
ADCY5, CAPN10 and JAZF1 Gene Polymorphisms and Placental Expression in Women with Gestational Diabetes

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Methods
2.3. RNA Isolation
2.4. Reverse-Transcription (RT-PCR)
2.5. Real-Time Quantitative Reverse Transcription PCR (RQ-PCR)
2.6. Statistical Analysis
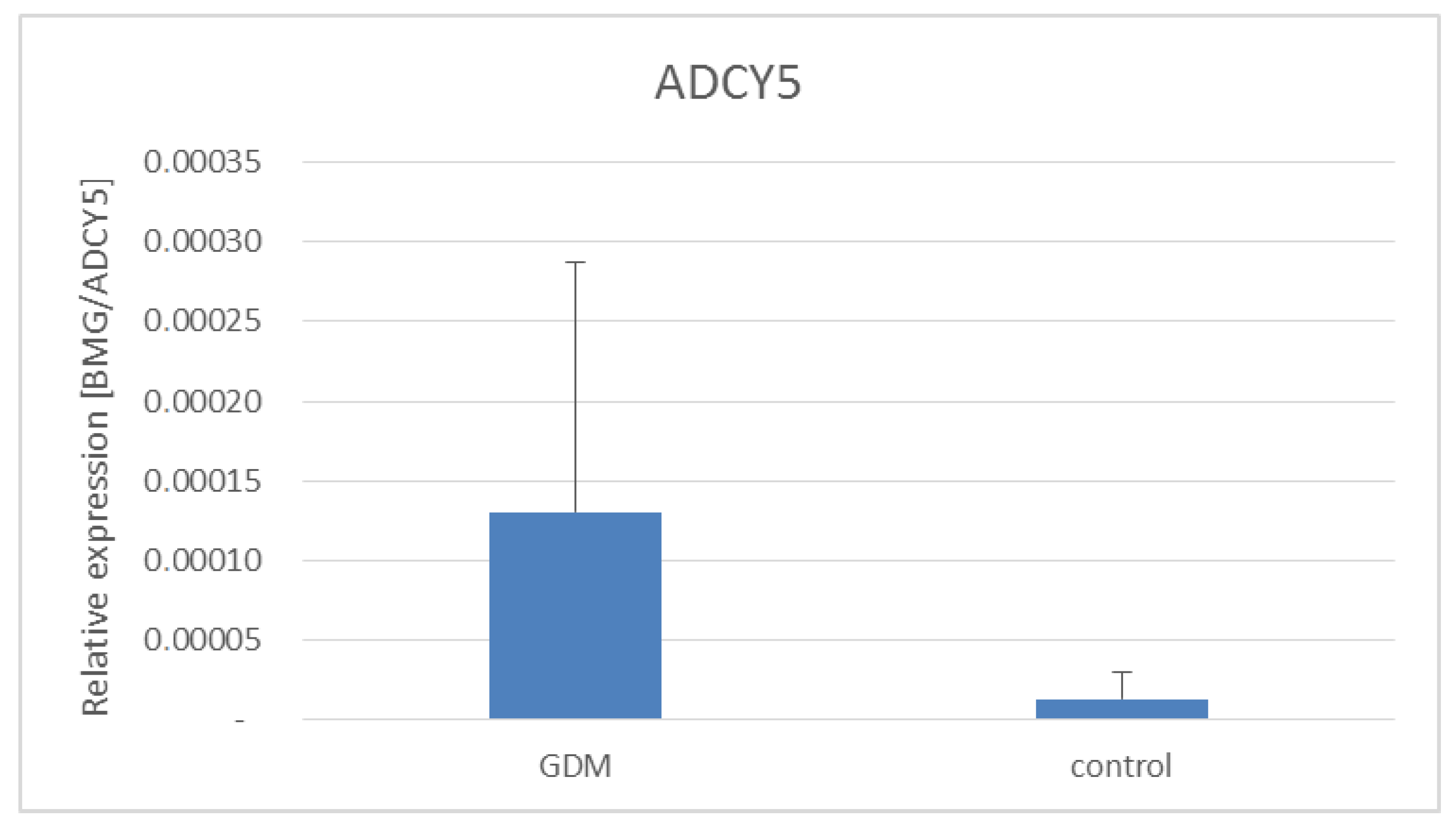
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sullivan, S.D.; Umans, J.G.; Ratner, R. Gestational diabetes: Implications for cardiovascular health. Curr. Diab. Rep. 2012, 12, 43–52. [Google Scholar] [CrossRef]
- Popova, P.V.; Grineva, E.N.; Gerasimov, A.S.; Kravchuk, E.N.; Ryazantseva, E.M.; Shelepova, E.S. The new combination of risk factors determining a high risk of gestational diabetes mellitus. Minerva Endocrinol. 2015, 40, 239–247. [Google Scholar] [PubMed]
- Popova, P.; Tkachuk, A.; Dronova, A.; Gerasimov, A.; Kravchuk, E.; Bolshakova, M.; Rozdestvenskaya, O.; Demidova, K.; Nikolaeva, A.; Grineva, E. Fasting glycemia at the first prenatal visit and pregnancy outcomes in Russia women. Minerva Endocrinol. 2016, 41, 477–485. [Google Scholar]
- Popova, P.V.; Klyushina, A.A.; Vasilyeva, L.B.; Tkachuk, A.S.; Vasukova, E.A.; Anopova, A.D.; Pustozerov, E.A.; Gorelova, I.; Kravchuk, E.N.; Li, O.; et al. Association of Common Genetic Risk Variants With Gestational Diabetes Mellitus and Their Role in GDM Prediction. Front. Endocrinol. 2021, 12, 628582. [Google Scholar] [CrossRef] [PubMed]
- Popova, P.; Vasilyeva, L.; Tkachuck, A.; Puzanov, M.; Golovkin, A.; Bolotko, Y.; Pustozerov, E.; Vasilyeva, E.; Li, O.; Zazerskaya, I.; et al. A Randomised, Controlled Study of Different Glycaemic Targets during Gestational Diabetes Treatment: Effect on the Level of Adipokines in Cord Blood and ANGPTL4 Expression in Human Umbilical Vein Endothelial Cells. Int. J. Endocrinol. 2018, 2018, 6481658. [Google Scholar] [CrossRef] [PubMed]
- Barbour, L.A.; McCurdy, C.E.; Hernandez, T.L.; Kirwan, J.P.; Catalano, P.M.; Friedman, J.E. Cellular mechanisms for insulin resistance in normal pregnancy and gestational diabetes. Diabetes Care 2007, 30 (Suppl. 2), S112–S119. [Google Scholar] [CrossRef] [Green Version]
