
Sleep Apnea in Idiopathic Pulmonary Fibrosis: A Molecular Investigation in an Experimental Model of Fibrosis and Intermittent Hypoxia

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Animals
2.3. Lung Fibrosis Induction
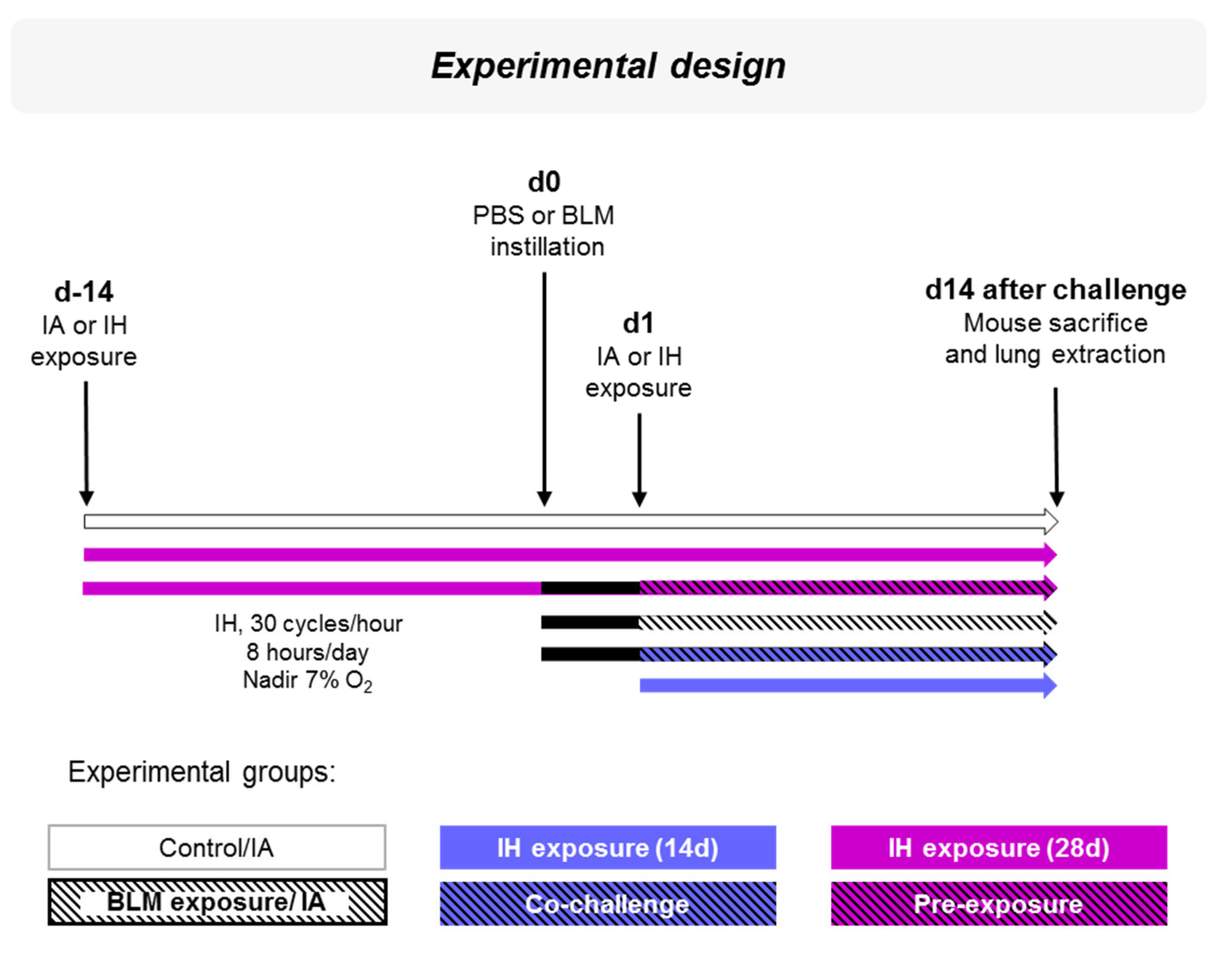
2.4. Experimental Design
2.5. Intermittent Hypoxia Exposure
2.6. Lung Extraction
2.7. Histological Staining
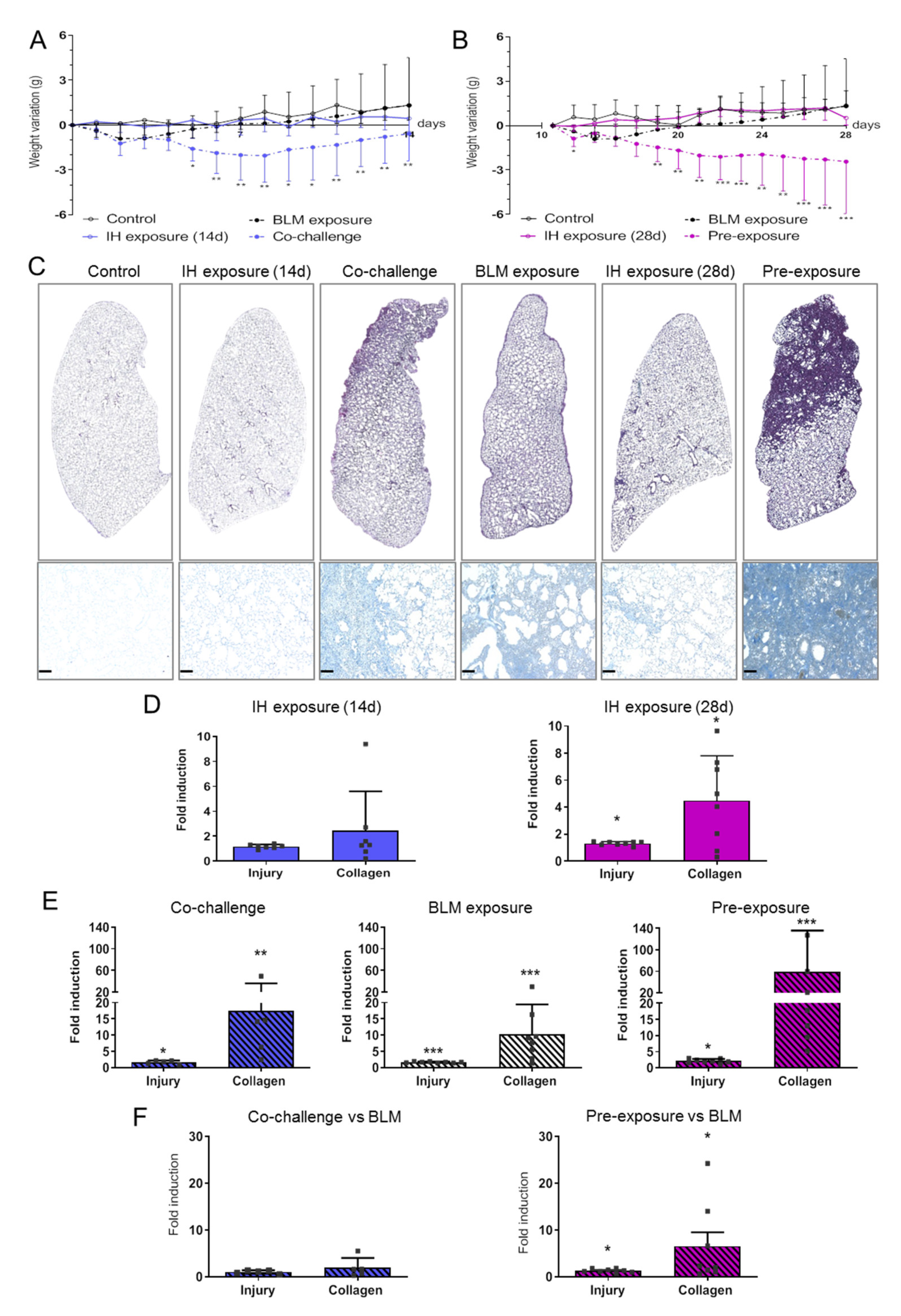
2.8. Lung Injury and Fibrosis Quantification
2.9. Quantitative Real-Time Reverse Transcription–Polymerase Chain Reaction
2.10. Lung Protein Extraction and Immunoblotting Analysis
2.11. Data and Statistical Analysis
3. Results
3.1. Intermittent Hypoxia Promotes Tissue Remodelling and Collagen Deposition That Worsens BLM-Induced Lung Fibrosis
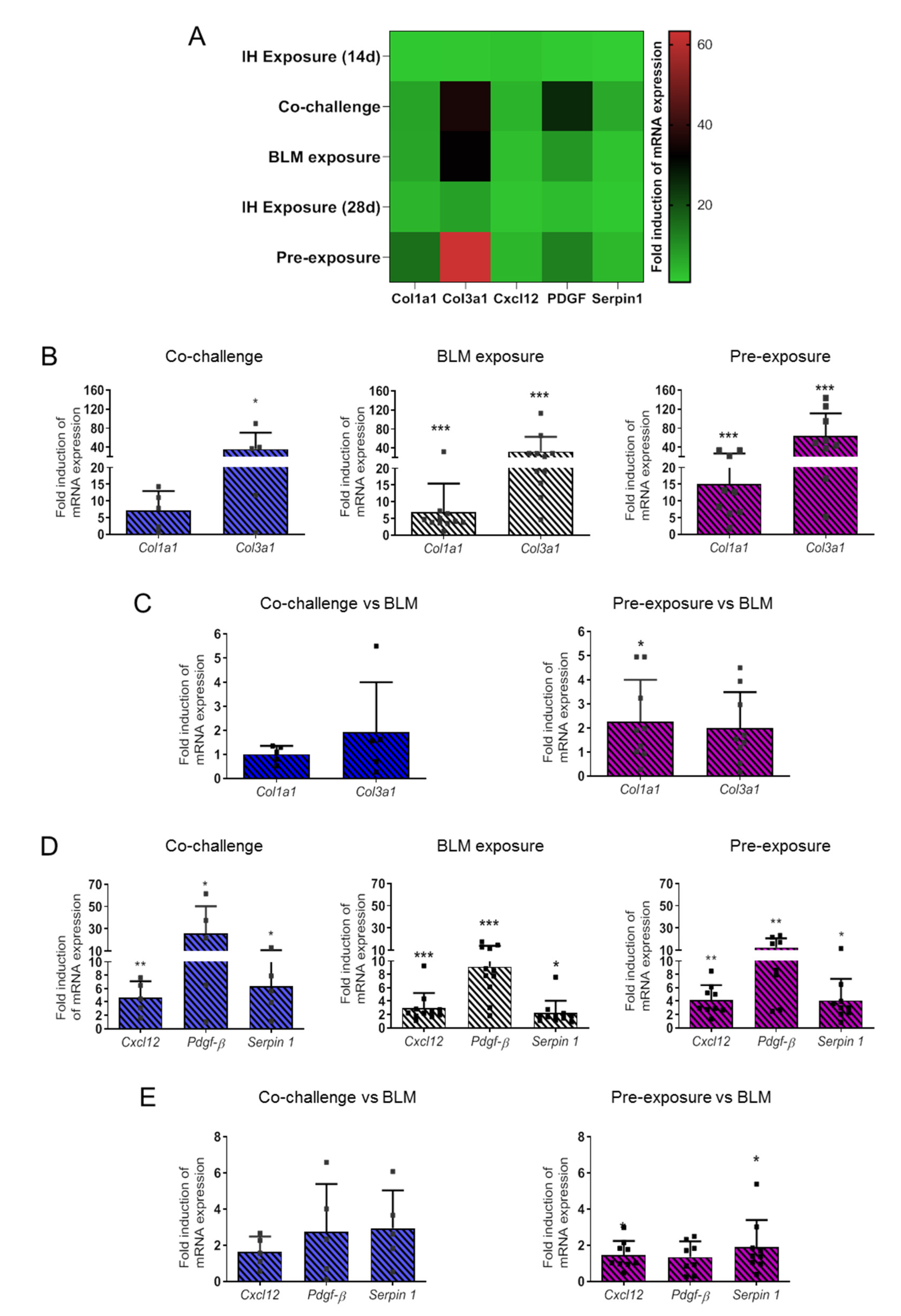
3.2. Intermittent Hypoxia Increases Collagen and the Production of Pro-Fibrotic Mediators and Contributes to its Worsening Effect on BLM-Induced Lung Fibrosis
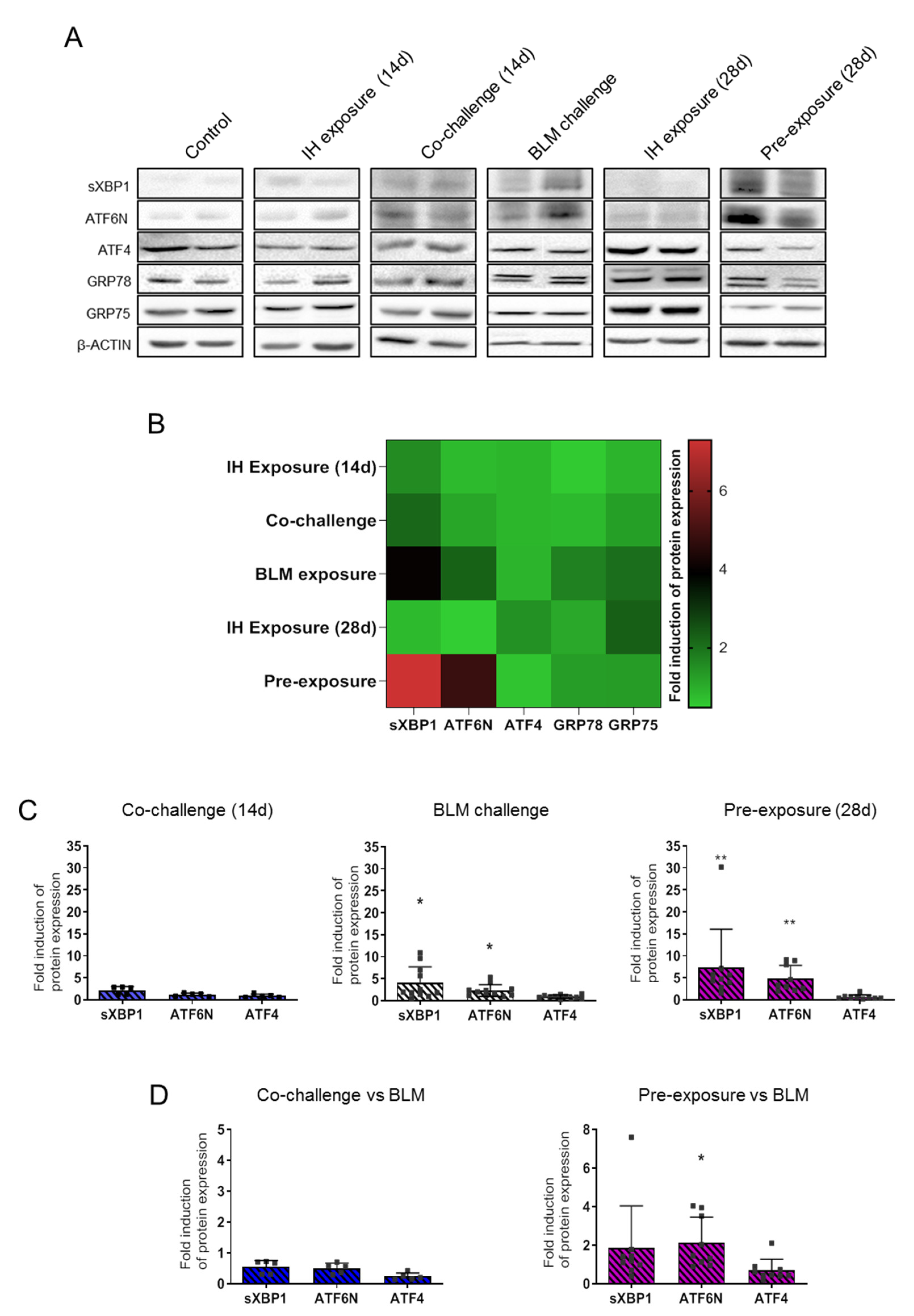
3.3. Intermittent Hypoxia Modulates ER Stress Response That Enhances BLM Impact on ER Stress Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Barratt, S.L.; Creamer, A.; Hayton, C.; Chaudhuri, N. Idiopathic Pulmonary Fibrosis (IPF): An Overview. J. Clin. Med. 2018, 7, 201. [Google Scholar] [CrossRef] [Green Version]
- Selman, M.; King, T.E.; Pardo, A. Idiopathic Pulmonary Fibrosis: Prevailing and Evolving Hypotheses about Its Pathogenesis and Implications for Therapy. Ann. Intern. Med. 2001, 134, 136–151. [Google Scholar] [CrossRef]
- Mermigkis, C.M.; Tsakanika, K.; Polychronopoulos, V.; Karagianidis, N.; Mermigkis, D.; Bouros, D. Expression of bcl-2 protein in bronchoalveolar lavage cell populations from patients with idiopathic pulmonary fibrosis. Acta Cytol. 2001, 45, 914–918. [Google Scholar] [CrossRef]
