
Continuous Feeding Reduces the Generation of Metabolic Byproducts and Increases Antibodies Expression in Chinese Hamster Ovary-K1 Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Media
2.2. Process Conditions
2.3. Cells, Metabolites and Osmolality Analysis
2.4. Antibody Analysis by HPLC
2.5. Physicochemical Analysis
2.6. Statistical Analysis
3. Results and Discussion
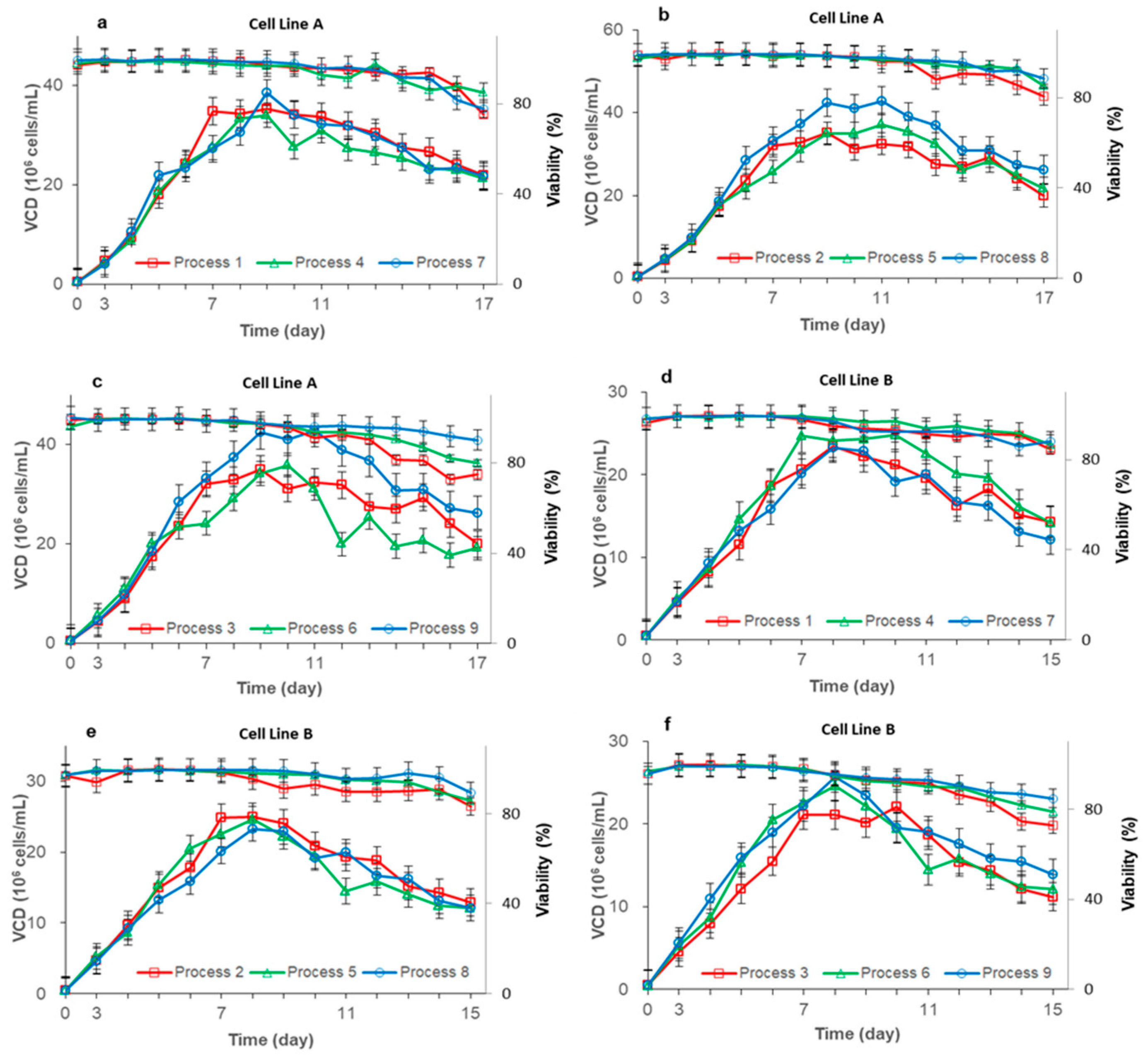
3.1. Effects of Three Different Feeding Processes on CHO Cell Growth
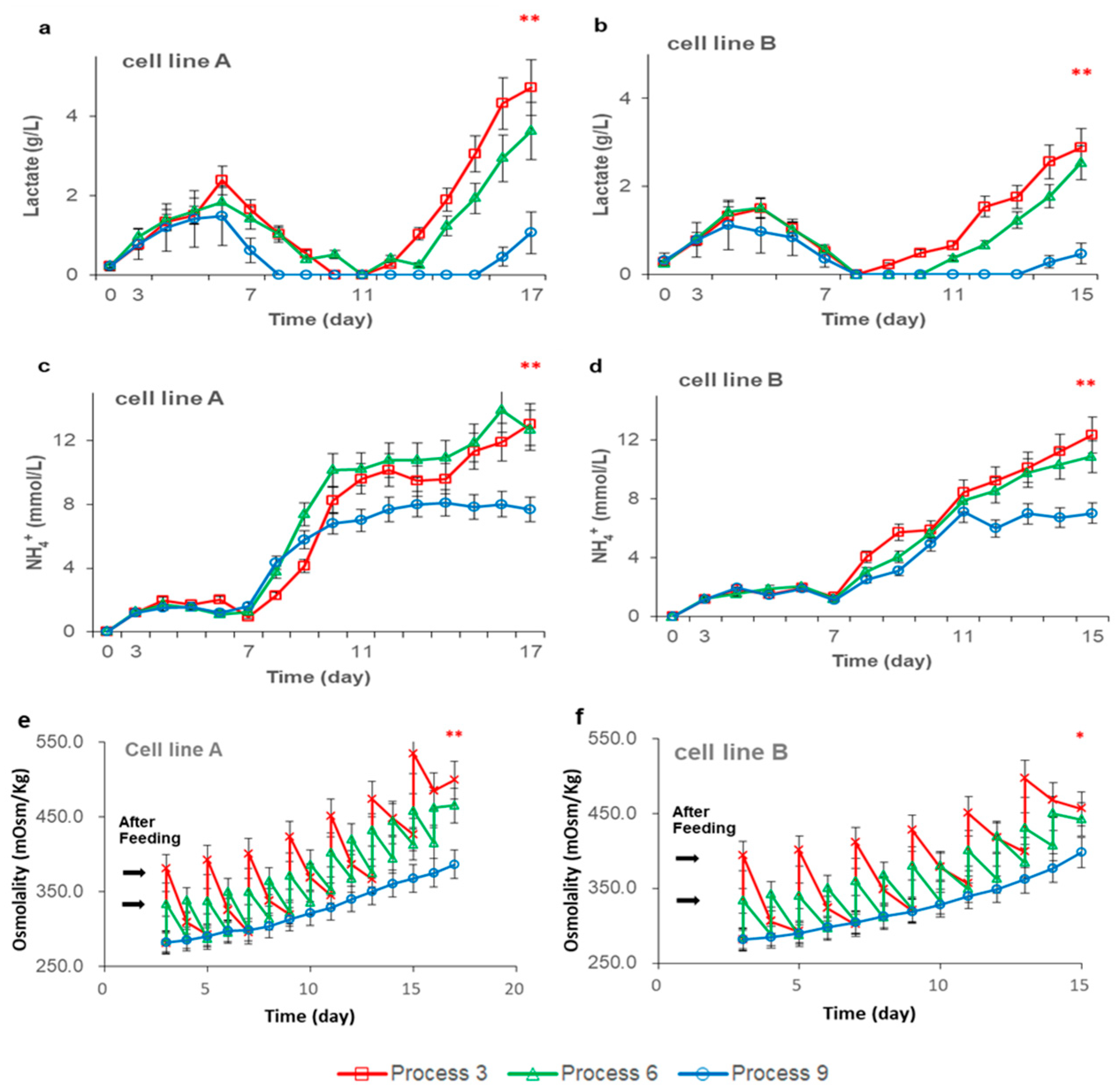
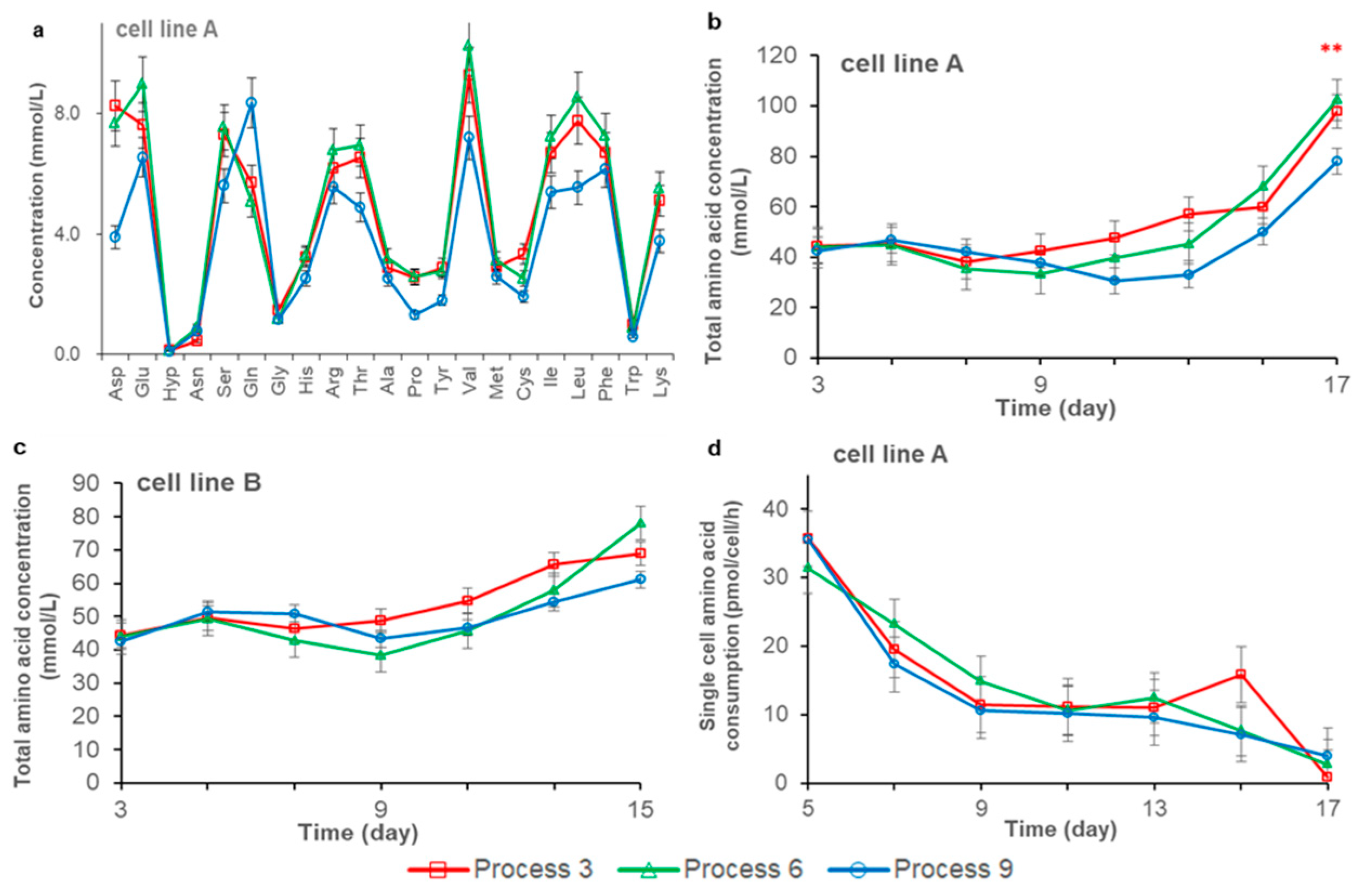
3.2. Effects of Feeding Strategies on Cell Metabolism
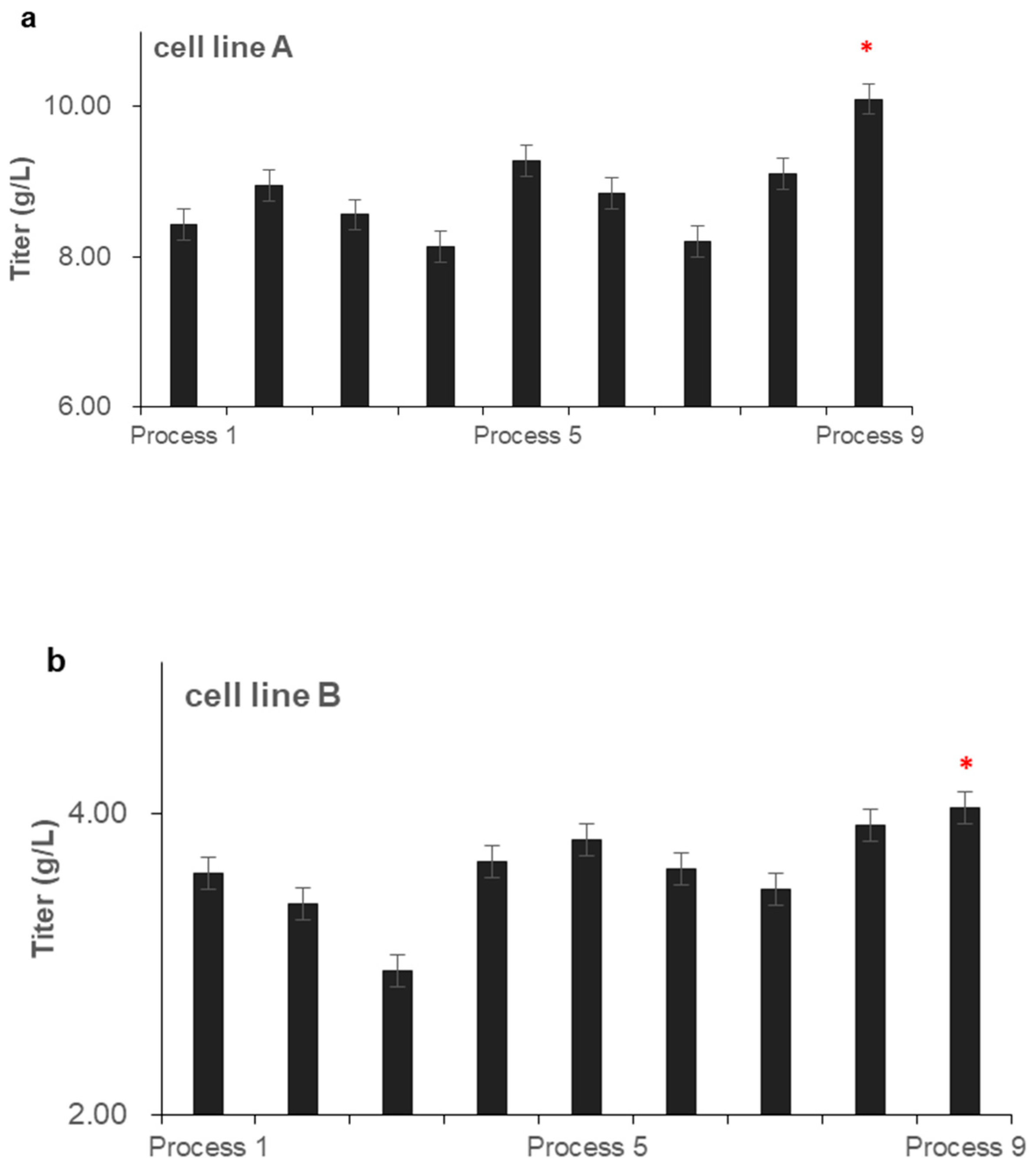
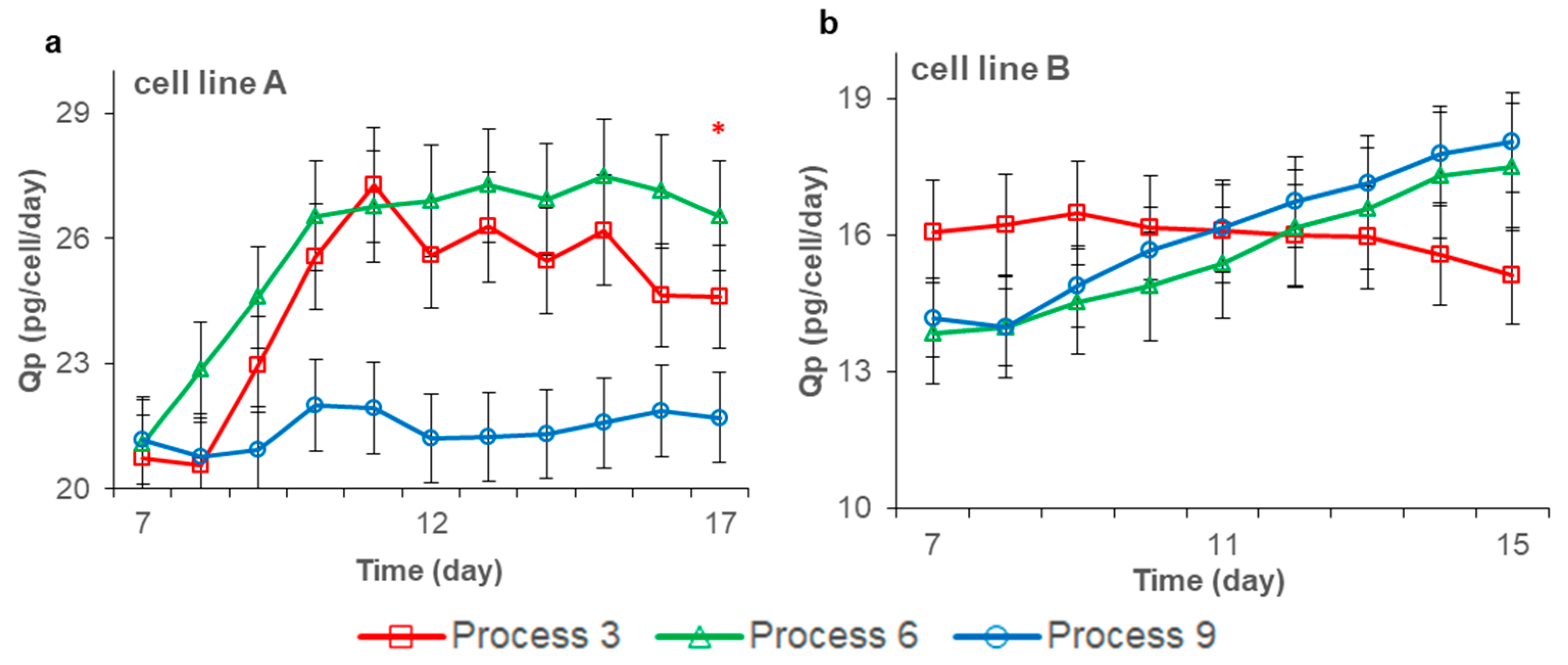
3.3. Effects of Feeding Strategies on Antibody Expression and Quality
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Urquhart, L. Market watch: Top drugs and companies by sales in 2017. Nat. Rev. Drug Discov. 2018, 17, 232. [Google Scholar] [CrossRef]
- Ecker, D.M.; Jones, S.D.; Levine, H.L. The therapeutic monoclonal antibody market. mAbs 2015, 7, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omasa, T.; Onitsuka, M.; Kim, W.D. Cell Engineering and Cultivation of Chinese Hamster Ovary (CHO) Cells. Curr. Pharm. Biotechnol. 2010, 11, 233–240. [Google Scholar] [CrossRef]
- Fischer, S.; Handrick, R.; Otte, K. The art of CHO cell engineering: A comprehensive retrospect and future perspectives. Biotechnol. Adv. 2015, 33, 1878–1896. [Google Scholar] [CrossRef]