- Roman, T.S.; Cannon, M.E.; Vadlamudi, S.; Buchkovich, M.L.; Wolford, B.N.; Welch, R.P.; Morken, M.A.; Kwon, G.J.; Varshney, A.; Kursawe, R.; et al. A Type 2 Diabetes-Associated Functional Regulatory Variant in a Pancreatic Islet Enhancer at the ADCY5 Locus. Diabetes 2017, 66, 2521–2530. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; You, N.C.; Hsu, Y.H.; Sul, J.; Wang, L.; Tinker, L.; Eaton, C.B.; Liu, S. Common genetic variation in calpain-10 gene (CAPN10) and diabetes risk in a multi-ethnic cohort of American postmenopausal women. Hum. Mol. Genet. 2007, 16, 2960–2971. [Google Scholar] [CrossRef] [Green Version]
- Langberg, K.A.; Ma, L.; Sharma, N.K.; Hanis, C.L.; Elbein, S.C.; Hasstedt, S.J.; Das, S.K.; American Diabetes Association GENNID Study Group. Single nucleotide polymorphisms in JAZF1 and BCL11A gene are nominally associated with type 2 diabetes in African-American families from the GENNID study. J. Hum. Genet. 2012, 57, 57–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatner, S.F.; Park, M.; Yan, L.; Lee, G.J.; Lai, L.; Iwatsubo, K.; Ishikawa, Y.; Pessin, J.; Vatner, D.E. Adenylyl cyclase type 5 in cardiac disease, metabolism, and aging. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H1–H8. [Google Scholar] [CrossRef] [Green Version]
- Shibasaki, T.; Sunaga, Y.; Seino, S. Integration of ATP, cAMP, and Ca2+ signals in insulin granule exocytosis. Diabetes 2004, 53 (Suppl. 3), S59–S62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodson, D.J.; Mitchell, R.K.; Marselli, L.; Pullen, T.J.; Gimeno Brias, S.; Semplici, F.; Everett, K.L.; Cooper, D.M.; Bugliani, M.; Marchetti, P.; et al. ADCY5 couples glucose to insulin secretion in human islets. Diabetes 2014, 63, 3009–3021. [Google Scholar] [CrossRef] [Green Version]
- van de Bunt, M.; Manning Fox, J.E.; Dai, X.; Barrett, A.; Grey, C.; Li, L.; Bennett, A.J.; Johnson, P.R.; Rajotte, R.V.; Gaulton, K.J.; et al. Transcript Expression Data from Human Islets Links Regulatory Signals from Genome-Wide Association Studies for Type 2 Diabetes and Glycemic Traits to Their Downstream Effectors. PLoS Genet. 2015, 11, e1005694. [Google Scholar] [CrossRef] [PubMed]
- Varshney, A.; Scott, L.J.; Welch, R.P.; Erdos, M.R.; Chines, P.S.; Narisu, N.; Albanus, R.D.; Orchard, P.; Wolford, B.N.; Kursawe, R.; et al. Genetic regulatory signatures underlying islet gene expression and type 2 diabetes. Proc. Natl. Acad. Sci. USA 2017, 114, 2301–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horikawa, Y.; Oda, N.; Cox, N.J.; Li, X.; Orho-Melander, M.; Hara, M.; Hinokio, Y.; Lindner, T.H.; Mashima, H.; Schwarz, P.E.; et al. Genetic variation in the gene encoding calpain-10 is associated with type 2 diabetes mellitus. Nat. Genet. 2000, 26, 163–175. [Google Scholar]