- Plataki, M.; Koutsopoulos, A.V.; Darivianaki, K.; Delides, G.; Siafakas, N.M.; Bouros, D. Expression of Apoptotic and Antiapoptotic Markers in Epithelial Cells in Idiopathic Pulmonary Fibrosis. Chest 2005, 127, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Goldmann, T.; Zissel, G.; Watz, H.; Drömann, D.; Reck, M.; Kugler, C.; Rabe, K.F.; Marwitz, S. Human alveolar epithelial cells type II are capable of TGFβ-dependent epithelial-mesenchymal-transition and collagen-synthesis. Respir. Res. 2018, 19, 138. [Google Scholar] [CrossRef]
- Willis, B.C.; Liebler, J.M.; Luby-Phelps, K.; Nicholson, A.G.; Crandall, E.D.; du Bois, R.M.; Borok, Z. Induction of Epithelial-Mesenchymal Transition in Alveolar Epithelial Cells by Transforming Growth Factor-β1: Potential Role in Idiopathic Pulmonary Fibrosis. Am. J. Pathol. 2005, 166, 1321–1332. [Google Scholar] [CrossRef]
- Allen, J.T.; A Spiteri, M. Growth factors in idiopathic pulmonary fibrosis: Relative roles. Respir. Res. 2002, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Gille, T.; Didier, M.; Boubaya, M.; Moya, L.; Sutton, A.; Carton, Z.; Baran-Marszak, F.; Sadoun-Danino, D.; Israël-Biet, D.; Cottin, V.; et al. Obstructive sleep apnoea and related comorbidities in incident idiopathic pulmonary fibrosis. Eur. Respir. J. 2017, 49, 1601934. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, L.H.; Mason, W.R.; Parnell, J.A.; Rice, T.; Loyd, J.; Milstone, A.P.; Collard, H.R.; Malow, B.A. Obstructive Sleep Apnea Is Common in Idiopathic Pulmonary Fibrosis. Chest 2009, 136, 772–778. [Google Scholar] [CrossRef] [Green Version]
- Mermigkis, C.; Bouloukaki, I.; Antoniou, K.M.; Mermigkis, D.; Psathakis, K.; Giannarakis, I.; Varouchakis, G.; Siafakas, N.M.; Schiza, S.E. CPAP therapy in patients with idiopathic pulmonary fibrosis and obstructive sleep apnea: Does it offer a better quality of life and sleep? Sleep Breath. 2013, 17, 1137–1143. [Google Scholar] [CrossRef]