- He, Y.; Wang, K.; Yan, N. The recombinant expression systems for structure determination of eukaryotic membrane proteins. Protein Cell 2014, 5, 658–672. [Google Scholar] [CrossRef] [Green Version]
- Kunert, R.; Reinhart, D. Advances in recombinant antibody manufacturing. Appl. Microbiol. Biotechnol. 2016, 100, 3451–3461. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Vijayasankaran, N.; Shen, A.Y.; Kiss, R.; Amanullah, A. Cell culture processes for monoclonal antibody production. mAbs 2010, 2, 466–479. [Google Scholar] [CrossRef] [Green Version]
- Brunner, M.; Doppler, P.; Klein, T.; Herwig, C.; Fricke, J. Elevated pCO2 affects the lactate metabolic shift in CHO cell culture processes. Eng. Life Sci. 2018, 18, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Val, I.J.; Muller, C.; Sen, J.W.; Rasmussen, S.K.; Kontoravdi, C.; Andersen, M.R. Amino acid and glucose metabolism in fed-batch CHO cell culture affects antibody production and glycosylation. Biotechnol. Bioeng. 2015, 112, 521–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, M.; Altamirano, C.; Dickson, A.J. Process and metabolic engineering perspectives of lactate production in mammalian cell cultures. Curr. Opin. Chem. Eng. 2018, 22, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Castan, A.; Stevenson, J.; Chatzissavidou, N.; Vilaplana, F.; Chotteau, V. Combined effects of glycosylation precursors and lactate on the glycoprofile of IgG produced by CHO cells. J. Biotechnol. 2019, 289, 71–79. [Google Scholar] [CrossRef]