- Fogarty, M.P.; Panhuis, T.M.; Vadlamudi, S.; Buchkovich, M.L.; Mohlke, K.L. Allele-specific transcriptional activity at type 2 diabetes-associated single nucleotide polymorphisms in regions of pancreatic islet open chromatin at the JAZF1 locus. Diabetes 2013, 62, 1756–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Association of Diabetes and Pregnancy Study Groups Consensus Panel; Metzger, B.E.; Gabbe, S.G.; Persson, B.; Buchanan, T.A.; Catalano, P.A.; Damm, P.; Dyer, A.R.; Leiva, A.D.; Hod, M.; et al. International association of diabetes and pregnancy study groups recommendations on the diagnosis and classification of hyperglycemia in pregnancy. Diabetes Care 2010, 33, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Jensen, D.P.; Urhammer, S.A.; Eiberg, H.; Borch-Johnsen, K.; Jørgensen, T.; Hansen, T.; Pedersen, O. Variation in CAPN10 in relation to type 2 diabetes, obesity and quantitative metabolic traits: Studies in 6018 whites. Mol. Genet. Metab. 2006, 89, 360–367. [Google Scholar] [CrossRef]
- Grarup, N.; Andersen, G.; Krarup, N.T.; Albrechtsen, A.; Schmitz, O.; Jørgensen, T.; Borch-Johnsen, K.; Hasen, T.; Pedersen, O. Association testing of novel type 2 diabetes risk alleles in the JAZF1, CDC123/CAMK1D, TSPAN8, THADA, ADAMTS9, and NOTCH2 loci with insulin release, insulin sensitivity, and obesity in a population-based sample of 4516 glucose-tolerant middle-aged Danes. Diabetes 2008, 57, 2534–2540. [Google Scholar] [CrossRef] [Green Version]
- Sohani, Z.N.; Deng, W.Q.; Pare, G.; Meyre, D.; Gerstein, H.C.; Anand, S.S. Does genetic heterogeneity account for the divergent risk of type 2 diabetes in South Asian and white European populations? Diabetologia 2014, 57, 2270–2281. [Google Scholar] [CrossRef] [Green Version]
- Arora, G.P.; Almgren, P.; Brøns, C.; Thaman, R.G.; Vaag, A.A.; Groop, L.; Prasad, R.B. Association between genetic risk variants and glucose intolerance during pregnancy in north Indian women. BMC Med. Genom. 2018, 11, 64. [Google Scholar] [CrossRef]