- Young, T.; Palta, M.; Dempsey, J.; Skatrud, J.; Weber, S.; Badr, S. The Occurrence of Sleep-Disordered Breathing among Middle-Aged Adults. N. Engl. J. Med. 1993, 328, 1230–1235. [Google Scholar] [CrossRef] [Green Version]
- Badran, M.; Ayas, N.; Laher, I. Cardiovascular Complications of Sleep Apnea: Role of Oxidative Stress. Oxidative Med. Cell. Longev. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Dewan, N.A.; Nieto, F.J.; Somers, V.K. Intermittent Hypoxemia and OSA. Chest 2015, 147, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Sforza, E.; Roche, F. Chronic intermittent hypoxia and obstructive sleep apnea: An experimental and clinical approach. Hypoxia 2016, 4, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Kim, I.K.; Yeo, C.D.; Kang, H.H.; Ban, W.H.; Kwon, H.Y.; Lee, S.H. Effects of chronic intermittent hypoxia caused by obstructive sleep apnea on lipopolysaccharide-induced acute lung injury. Exp. Lung Res. 2020, 46, 1–11. [Google Scholar] [CrossRef]
- Braun, R.; Broytman, O.; Braun, F.M.; Brinkman, J.A.; Clithero, A.; Modi, D.; Pegelow, D.F.; Eldridge, M.; Teodorescu, M. Chronic intermittent hypoxia worsens bleomycin-induced lung fibrosis in rats. Respir. Physiol. Neurobiol. 2018, 256, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Gille, T.; Didier, M.; Rotenberg, C.; Delbrel, E.; Marchant, D.; Sutton, A.; Dard, N.; Haine, L.; Voituron, N.; Bernaudin, J.-F.; et al. Intermittent Hypoxia Increases the Severity of Bleomycin-Induced Lung Injury in Mice. Oxidative Med. Cell. Longev. 2018, 2018, 1–13. [Google Scholar] [CrossRef]