- Konakovsky, V.; Clemens, C.; Müller, M.M.; Bechmann, J.; Berger, M.; Schlatter, S.; Herwig, C. Metabolic Control in Mammalian Fed-Batch Cell Cultures for Reduced Lactic Acid Accumulation and Improved Process Robustness. Bioengineering 2016, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Karengera, E.; Robotham, A.; Kelly, J.; Durocher, Y.; De, C.G.; Henry, O. Concomitant reduction of lactate and ammonia accumulation in fed-batch cultures: Impact on glycoprotein production and quality. Biotechnol. Prog. 2018, 34, 494–504. [Google Scholar] [CrossRef]
- Kuwae, S.; Miyakawa, I.; Doi, T. Development of a chemically defined platform fed-batch culture media for monoclonal antibody-producing CHO cell lines with optimized choline content. Cytotechnology 2018, 70, 939–948. [Google Scholar] [CrossRef]
- Pan, X.; Streefland, M.; Dalm, C.; Wijffels, R.H.; Martens, D.E. Selection of chemically defined media for CHO cell fed-batch culture processes. Cytotechnology 2017, 69, 39–56. [Google Scholar] [CrossRef] [Green Version]
- Graham, R.J.; Bhatia, H.; Yoon, S. Consequences of trace metal variability and supplementation on Chinese hamster ovary (CHO) cell culture performance: A review of key mechanisms and considerations. Biotechnol. Bioeng. 2019, 116, 3446–3456. [Google Scholar] [CrossRef]
- Lu, F.; Toh, P.C.; Burnett, I.; Li, F.; Hudson, T.; Amanullah, A.; Li, J. Automated dynamic fed-batch process and media optimization for high productivity cell culture process development. Biotechnol. Bioeng. 2013, 110, 191–205. [Google Scholar] [CrossRef]