- Andersson, E.A.; Pilgaard, K.; Pisinger, C.; Harder, M.N.; Grarup, N.; Faerch, K.; Poulsen, P.; Witte, D.R.; Jørgensen, T.; Vaag, A.; et al. Type 2 diabetes risk alleles near ADCY5, CDKAL1 and HHEX-IDE are associated with reduced birthweight. Diabetologia 2010, 53, 1908–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, I.A.; Movva, S.; Shaik, N.A.; Chava, S.; Jahan, P.; Mukkavali, K.K.; Kamineni, V.; Hasan, Q.; Rao, P. Investigation of Calpain 10 (rs2975760) gene polymorphism in Asian Indians with Gestational Diabetes Mellitus. Meta Gene 2014, 2, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, C.; Wei, L.; Sun, F.; Ji, L. The Association Between the rs2975760 and rs3792267 Single Nucleotide Polymorphisms of Calpain 10 (CAPN10) and Gestational Diabetes Mellitus. Med. Sci. Monit. 2019, 25, 5137–5142. [Google Scholar] [CrossRef]
- Stuebe, A.M.; Wise, A.; Nguyen, T.; Herring, A.; North, K.E.; Siega-Riz, A.M. Maternal genotype and gestational diabetes. Am. J. Perinatol. 2014, 31, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; He, J.; Xu, X.L.; Jiang, J.; He, C.Q.; Ma, H.M. Molecular characterization and tissue expression profile analysis of the porcine JAZF1 gene. Genet. Mol. Res. 2015, 14, 542–551. [Google Scholar] [CrossRef]
- Yuan, L.; Luo, X.; Zeng, M.; Zhang, Y.; Yang, M.; Zhang, L.; Liu, R.; Boden, G.; Liu, H.; Ma, Z.A.; et al. Transcription factor TIP27 regulates glucose homeostasis and insulin sensitivity in a PI3-kinase/Akt-dependent manner in mice. Int. J. Obes. 2015, 39, 949–958. [Google Scholar] [CrossRef]
- Li, L.; Yang, Y.; Yang, G.; Lu, C.; Yang, M.; Liu, H.; Zong, H. The role of JAZF1 on lipid metabolism and related genes in vitro. Metabolism 2011, 60, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Ming, G.F.; Xiao, D.; Gong, W.J.; Liu, H.X.; Liu, J.; Zhou, H.H.; Liu, Z.Q. JAZF1 can regulate the expression of lipid metabolic genes and inhibit lipid accumulation in adipocytes. Biochem. Biophys. Res. Commun. 2014, 445, 673–680. [Google Scholar] [CrossRef]
- Wei, Q.; Zhou, B.; Yang, G.; Hu, W.; Zhang, L.; Liu, R.; Li, M.; Wang, K.; Gu, H.F.; Guan, Y.; et al. JAZF1 ameliorates age and diet-associated hepatic steatosis through SREBP-1c -dependent mechanism. Cell Death Dis. 2018, 9, 859. [Google Scholar] [CrossRef]
- Kobiita, A.; Godbersen, S.; Araldi, E.; Ghoshdastider, U.; Schmid, M.W.; Spinas, G.; Moch, H.; Stoffel, M. The Diabetes Gene JAZF1 Is Essential for the Homeostatic Control of Ribosome Biogenesis and Function in Metabolic Stress. Cell Rep. 2020, 32, 107846. [Google Scholar] [CrossRef]
