
The Genetic Variants of NOTCH3 (6746T>C) and PSMA6 (-8C>G) as Possible Risk Factors of Psoriasis Development

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Groups
2.2. DNA Isolation and Genotyping of NOTCH3 6746T>C and PSMA6 -8C>G Polymorphisms
2.3. Statistical Methods
3. Results
3.1. Characteristics of the Patients
3.2. Frequencies of Genotypes and Alleles of NOTCH3 and PMSA6 Polymorphisms
3.3. NOTCH3 and PSMA6 Polymorphisms and Psoriasis in the Analysis of Genetic Models
3.4. Joint Analysis of NOTCH3 and PSMA6 Polymorphisms
3.5. Correlations between Analyzed Polymorphisms and Psoriasis Clinical Features
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dand, N.; Mahil, S.K.; Capon, F.; Smith, C.H.; Simpson, M.A.; Barker, J.N. Psoriasis and Genetics. Acta Derm. Venereol. 2020, 100, adv00030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nedoszytko, B.; Szczerkowska-Dobosz, A.; Stawczyk-Macieja, M.; Owczarczyk-Saczonek, A.; Reich, A.; Bartosińska, J.; Batycka-Baran, A.; Czajkowski, R.; Dobrucki, I.T.; Dobrucki, L.W.; et al. Pathogenesis of psoriasis in the “omic” era. Part II. Genetic, genomic and epigenetic changes in psoriasis. Postep. Dermatol. Alergol. 2020, 37, 283–298. [Google Scholar] [CrossRef]
- Lowes, M.A.; Russell, C.B.; Martin, D.A.; Towne, J.E.; Krueger, J.G. The IL-23/T17 pathogenic axis in psoriasis is amplified by keratinocyte responses. Trends Immunol. 2013, 34, 174–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nussbaum, L.; Chen, Y.L.; Ogg, G.S. Role of regulatory T cells in psoriasis pathogenesis and treatment. Br. J. Dermatol. 2021, 184, 14–24. [Google Scholar] [CrossRef]
- Karabowicz, P.; Wroński, A.; Ostrowska, H.; Waeg, G.; Zarkovic, N.; Skrzydlewska, E. Reduced Proteasome Activity and Enhanced Autophagy in Blood Cells of Psoriatic Patients. Int. J. Mol. Sci. 2020, 21, 7608. [Google Scholar] [CrossRef]