- Hu, S.; Deng, L.; Wang, H.; Zhuang, Y.; Chu, J.; Zhang, S.; Li, Z.; Guo, M. Bioprocess development for the production of mouse-human chimeric anti-epidermal growth factor receptor vIII antibody C12 by suspension culture of recombinant Chinese hamster ovary cells. Cytotechnology 2011, 63, 247–258. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Harcum, S.W. Effects of amino acid additions on ammonium stressed cho cells. J. Biotechnol. 2005, 117, 277–286. [Google Scholar] [CrossRef]
- Romanova, N.; Niemann, T.; Greiner, J.F.W.; Kaltschmidt, B.; Kaltschmidt, C.; Noll, T. Hyperosmolality in CHO Culture: Effects on cellular behavior and morphology. Biotechnol. Bioeng. 2021, 118, 2348–2359. [Google Scholar] [CrossRef]
- Ahn, W.S.; Antoniewicz, M.R. Metabolic flux analysis of CHO cells at growth and non-growth phases using isotopic tracers and mass spectrometry. Metab. Eng. 2011, 13, 598–609. [Google Scholar] [CrossRef]
- Li, J.; Wong, C.L.; Vijayasankaran, N.; Hudson, T.; Amanullah, A. Feeding lactate for cho cell culture processes: Impact on culture metabolism and performance. Biotechnol. Bioeng. 2012, 109, 1173–1186. [Google Scholar] [CrossRef]
- Hassell, T.; Gleave, S.; Butler, M. Growth Inhibition in Animal Cell Culture: The Effect of Lactate and Ammonia. Appl. Biochem. Biotechnol. 1991, 30, 29–41. [Google Scholar] [CrossRef]
- Dean, J.; Reddy, P. Metabolic analysis of antibody producing CHO cells in fed-batch production. Biotechnol. Bioeng. 2013, 110, 1735–1747. [Google Scholar] [CrossRef]
- Xu, S.; Hoshan, L.; Chen, H. Improving lactate metabolism in an intensified CHO culture process: Productivity and product quality considerations. Bioprocess. Biosyst. Eng. 2016, 39, 1689–1702. [Google Scholar] [CrossRef]