- Rasheed, M.A.; Kantoush, N.; Abd El-Ghaffar, N.; Farouk, H.; Kamel, S.; Ibrahim, A.A.; Shalaby, A.; Mahmoud, E.; Raslan, H.M.; Saleh, O.M. Expression of JAZF1, ABCC8, KCNJ11and Notch2 genes and vitamin D receptor polymorphisms in type 2 diabetes, and their association with microvascular complications. Ther. Adv. Endocrinol. Metab. 2017, 8, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Lin, Y.; Yang, M.; Li, M.; Yang, G.; Hao, P.; Li, L. JAZF1 Inhibits Adipose Tissue Macrophages and Adipose Tissue Inflammation in Diet-Induced Diabetic Mice. Biomed. Res. Int. 2018, 2018, 4507659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taneera, J.; Lang, S.; Sharma, A.; Fadista, J.; Zhou, Y.; Ahlqvist, E.; Jonsson, A.; Lyssenko, V.; Vikman, P.; Hansson, O.; et al. A systems genetics approach identifies genes and pathways for type 2 diabetes in human islets. Cell Metab. 2012, 16, 122–134. [Google Scholar] [CrossRef] [Green Version]
- Marshall, C.; Hitman, G.A.; Partridge, C.J.; Clark, A.; Ma, H.; Shearer, T.R.; Turner, M.D. Evidence that an isoform of calpain-10 is a regulator of exocytosis in pancreatic beta-cells. Mol. Endocrinol. 2005, 19, 213–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parnaud, G.; Hammar, E.; Rouiller, D.G.; Bosco, D. Inhibition of calpain blocks pancreatic beta-cell spreading and insulin secretion. Am. J. Physiol Endocrinol. Metab. 2005, 289, E313–E321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baier, L.J.; Permana, P.A.; Yang, X.; Pratley, R.E.; Hanson, R.L.; Shen, G.Q.; Mott, D.; Knowler, W.C.; Cox, N.J.; Horikawa, Y.; et al. A calpain-10 gene polymorphism is associated with reduced muscle mRNA levels and insulin resistance. J. Clin. Invest. 2000, 106, R69–R73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlsson, E.; Poulsen, P.; Storgaard, H.; Almgren, P.; Ling, C.; Jensen, C.B.; Madsbad, S.; Groop, L.; Vaag, A.; Ridderstråle, M. Genetic and nongenetic regulation of CAPN10 mRNA expression in skeletal muscle. Diabetes 2005, 54, 3015–3020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otani, K.; Han, D.H.; Ford, E.L.; Garcia-Roves, P.M.; Ye, H.; Horikawa, Y.; Bell, G.I.; Holloszy, J.O.; Polonsky, K.S. Calpain system regulates muscle mass and glucose transporter GLUT4 turnover. J. Biol. Chem. 2004, 279, 20915–20920. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Parameters | Control Group | GDM | p-Value & |
|---|---|---|---|
| Mean ± SD | Mean ± SD | ||
| Age (years) | 30.3 ± 4.6 | 31.7 ± 5.4 | 0.00014 |
| Height (cm) | 166.1 ± 6.1 | 165.9 ± 6.2 | 0.75 |
| Pregnancy number | 1.9 ± 1.1 | 2.1 ± 1.2 | 0.27 |