- Korfei, M.; Ruppert, C.; Mahavadi, P.; Henneke, I.; Markart, P.; Koch, M.; Lang, G.; Fink, L.; Bohle, R.-M.; Seeger, W.; et al. Epithelial Endoplasmic Reticulum Stress and Apoptosis in Sporadic Idiopathic Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 2008, 178, 838–846. [Google Scholar] [CrossRef] [Green Version]
- Delbrel, E.; Soumare, A.; Naguez, A.; Label, R.; Bernard, O.; Bruhat, A.; Fafournoux, P.; Tremblais, G.; Marchant, D.; Gille, T.; et al. HIF-1α triggers ER stress and CHOP-mediated apoptosis in alveolar epithelial cells, a key event in pulmonary fibrosis. Sci. Rep. 2018, 8, 17939. [Google Scholar] [CrossRef] [PubMed]
- Belaidi, E.; Thomas, A.; Bourdier, G.; Moulin, S.; Lemarié, E.; Levy, P.; Pépin, J.-L.; Korichneva, I.; Godin-Ribuot, D.; Arnaud, C. Endoplasmic reticulum stress as a novel inducer of hypoxia inducible factor-1 activity: Its role in the susceptibility to myocardial ischemia-reperfusion induced by chronic intermittent hypoxia. Int. J. Cardiol. 2016, 210, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Xu, L.; Xie, H.; Ouyang, R.; Ke, Y.; Zhou, R.; Yung, W.-H. Attenuation of intermittent hypoxia-induced apoptosis and fibrosis in pulmonary tissues via suppression of ER stress activation. BMC Pulm. Med. 2020, 20, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tanjore, H.; Lawson, W.E.; Blackwell, T.S. Endoplasmic reticulum stress as a pro-fibrotic stimulus. Biochim. et Biophys. Acta (BBA)-Mol. Basis Dis. 2013, 1832, 940–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delbrel, E.; Uzunhan, Y.; Soumare, A.; Gille, T.; Marchant, D.; Planès, C.; Boncoeur, E. ER Stress is Involved in Epithelial-To-Mesenchymal Transition of Alveolar Epithelial Cells Exposed to a Hypoxic Microenvironment. Int. J. Mol. Sci. 2019, 20, 1299. [Google Scholar] [CrossRef] [Green Version]