- Zhu, M.M.; Goyal, A.; Rank, D.L.; Gupta, S.K.; Boom, V.T.; Lee, S.S. Effects of elevated pCO2 and osmolality on growth of CHO cells and production of antibody-fusion protein B1: A case study. Biotechnol. Prog. 2005, 21, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Zagari, F. Multidisciplinary Analysis of the Metabolic Shift to Lactate Consumption in CHO Cell Culture; EPFL: Lausanne, Switzerland, 2012. [Google Scholar] [CrossRef]
- Alhuthali, S.; Kotidis, P.; Kontoravdi, C. Osmolality Effects on CHO Cell Growth, Cell Volume, Antibody Productivity and Glycosylation. Int. J. Mol. Sci. 2021, 22, 3290. [Google Scholar] [CrossRef]
- Du, Z.; Treiber, D.; McCarter, J.D.; Fomina-Yadlin, D.; Saleem, R.A.; McCoy, R.E.; Zhang, Y.; Tharmalingam, T.; Leith, M.; Follstad, B.D.; et al. Use of a small molecule cell cycle inhibitor to control cell growth and improve specific productivity and product quality of recombinant proteins in CHO cell cultures. Biotechnol. Bioeng. 2015, 112, 141–155. [Google Scholar] [CrossRef] [Green Version]
- Pacis, E.; Yu, M.; Autsen, J.; Bayer, R.; Li, F. Effects of cell culture conditions on antibody N-linked glycosylation—What affects high mannose 5 glycoform. Biotechnol. Bioeng. 2011, 108, 2348–2358. [Google Scholar] [CrossRef]
- Slade, P.G.; Caspary, R.G.; Nargund, S.; Huang, C.J. Mannose metabolism in recombinant CHO cells and its effect on IgG glycosylation. Biotechnol. Bioeng. 2016, 113, 1468–1480. [Google Scholar] [CrossRef]
