digestion by Caveolin-1 of the Retinal Pigment Epithelium Is Essential for Vision. J. Biol. Chem. 2016, 291, 6494–6506. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; McClellan, M.E.; Tanito, M.; Garteiser, P.; Towner, R.; Bissig, D.; Berkowitz, B.A.; Fliesler, S.J.; Woodruff, M.L.; Fain, G.L.; et al. Loss of caveolin-1 impairs retinal function due to disturbance of subretinal microenvironment. J. Biol. Chem. 2012, 287, 16424–16434. [Google Scholar]
- Tian, X.F.; Xia, X.B.; Xu, H.Z.; Xiong, S.Q.; Jiang, J. Caveolin-1 expression regulates blood-retinal barrier permeability and retinal neovascularization in oxygen-induced retinopathy. Clin. Exp. Ophthalmol. 2012, 40, e58–e66. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Lin, X.; Tang, Z.; Lee, C.; Tian, G.; Du, Y.; Yin, X.; Ren, X.; Huang, L.; Ye, Z.; et al. Critical role of caveolin-1 in ocular neovascularization and multitargeted antiangiogenic effects of cavtratin via JNK. Proc. Natl. Acad. Sci. USA 2017, 114, 10737–10742. [Google Scholar] [CrossRef] [Green Version]
- Lambooij, A.C.; van Wely, K.H.; Lindenbergh-Kortleve, D.J.; Kuijpers, R.W.; Kliffen, M.; Mooy, C.M. Insulin-like growth factor-I and its receptor in neovascular age-related macular degeneration. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2192–2198. [Google Scholar] [CrossRef]
- Punglia, R.S.; Lu, M.; Hsu, J.; Kuroki, M.; Tolentino, M.J.; Keough, K.; Levy, A.P.; Levy, N.S.; Goldberg, M.A.; D’Amato, R.J.; et al. Regulation of vascular endothelial growth factor expression by insulin-like growth factor I. Diabetes 1997, 46, 1619–1626. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.Y.; Kothary, P.C.; Verkade, A.J.; Reed, D.M.; Del Monte, M.A. MAP kinase pathway is involved in IGF-1-stimulated proliferation of human retinal pigment epithelial cells (hRPE). Curr. Eye Res. 2009, 34, 867–876. [Google Scholar] [CrossRef]
- Zheng, W.; Meng, Q.; Wang, H.; Yan, F.; Little, P.J.; Deng, X.; Lin, S. IGF-1-Mediated Survival from Induced Death of Human Primary Cultured Retinal Pigment Epithelial Cells Is Mediated by an Akt-Dependent Signaling Pathway. Mol. Neurobiol. 2018, 55, 1915–1927. [Google Scholar] [CrossRef] [PubMed]
- Maggi, D.; Biedi, C.; Segat, D.; Barbero, D.; Panetta, D.; Cordera, R. IGF-I induces caveolin 1 tyrosine phosphorylation and translocation in the lipid rafts. Biochem. Biophys. Res. Commun. 2002, 295, 1085–1089. [Google Scholar] [CrossRef]
- Panetta, D.; Biedi, C.; Repetto, S.; Cordera, R.; Maggi, D. IGF-I regulates caveolin 1 and IRS1 interaction in caveolae. Biochem. Biophys. Res. Commun. 2004, 316, 240–243. [Google Scholar] [CrossRef]
- Grada, A.; Otero-Vinas, M.; Prieto-Castrillo, F.; Obagi, Z.; Falanga, V. Research Techniques Made Simple: Analysis of Collective Cell Migration Using the Wound Healing Assay. J. Investig. Dermatol. 2017, 137, e11–e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brannmark, C.; Kay, E.I.; Ortegren Kugelberg, U.; Chanclon, B.; Shrestha, M.M.; Wernstedt Asterholm, I.; Stralfors, P.; Olofsson, C.S. Adiponectin is secreted via caveolin 1-dependent mechanisms in white adipocytes. J. Endocrinol. 2020, 247, 25–38. [Google Scholar] [CrossRef]
- Haddad, D.; Al Madhoun, A.; Nizam, R.; Al-Mulla, F. Role of Caveolin-1 in Diabetes and Its Complications. Oxidative Med. Cell. Longev. 2020, 2020, 9761539. [Google Scholar] [CrossRef] [Green Version]
- Burgos, R.; Mateo, C.; Canton, A.; Hernandez, C.; Mesa, J.; Simo, R. Vitreous levels of IGF-I, IGF binding protein 1, and IGF binding protein 3 in proliferative diabetic retinopathy: A case-control study. Diabetes Care 2000, 23, 80–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.E.; Shen, W.; Perruzzi, C.; Soker, S.; Kinose, F.; Xu, X.; Robinson, G.; Driver, S.; Bischoff, J.; Zhang, B.; et al. Regulation of vascular endothelial growth factor-dependent retinal neovascularization by insulin-like growth factor-1 receptor. Nat. Med. 1999, 5, 1390–1395. [Google Scholar] [CrossRef]