- Szegezdi, E.; Logue, S.; Gorman, A.; Samali, A. Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep. 2006, 7, 880–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pun, M. Human model of obstructive sleep apnea: How incomplete is it? J. Appl. Physiol. 2020, 129, 47. [Google Scholar] [CrossRef] [PubMed]
- Tashiro, J.; Rubio, G.A.; Limper, A.H.; Williams, K.; Elliot, S.J.; Ninou, I.; Aidinis, V.; Tzouvelekis, A.; Glassberg, M.K. Exploring Animal Models That Resemble Idiopathic Pulmonary Fibrosis. Front. Med. 2017, 4, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, T.; Hoyne, G.F.; A Knight, D.; Fear, M.; E Mutsaers, S.; Prêle, C.M. The contribution of animal models to understanding the role of the immune system in human idiopathic pulmonary fibrosis. Clin. Transl. Immunol. 2020, 9, e1153. [Google Scholar] [CrossRef]
- Hansen, N.U.B.; Karsdal, M.A.; Brockbank, S.; Cruwys, S.; Rønnow, S.R.; Leeming, D.J. Tissue turnover of collagen type I, III and elastin is elevated in the PCLS model of IPF and can be restored back to vehicle levels using a phosphodiesterase inhibitor. Respir. Res. 2016, 17, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Mo, L.; Feng, X.; Huang, M.; Li, L. Using bioinformatics approach identifies key genes and pathways in idiopathic pulmonary fibrosis. Medicine 2020, 99, e22099. [Google Scholar] [CrossRef]
- Nishioka, Y.; Azuma, M.; Kishi, M.; Aono, Y. Targeting platelet-derived growth factor as a therapeutic approach in pulmonary fibrosis. J. Med. Investig. 2013, 60, 175–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, R.J.; Burdick, M.D.; Hong, K.; Lutz, M.A.; Murray, L.A.; Xue, Y.Y.; Belperio, J.A.; Keane, M.P.; Strieter, R.M. Circulating fibrocytes traffic to the lungs in response to CXCL12 and mediate fibrosis. J. Clin. Investig. 2004, 114, 438–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.-S.; Liu, C.-C.; Lin, J.-H.; Hsu, T.-W.; Hsu, J.-W.; Su, K.; Hung, S.