- Shi, H.H.; Goudar, C.T. Recent advances in the understanding of biological implications and modulation methodologies of monoclonal antibody N-linked high mannose glycans. Biotechnol. Bioeng. 2014, 111, 1907–1919. [Google Scholar] [CrossRef]
- Boswell, C.A.; Tesar, D.B.; Mukhyala, K.; Theil, F.P.; Khawli, L.A. Effects of charge on antibody tissue distribution and pharmacokinetics. Bioconjug. Chem. 2010, 21, 2153–2163. [Google Scholar] [CrossRef]
- Zheng, Y.; Tesar, D.B.; Benincosa, L.; Birnböck, H.; Boswell, C.A.; Bumbaca, D.; Cowan, K.J.; Danilenko, D.M.; Daugherty, A.L.; Fielder, P.J.; et al. Minipig as a potential translatable model for monoclonal antibody pharmacokinetics after intravenous and subcutaneous administration. MAbs 2012, 4, 243–255. [Google Scholar] [CrossRef] [Green Version]
- Visser, J.; Feuerstein, I.; Stangler, T.; Schmiederer, T.; Schiestl, M. Physicochemical and functional comparability between the proposed biosimilar rituximab GP2013 and originator rituximab. BioDrugs 2013, 27, 495–507. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.M.; Hu, W.; Rustandi, E.; Chang, K.; Yusuf-Makagiansar, H.; Ryll, T. Maximizing productivity of CHO cell-based fed-batch culture using chemically defined media conditions and typical manufacturing equipment. Biotechnol. Prog. 2010, 26, 1400–1410. [Google Scholar] [CrossRef]
- Liste-Calleja, L.; Lecina, M.; Lopez-Repullo, J.; Albiol, J.; Solà, C.; Cairó, J.J. Lactate and glucose concomitant consumption as a self-regulated pH detoxification mechanism in HEK293 cell cultures. Appl. Microbiol. Biotechnol. 2015, 99, 9951–9960. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process | Feed Methods | Total CB7a Feeding Amount (V/V%) ① | Feeding Strategy | ||