- Gratton, R.; Tricarico, P.M.; Moltrasio, C.; Lima Estevão de Oliveira, A.S.; Brandão, L.; Marzano, A.V.; Zupin, L.; Crovella, S. Pleiotropic Role of Notch Signaling in Human Skin Diseases. Int. J. Mol. Sci. 2020, 21, 4214. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Kong, Y.; Xu, M.; Zhang, H. Notch3 functions as a tumor suppressor by controlling cellular senescence. Cancer Res. 2013, 73, 3451–3459. [Google Scholar] [CrossRef] [Green Version]
- Rooney, P.; Connolly, M.; Gao, W.; McCormick, J.; Biniecka, M.; Sullivan, O.; Kirby, B.; Sweeney, C.; Molloy, E.; Markham, T.; et al. Notch-1 mediates endothelial cell activation and invasion in psoriasis. Exp. Dermatol. 2014, 23, 113–118. [Google Scholar] [CrossRef]
- Ota, T.; Takekoshi, S.; Takagi, T.; Kitatani, K.; Toriumi, K.; Kojima, T.; Kato, M.; Ikoma, N.; Mabuchi, T.; Ozawa, A. Notch signaling may be involved in the abnormal differentiation of epidermal keratinocytes in psoriasis. Acta Histochem. Cytochem. 2014, 47, 175–183. [Google Scholar] [CrossRef] [Green Version]
- Nickoloff, B.J.; Qin, J.Z.; Chaturvedi, V.; Denning, M.F.; Bonish, B.; Miele, L. Jagged-1 mediated activation of notch signaling induces complete maturation of human keratinocytes through NF-kappaB and PPARgamma. Cell Death Differ. 2002, 9, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Mitra, A.; Shanthalingam, S.; Sherman, H.L.; Singh, K.; Canakci, M.; Torres, J.A.; Lawlor, R.; Ran, Y.; Golde, T.E.; Miele, L.; et al. CD28 Signaling Drives Notch Ligand Expression on CD4 T Cells. Front. Immunol. 2020, 11, 735. [Google Scholar] [CrossRef]
- Lehar, S.M.; Bevan, M.J. Immunology: Polarizing a T-cell response. Nature 2004, 430, 150–151. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.; Huang, Y.; Zhu, X.; Lin, X.; Luo, D. miR-125b-mediated regulation of cell proliferation through the Jagged-1/Notch signaling pathway by inhibiting BRD4 expression in psoriasis. Mol. Med. Rep. 2019, 19, 5227–5236. [Google Scholar] [CrossRef] [Green Version]