-C. Involvement of ER stress, PI3K/AKT activation, and lung fibroblast proliferation in bleomycin-induced pulmonary fibrosis. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Jiang, C.; Liu, G.; Luckhardt, T.; Antony, V.; Zhou, Y.; Carter, A.B.; Thannickal, V.J.; Liu, R.-M. Serpine 1 induces alveolar type II cell senescence through activating p53-p21-Rb pathway in fibrotic lung disease. Aging Cell 2017, 16, 1114–1124. [Google Scholar] [CrossRef]
- Rabieian, R.; Boshtam, M.; Zareei, M.; Kouhpayeh, S.; Masoudifar, A.; Mirzaei, H. Plasminogen Activator Inhibitor Type-1 as a Regulator of Fibrosis. J. Cell. Biochem. 2018, 119, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Rana, T.; Jiang, C.; Liu, G.; Miyata, T.; Antony, V.; Thannickal, V.J.; Liu, R.-M. PAI-1 Regulation of TGF-β1–induced Alveolar Type II Cell Senescence, SASP Secretion, and SASP-mediated Activation of Alveolar Macrophages. Am. J. Respir. Cell Mol. Biol. 2020, 62, 319–330. [Google Scholar] [CrossRef]
- Liu, R.-M.; Liu, G. Cell senescence and fibrotic lung diseases. Exp. Gerontol. 2020, 132, 110836. [Google Scholar] [CrossRef]
- Sarcinelli, C.; Dragic, H.; Piecyk, M.; Barbet, V.; Duret, C.; Barthelaix, A.; Ferraro-Peyret, C.; Fauvre, J.; Renno, T.; Chaveroux, C.; et al. ATF4-Dependent NRF2 Transcriptional Regulation Promotes Antioxidant Protection during Endoplasmic Reticulum Stress. Cancers 2020, 12, 569. [Google Scholar] [CrossRef] [Green Version]
- Tuleta, I.; Pizarro, C.; Nickenig, G.; Juergens, U.; Skowasch, D. Deleterious effects of intermittent hypoxia on lung tissue. Eur. Respir. J. 2016, 48, PA2070. [Google Scholar] [CrossRef]
- Kang, H.H.; Kim, I.K.; Yeo, C.D.; Kim, S.W.; Lee, H.Y.; Im, J.H.; Kwon, H.Y.; Lee, S.H. The Effects of Chronic Intermittent Hypoxia in Bleomycin-Induced Lung Injury on Pulmonary Fibrosis via Regulating the NF-κB/Nrf2 Signaling Pathway. Tuberc. Respir. Dis. 2020, 83, S63–S74. [Google Scholar] [CrossRef] [PubMed]