|---|---|---|---|---|---|
| Cell Line A | Cell Line B | Cell Line A | Cell Line B | ||
| Process 1 | Once every two days | 28 | 24 | 120 mL each time from Day 3 | |
| Process 2 | Once every two days | 35 | 30 | 150 mL each time from Day 3 | |
| Process 3 | Once every two days | 42 | 36 | 180 mL each time from Day 3 | |
| Process 4 | Daily feeding | 28 | 24 | 60 mL each time from Day 3 | |
| Process 5 | Daily feeding | 35 | 30 | 75 mL each time from Day 3 | |
| Process 6 | Daily feeding | 42 | 36 | 90 mL each time from Day 3 | |
| Process 7 | Continuous feeding ② | 28 | 24 | Starting from the third day, the on-time of the pump was set to 2S; the off-time was set to 738S | |
| Process 8 | Continuous feeding | 35 | 30 | Starting from the third day, the on-time of the pump was set to 2S; the off-time was set to 590S | |
| Process 9 | Continuous feeding | 42 | 36 | Starting from the third day, the on-time of the pump was set to 2S; the off-time was set to 491S | |
| Quality | Process 3 | Process 6 | Process 9 | |
|---|---|---|---|---|
| Cell line A | SEC purity (%) | 98.79 ± 0.43 | 99.05 ± 0.48 | 98.88 ± 0.54 |
| CEX AV (%) | 24.60 ± 3.21 | 24.75 ± 2.88 | 22.70 ± 3.01 | |
| CEX MP (%) | 66.08 ± 5.25 | 67.16 ± 4.72 | 68.23 ± 3.76 | |
| CEX BV (%) | 9.32 ± 2.23 | 8.09 ± 1.85 | 9.06 ± 1.54 | |
| G2F (%) | 0.27 ± 0.15 | 0.13 ± 0.13 | 0.33 ± 0.18 | |
| G1F (%) | 7.80 ± 1.26 | 8.30 ± 1.83 | 7.53 ± 1.37 | |
| G0F (%) | 86.63 ± 9.19 | 88.17 ± 8.38 | 89.37 ± 9.02 | |