- Caporarello, N.; Lupo, G.; Olivieri, M.; Cristaldi, M.; Cambria, M.T.; Salmeri, M.; Anfuso, C.D. Classical VEGF, Notch and Ang signaling in cancer angiogenesis, alternative approaches and future directions (Review). Mol. Med. Rep. 2017, 16, 4393–4402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, H.E.; Neves, K.B.; Montezano, A.C.; MacLean, M.R.; Touyz, R.M. Notch3 signaling and vascular remodeling in pulmonary arterial hypertension. Clin. Sci. 2019, 133, 2481–2498. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Xue, H.; Gao, T.; Gao, M.; Zhang, Y. Notch1 Signaling Regulates the Th17/Treg Immune Imbalance in Patients with Psoriasis Vulgaris. Mediat. Inflamm. 2018, 2018, 3069521. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, S.; Natsuizaka, M.; Yashiro-Ohtani, Y.; Kalman, R.A.; Nakagawa, M.; Wu, L.; Klein-Szanto, A.J.; Herlyn, M.; Diehl, J.A.; Katz, J.P.; et al. NOTCH1 and NOTCH3 coordinate esophageal squamous differentiation through a CSL-dependent transcriptional network. Gastroenterology 2010, 139, 2113–2123. [Google Scholar] [CrossRef] [Green Version]
- Williams, S.E.; Beronja, S.; Pasolli, H.A.; Fuchs, E. Asymmetric cell divisions promote Notch-dependent epidermal differentiation. Nature 2011, 470, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-López, S.; Monsalve, E.M.; Romero de Ávila, M.J.; González-Gómez, J.; Hernández de León, N.; Ruiz-Marcos, F.; Baladrón, V.; Nueda, M.L.; García-León, M.J.; Screpanti, I.; et al. NOTCH3 signaling is essential for NF- ĸB activation in TLR-activated macrophages. Sci. Rep. 2020, 10, 14839. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, Y.; Tsukumo, S.; Chiba, S.; Hirai, H.; Hayashi, Y.; Okada, H.; Kishihara, K.; Yasutomo, K. Delta1-Notch3 interactions bias the functional differentiation of activated CD4+ T cells. Immunity 2003, 19, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Chitra, S.; Nalini, G.; Rajasekhar, G. The ubiquitin proteasome system and efficacy of proteasome inhibitors in diseases. Int. J. Rheum. Dis. 2012, 15, 249–260. [Google Scholar] [CrossRef]
- Pleńkowska, J.; Gabig-Cimińska, M.; Mozolewski, P. Oxidative Stress as an Important Contributor to the Pathogenesis of Psoriasis. Int. J. Mol. Sci. 2020, 21, 6206. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, K.; Sato, H.; Iida, A.; Mizuno, H.; Nakamura, T.; Miyamoto, Y.; Takahashi, A.; Tsunoda, T.; Ikegawa, S.; Kamatani, N.; et al. A functional SNP in PSMA6 confers risk of myocardial infarction in the Japanese population. Nat. Genet. 2006, 38, 921–925. [Google Scholar] [CrossRef]