- Burman, A.; Kropski, J.A.; Calvi, C.L.; Serezani, A.P.; Pascoalino, B.D.; Han, W.; Sherrill, T.; Gleaves, L.; Lawson, W.E.; Young, L.R.; et al. Localized hypoxia links ER stress to lung fibrosis through induction of C/EBP homologous protein. JCI Insight 2018, 3, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Kim, Y.; Lim, M.J.; Park, Y.-G.; Park, S.I.; Sohn, J. The p38-activated ER stress-ATF6α axis mediates cellular senescence. FASEB J. 2019, 33, 2422–2434. [Google Scholar] [CrossRef] [Green Version]
- Polonis, K.; Becari, C.; Chahal, C.A.A.; Zhang, Y.; Allen, A.M.; Kellogg, T.A.; Somers, V.K.; Singh, P. Chronic Intermittent Hypoxia Triggers a Senescence-like Phenotype in Human White Preadipocytes. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Dong, Y.; Li, W.-C.; Tang, B.-X.; Li, J.; Zang, Y. Roxithromycin attenuates bleomycin-induced pulmonary fibrosis by targeting senescent cells. Acta Pharmacol. Sin. 2021, 1–11. [Google Scholar] [CrossRef]
- Hagiwara, S.-I.; Ishii, Y.; Kitamura, S. Aerosolized Administration of N-Acetylcysteine Attenuates Lung Fibrosis Induced by Bleomycin in Mice. Am. J. Respir. Crit. Care Med. 2000, 162, 225–231. [Google Scholar] [CrossRef]
- Perrini, S.; Cignarelli, A.; Quaranta, V.N.; Falcone, V.A.; Kounaki, S.; Porro, S.; Ciavarella, A.; Ficarella, R.; Barbaro, M.; Genchi, V.A.; et al. Correction of intermittent hypoxia reduces inflammation in obese subjects with obstructive sleep apnea. JCI Insight 2017, 2, e94379. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| Collagen 1a1 | GTGGTGACAAGGGTGAGACA | GAGAACCAGGAGAACCAGGA |
| Collagen 3a1 | TACACCTGCTCCTGTGCTTC | CATTCCTCCCACTCCAGACT |
| Serpin 1 | GCACAACCCGACAGAGACAA | ATGAAGGCGTCTCTTCCCAC |