| High-mannose (%) | 2.15 ± 0.47 | 1.39 ± 0.43 | 0.88 ± 0.24 | |
| NR-CE (%) | 96.12 ± 1.13 | 96.85 ± 1.56 | 96.52 ± 1.45 |
| Quality | Process 3 | Process 6 | Process 9 | |
|---|---|---|---|---|
| Cell line B | SEC purity (%) | 97.75 ± 0.54 | 97.45 ± 0.50 | 98.05 ± 0.59 |
| CEX AV (%) | 18.24 ± 2.34 | 19.23 ± 2.07 | 18.01 ± 1.34 | |
| CEX MP (%) | 72.55 ± 4.54 | 72.01 ± 3.54 | 72.26 ± 3.29 | |
| CEX BV (%) | 9.21 ± 1.47 | 8.76 ± 1.09 | 9.73 ± 1.21 | |
| G2F (%) | 2.17 ± 1.38 | 1.89 ± 1.53 | 2.23 ± 1.77 | |
| G1F (%) | 11.20 ± 1.88 | 13.15 ± 2.01 | 12.45 ± 2.34 | |
| G0F (%) | 78.28 ± 8.78 | 77.58 ± 7.56 | 80.30 ± 9.57 | |
| High-mannose (%) | 4.35 ± 1.43 | 3.38 ± 0.76 | 1.02 ± 0.35 | |
| NR-CE (%) | 95.64 ± 1.02 | 95.43 ± 0.87 | 95.78 ± 0.93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, S.; Ahmed, W.; Mohsin, A.; Guo, M. Continuous Feeding Reduces the Generation of Metabolic Byproducts and Increases Antibodies Expression in Chinese Hamster Ovary-K1 Cells. Life 2021, 11, 945. https://doi.org/10.3390/life11090945
Xiao S, Ahmed W, Mohsin A, Guo M. Continuous Feeding Reduces the Generation of Metabolic Byproducts and Increases Antibodies Expression in Chinese Hamster Ovary-K1 Cells. Life. 2021; 11(9):945. https://doi.org/10.3390/life11090945
Chicago/Turabian StyleXiao, Shang, Waqas Ahmed, Ali Mohsin, and Meijin Guo. 2021. "Continuous Feeding Reduces the Generation of Metabolic Byproducts and Increases Antibodies Expression in Chinese Hamster Ovary-K1 Cells" Life 11, no. 9: 945. https://doi.org/10.3390/life11090945