- Ogawa, E.; Sato, Y.; Minagawa, A.; Okuyama, R. Pathogenesis of psoriasis and development of treatment. J. Dermatol. 2018, 45, 264–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuart, P.E.; Nair, R.P.; Ellinghaus, E.; Ding, J.; Tejasvi, T.; Gudjonsson, J.E.; Li, Y.; Weidinger, S.; Eberlein, B.; Gieger, C.; et al. Genome-wide association analysis identifies three psoriasis susceptibility loci. Nat. Genet. 2010, 42, 1000–1004. [Google Scholar] [CrossRef] [PubMed]
- Comegna, I.; Sainz, B., Jr.; Citera, G.; Maldonado-Cocco, J.A.; Garry, R.F.; Espinoza, L.R. Anti-20S proteasome antibodies in psoriatic arthritis. J. Rheumatol. 2008, 35, 674–676. [Google Scholar]
- Bachmann, H.S.; Novotny, J.; Sixt, S.; Liebisch, P.; Frey, U.H.; Duhrsen, U.; Siffert, W.; Nuckel, H. The G-Allele of the PSMA6-8C>G polymorphism is associated with poor outcome in multiple myeloma independently of circulating proteasome serum levels. Eur. J. Haematol. 2010, 85, 108–113. [Google Scholar] [CrossRef]
- Buraczynska, M.; Stec, A.; Filipczak, A.; Książek, A. Association between functional variant of inflammatory system gene (PSMA6) and end-stage kidney disease. Int. Urol. Nephrol. 2016, 48, 2083–2087. [Google Scholar] [CrossRef] [Green Version]
- Zmorzyński, S.; Wojcierowska-Litwin, M.; Kowal, M.; Michalska-Jakubus, M.; Styk, W.; Filip, A.A.; Walecka, I.; Krasowska, D. NOTCH3 T6746C and TP53 P72R Polymorphisms Are Associated with the Susceptibility to Diffuse Cutaneous Systemic Sclerosis. Biomed. Res. Int. 2020, 2020, 8465971. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
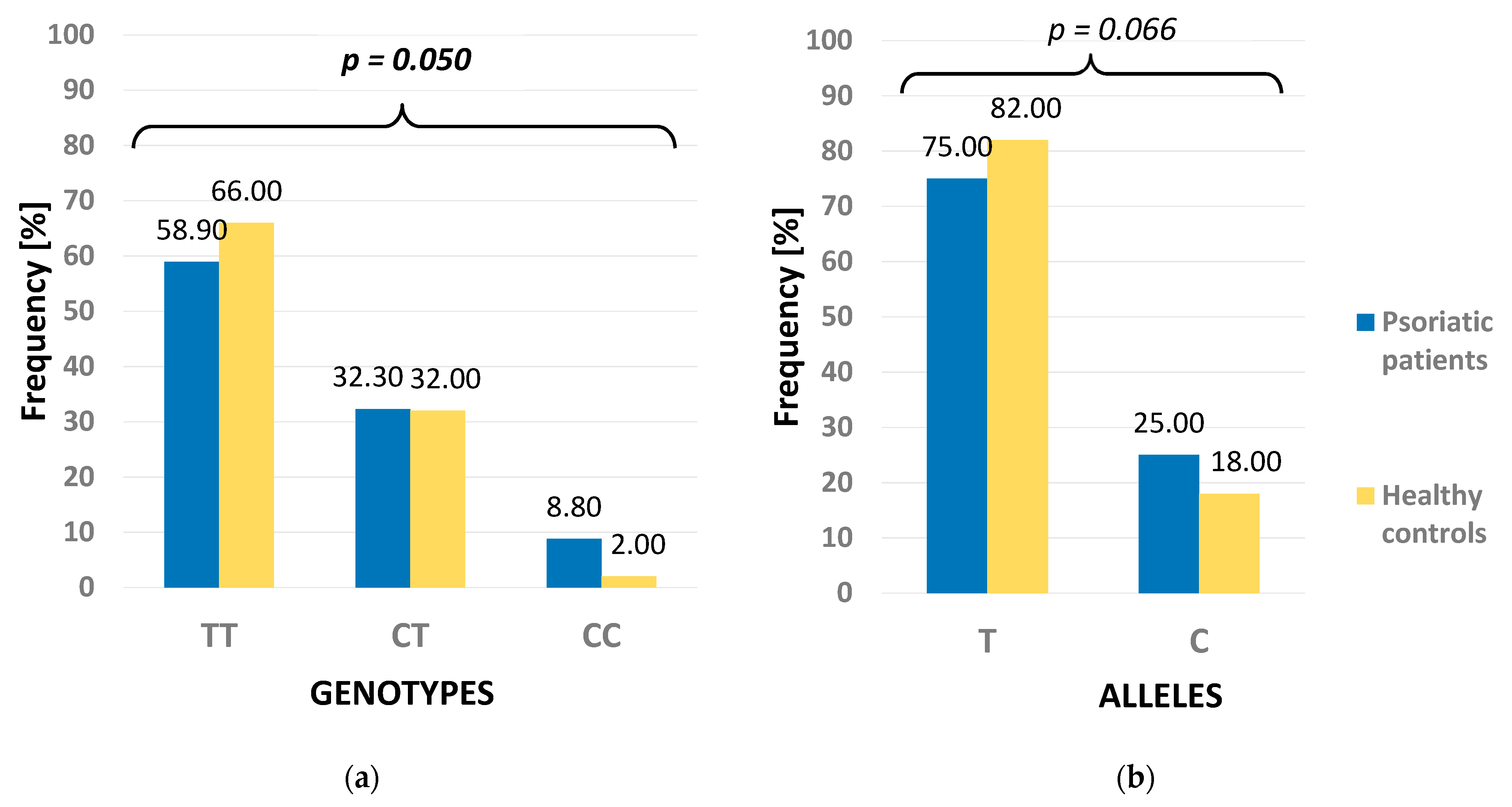{kind=link}
{kind=link}
| Variable, Parameter | Unit or Category | Psoriatic Patients (n = 158) | Healthy Controls (n = 100) | p |
|---|---|---|---|---|
| Age, Min–Max, M ± SD | years | 20–84, 46.5 ± 14.1 | 18–61, 34.3 ± 11.4 | <0.001 |