| Cxcl 12 | CCTTCAGATTGTTGCAAGGCTG | TCCTTTGGGCTGTTGTGCTT |
| Pdgf β | TCGCCTGCAAGTGTGAGACA | CCGAATGGTCACCCGAGCTT |
| 18S | GTA AGT GCG GGC CAT AAG CTT | AGT CAA GTT CGA CCG TCT TCT CA |
| Antibodies (anti-) | Type | Host | Dilution | Reference | Manufacturer |
|---|---|---|---|---|---|
| β-ACTIN | Primary antibodies | Mouse | 1/5000 | A5316 | Sigma |
| ATF6-N | Mouse | 1/1000 | Clone 1-7 MAB 6762 | Abnova | |
| ATF4 | Rabbit | 1/1000 | SC-200 (C-20) | Santa Cruz | |
| GRP 78 | Rabbit | 1/1000 | ADI-SPA-7680050 | Enzo | |
| GRP 75 | Rabbit | 1/1000 | D 175 9661L | Cell Signaling | |
| sXBP1 | Mouse | 1/1000 | SC-8015 (F-4) | Santa Cruz | |
| Anti-mouse IgG HRP | Secondary antibodies | Goat | 1/3000 | P0447 | Dako |
| Anti-rabbit IgG HRP | Goat | 1/3000 | P0448 | Dako |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haine, L.; Bravais, J.; Yegen, C.-H.; Bernaudin, J.-F.; Marchant, D.; Planès, C.; Voituron, N.; Boncoeur, E. Sleep Apnea in Idiopathic Pulmonary Fibrosis: A Molecular Investigation in an Experimental Model of Fibrosis and Intermittent Hypoxia. Life 2021, 11, 973. https://doi.org/10.3390/life11090973
Haine L, Bravais J, Yegen C-H, Bernaudin J-F, Marchant D, Planès C, Voituron N, Boncoeur E. Sleep Apnea in Idiopathic Pulmonary Fibrosis: A Molecular Investigation in an Experimental Model of Fibrosis and Intermittent Hypoxia. Life. 2021; 11(9):973. https://doi.org/10.3390/life11090973
Chicago/Turabian StyleHaine, Liasmine, Juliette Bravais, Céline-Hivda Yegen, Jean-Francois Bernaudin, Dominique Marchant, Carole Planès, Nicolas Voituron, and Emilie Boncoeur. 2021. "Sleep Apnea in Idiopathic Pulmonary Fibrosis: A Molecular Investigation in an Experimental Model of Fibrosis and Intermittent Hypoxia" Life 11, no. 9: 973. https://doi.org/10.3390/life11090973