- Rosenthal, R.; Wohlleben, H.; Malek, G.; Schlichting, L.; Thieme, H.; Bowes Rickman, C.; Strauss, O. Insulin-like growth factor-1 contributes to neovascularization in age-related macular degeneration. Biochem. Biophys. Res. Commun. 2004, 323, 1203–1208. [Google Scholar] [CrossRef]
- Salani, B.; Briatore, L.; Garibaldi, S.; Cordera, R.; Maggi, D. Caveolin-1 down-regulation inhibits insulin-like growth factor-I receptor signal transduction in H9C2 rat cardiomyoblasts. Endocrinology 2008, 149, 461–465. [Google Scholar] [CrossRef] [Green Version]
- Hakuno, F.; Takahashi, S.I. IGF1 receptor signaling pathways. J. Mol. Endocrinol. 2018, 61, T69–T86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, R.; Yan, F.; Zeng, Z.; Wang, H.; Qiu, K.; Xu, J.; Zheng, W. Insulin-like growth factor-1 activates PI3K/Akt signalling to protect human retinal pigment epithelial cells from amiodarone-induced oxidative injury. Br. J. Pharmacol. 2018, 175, 125–139. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liao, S.; Geng, R.; Zheng, Y.; Liao, R.; Yan, F.; Thrimawithana, T.; Little, P.J.; Feng, Z.P.; Lazarovici, P.; et al. IGF-1 signaling via the PI3K/Akt pathway confers neuroprotection in human retinal pigment epithelial cells exposed to sodium nitroprusside insult. J. Mol. Neurosci. MN 2015, 55, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Courtaut, F.; Scagliarini, A.; Aires, V.; Cornebise, C.; Pais de Barros, J.P.; Olmiere, C.; Delmas, D. VEGF-R2/Caveolin-1 Pathway of Undifferentiated ARPE-19 Retina Cells: A Potential Target as Anti-VEGF-A Therapy in Wet AMD by Resvega, an Omega-3/Polyphenol Combination. Int. J. Mol. Sci. 2021, 22, 12. [Google Scholar] [CrossRef]
- Klettner, A.; Roider, J. Constitutive and oxidative-stress-induced expression of VEGF in the RPE are differently regulated by different Mitogen-activated protein kinases. Graefe’s Arch. Clin. Exp. Ophthalmol. = Albrecht Von Graefes Arch. Fur Klin. Und Exp. Ophthalmol. 2009, 247, 1487–1492. [Google Scholar] [CrossRef] [PubMed]
- Ponnalagu, M.; Subramani, M.; Jayadev, C.; Shetty, R.; Das, D. Retinal pigment epithelium-secretome: A diabetic retinopathy perspective. Cytokine 2017, 95, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Reagan, A.M.; McClellan, M.E.; Elliott, M.H. Caveolins and caveolae in ocular physiology and pathophysiology. Prog. Retin. Eye Res. 2017, 56, 84–106. [Google Scholar] [CrossRef] [Green Version]
- Mora, R.C.; Bonilha, V.L.; Shin, B.C.; Hu, J.; Cohen-Gould, L.; Bok, D.; Rodriguez-Boulan, E. Bipolar assembly of caveolae in retinal pigment epithelium. Am. J. Physiol. Cell Physiol. 2006, 290, C832–C843. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Fliesler, S.J.; Zhao, Y.Y.; Stallcup, W.B.; Cohen, A.W.; Elliott, M.H. Loss of caveolin-1 causes blood-retinal barrier breakdown, venous enlargement, and mural cell alteration. Am. J. Pathol. 2014, 184, 541–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagasaka, Y.; Kaneko, H.; Ye, F.; Kachi, S.; Asami, T.; Kato, S.; Takayama, K.; Hwang, S.J.; Kataoka, K.; Shimizu, H.; et al. Role of Caveolin-1 for Blocking the Epithelial-Mesenchymal Transition in Proliferative Vitreoretinopathy. Investig. Ophthalmol. Vis. Sci. 2017, 58, 221–229. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puddu, A.; Sanguineti, R.; Maggi, D. Caveolin-1 Down-Regulation Reduces VEGF-A Secretion Induced by IGF-1 in ARPE-19 Cells. Life 2022, 12, 44. https://doi.org/10.3390/life12010044
Puddu A, Sanguineti R, Maggi D. Caveolin-1 Down-Regulation Reduces VEGF-A Secretion Induced by IGF-1 in ARPE-19 Cells. Life. 2022; 12(1):44. https://doi.org/10.3390/life12010044
Chicago/Turabian StylePuddu, Alessandra, Roberta Sanguineti, and Davide Maggi. 2022. "Caveolin-1 Down-Regulation Reduces VEGF-A Secretion Induced by IGF-1 in ARPE-19 Cells" Life 12, no. 1: 44. https://doi.org/10.3390/life12010044