| Gender, n (%) | men | 112 (70.89) | 50 (50.00) | 0.001 |
| women | 46 (29.11) | 50 (50.00) | ||
| Age of psoriasis onset, Min–Max, M ± SD | years | 1–63, 23.3 ± 12.3 | ||
| Psoriasis subtype, n (%) | at/under 40 years of age | 137 (86.71) | ||
| over 40 years of age | 21 (13.29) | |||
| Psoriasis duration, Min–Max, M ± SD | years | 2–52, 23.3 ± 12.5 | ||
| Psoriatic arthritis, n (%) | yes | 56 (35.44) | ||
| Severity of psoriasis, n (%) | mild | 40 (25.32) | ||
| moderate or severe | 118 (74.68) | |||
| Positive family history, n (%) (n = 135) | yes | 67 (49.63) |
| Polymorphism | Genetic Model | Genotypes/Alleles | Psoriatic Patients (n = 158) n (%) | Healthy Controls (n = 100) n (%) | p |
|---|---|---|---|---|---|
| NOTCH3 | Dominant | TC + CC | 65 (41.14) | 34 (34.00) | 0.293 |
| TT | 93 (58.86) | 66 (66.00) | |||
| Recessive | CC | 14 (8.86) | 2 (2.00) | 0.032 | |
| TT + TC | 144 (91.14) | 98 (98.00) | |||
| Additive | CC | 14 (13.08) | 2 (2.94) | 0.030 | |
| TT | 93 (86.92) | 66 (97.06) | |||
| Heterozygote | TC | 51 (35.42) | 32 (32.65) | 0.681 | |
| TT | 93 (64.58) | 66 (66.35) | |||
| Allelic | C | 79 (25.00) | 36 (18.00) | 0.066 | |
| T | 237 (75.00) | 164 (82.00) | |||
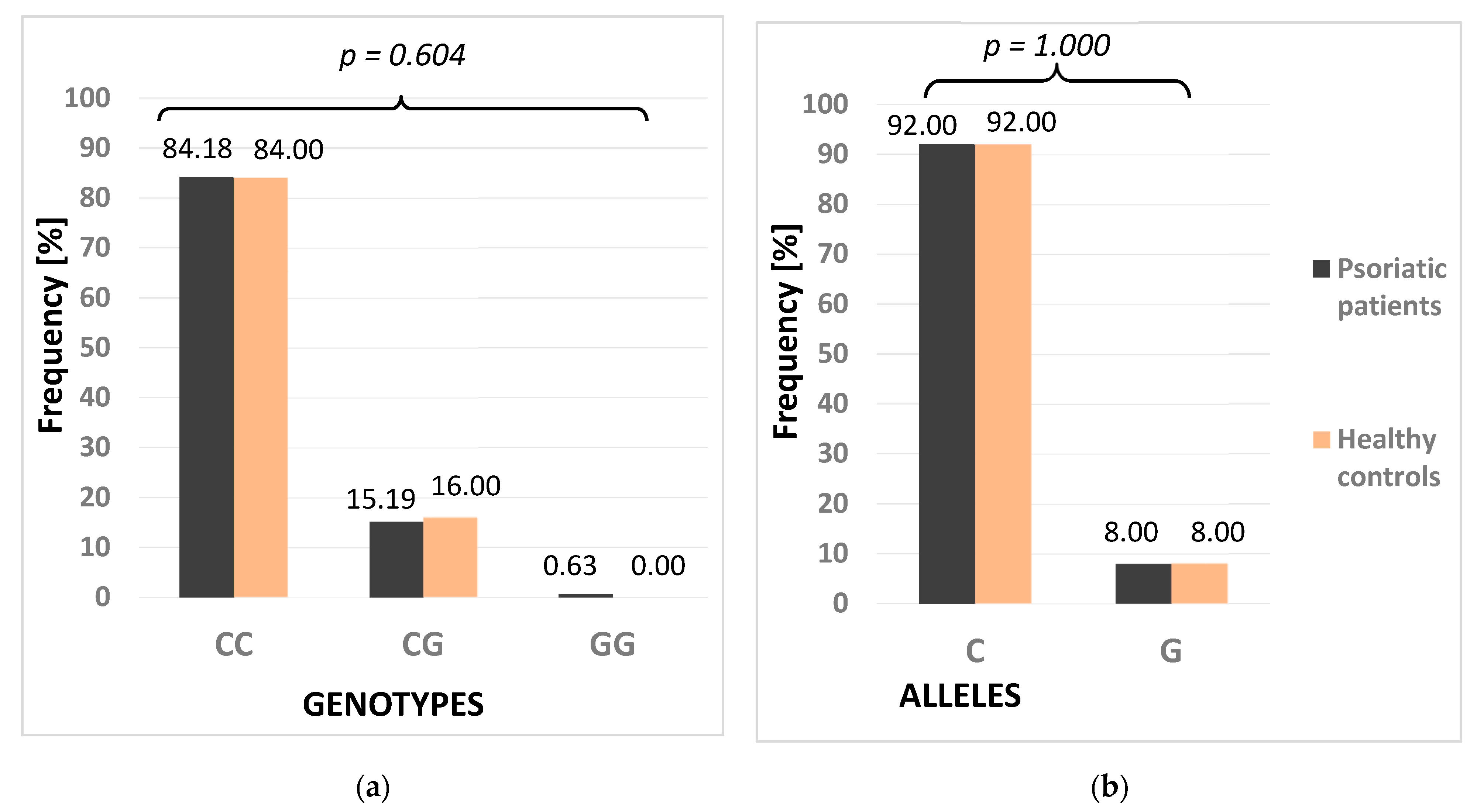
| PSMA6 | Dominant | CG + GG | 25 (15.82) | 16 (16.00) | 1.000 |
| CC | 133 (84.18) | 84 (84.00) | |||
| Recessive | GG | 1 (0.63) | 0 (0.00) | 1.000 | |
| CC + CG | 157 (99.37) | 100 (100.00) | |||
| Additive | GG | 1 (0.75) | 0 (0.00) | 1.000 | |
| CC | 133 (99.25) | 84 (84.00) | |||
| Heterozygote | CG | 24 (15.29) | 16 (16.00) | 1.000 | |
| CC | 133 (84.71) | 84 (84.00) | |||
| Allelic | G | 25 (8.00) | 16 (8.00) | 1.000 | |
| C | 291 (92.00) | 184 (92.00) |
| NOTCH3 Genotypes | PSMA6 Genotypes | Psoriatic Patients (n = 158) n (%) | Healthy Controls (n = 100) n (%) |
|---|---|---|---|
| TT | CC | 79 (50.00) | 58 (58.00) |
| TC | CC | 40 (25.32) | 24 (24.00) |
| CC | CC | 14 (8.86) | 2 (2.00) |
| TT | CG | 14 (8.86) | 8 (8.00) |
| TC | CG | 10 (6.33) | 8 (8.00) |
| CC | CG | 0 (0.00) | 0 (0.00) |
| TT | GG | 0 (0.00) | 0 (0.00) |
| TC | GG | 1 (0.63) | 0 (0.00) |
| CC | GG | 0 (0.00) | 0 (0.00) |
| Variable, Category, Parameter | NOTCH3 Polymorphism | PSMA6 Polymorphism | ||||
|---|---|---|---|---|---|---|
| C Allele Carriers (CC + TC) (n = 65) | Wild-Type Homozygotes (TT) (n = 93) | p | G Allele Carriers (GG + CG) (n = 25) | Wild-Type Homozygotes (CC) (n = 133) | p | |
| Gender, male, n (%) | 46 (70.77) | 66 (70.97) | 0.558 | 18 (72.00) | 94 (70.68) | 0.551 |
| Age of onset, M ± SD | 22.9 ± 11.2 | 23.6 ± 13.1 | 0.727 | 23.7 ± 12.8 | 23.2 ± 12.3 | 0.855 |
| Psoriasis subtype, over 40 years of age, n (%) | 6 (9.23) | 15 (16.13) | 0.154 | 3 (12.00) | 18 (13.53) | 0.567 |
| Psoriasis arthritis, n (%) | 25 (39.68) | 31 (33.33) | 0.310 | 5 (20.00) | 51 (38.35) | 0.059 |
| Severity, moderate or severe, n (%) | 47 (72.31) | 71 (76.34) | 0.347 | 20 (80.00) | 98 (73.68) | 0.348 |
| Positive family history, n (%) * | 27 (49.09) | 40 (50.00) | 0.528 | 13 (54.17) | 54 (48.65) | 0.396 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartosińska, J.; Zmorzyński, S.; Sarecka-Hujar, B.; Raczkiewicz, D.; Wojcierowska-Litwin, M.; Korszeń-Pilecka, I.; Michalak-Stoma, A.; Kowal, M.; Bartosiński, J.; Filip, A.; et al. The Genetic Variants of NOTCH3 (6746T>C) and PSMA6 (-8C>G) as Possible Risk Factors of Psoriasis Development. Life 2021, 11, 887. https://doi.org/10.3390/life11090887
Bartosińska J, Zmorzyński S, Sarecka-Hujar B, Raczkiewicz D, Wojcierowska-Litwin M, Korszeń-Pilecka I, Michalak-Stoma A, Kowal M, Bartosiński J, Filip A, et al. The Genetic Variants of NOTCH3 (6746T>C) and PSMA6 (-8C>G) as Possible Risk Factors of Psoriasis Development. Life. 2021; 11(9):887. https://doi.org/10.3390/life11090887
Chicago/Turabian StyleBartosińska, Joanna, Szymon Zmorzyński, Beata Sarecka-Hujar, Dorota Raczkiewicz, Magdalena Wojcierowska-Litwin, Iwona Korszeń-Pilecka, Anna Michalak-Stoma, Małgorzata Kowal, Jarosław Bartosiński, Agata Filip, and et al. 2021. "The Genetic Variants of NOTCH3 (6746T>C) and PSMA6 (-8C>G) as Possible Risk Factors of Psoriasis Development" Life 11, no. 9: 887. https://doi.org/10.3390/life11090887