| Fasting glucose in an oral glucose tolerance test (mg/dL) | 80.3 ± 6.7 | 95.7 ± 11.6 | <0.00001 |
| Glucose after 1 h in an oral glucose tolerance test (mg/dL) | 122.2 ± 23.6 | 156.9 ± 37.1 | <0.00001 |
| Glucose after 2 h in an oral glucose tolerance test (mg/dL) | 102.3 ± 18.7 | 130.9 ± 34.1 | <0.00001 |
| Body mass before pregnancy (kg) | 64.3 ± 13.4 | 73.3 ± 17.5 | <0.00001 |
| Body mass at birth (kg) | 78.1 ± 13.4 | 84.8 ± 15.9 | <0.00001 |
| Body mass increase during pregnancy (kg) | 13.8 ± 5.4 | 11.5 ± 7.2 | <0.00001 |
| BMI before pregnancy (kg/m2) | 23.3 ± 4.3 | 26.6 ± 6.2 | <0.00001 |
| BMI at birth (kg/m2) | 28.3 ± 4.3 | 30.8 ± 5.7 | <0.00001 |
| BMI increase during pregnancy (kg/m2) | 5.0 ± 1.9 | 4.2 ± 2.7 | <0.00001 |
| Newborn body mass (g) | 3287 ± 529 | 3286 ± 566 | 0.97 |
| APGAR (0–10) | 9.4 ± 1.0 | 9.3 ± 1.1 | 0.13 |
| Genotypes and Alleles | Control Group | GDM | p-Value ^ | Compared Genotypes or Alleles | OR (95% CI) | p-Value ^ | ||
|---|---|---|---|---|---|---|---|---|
| n | % | n | % | |||||
| ADCY5rs11708067 genotype | ||||||||
| AA | 244 | 70.11% | 193 | 70.96% | 0.69 | GG+AG vs. AA | 0.96 (0.68–1.36) | 0.82 |
| AG | 97 | 27.87% | 71 | 26.10% | GG vs. AG+AA | 1.48 (0.53–4.12) | 0.45 | |
| GG | 7 | 2.01% | 8 | 2.94% | GG vs. AA | 1.44 (0.51–4.05) | 0.48 | |
| AG vs. AA | 0.93 (0.65–1.33) | 0.67 | ||||||
| AG vs. AA | 1.56 (0.54–4.50) | 0.41 | ||||||
| Allele | ||||||||
| A | 585 | 84.05% | 457 | 84.01% | G vs. A | 1.00 (0.74–1.36) | 0.98 | |
| G | 111 | 15.95% | 87 | 15.99% | ||||
| ADCY5rs2877716 genotype | ||||||||
| CC | 237 | 68.10% | 185 | 68.01% | 1.00 | TT+CT vs. CC | 1.00 (0.71–1.41) | 0.98 |
| CT | 102 | 29.31% | 80 | 29.41% | TT vs. CT+CC | 0.99 (0.37–2.71) | 0.99 | |
| TT | 9 | 2.59% | 7 | 2.57% | TT vs. CC | 1.00 (0.36–2.73) | 0.99 | |
| CT vs. CC | 1.00 (0.71–1.43) | 0.98 | ||||||
| TT vs. CT | 0.99 (0.35–2.78) | 0.99 | ||||||
| Allele | ||||||||
| C | 576 | 82.76% | 450 | 82.72% | T vs. C | 1.00 (0.75–1.35) | 0.99 | |
| T | 120 | 17.24% | 94 | 17.28% | ||||
| CAPN10rs2975760 genotype | ||||||||
| TT | 261 | 75.22% | 196 | 72.59% | 0.74 | CC+TC vs. TT | 1.15 (0.80–1.65) | 0.46 |
| TC | 77 | 22.19% | 67 | 24.81% | CC vs. TC+TT | 1.00 (0.37–2.72) | 1.00 | |
| CC | 9 | 2.59% | 7 | 2.59% | CC vs. TT | 1.04 (0.38–2.83) | 0.95 | |
| TC vs. TT | 1.16 (0.80–1.69) | 0.44 | ||||||
| CC vs. TC | 0.89 (0.32–2.53) | 0.83 | ||||||
| Allele | ||||||||
| T | 599 | 86.31% | 459 | 85.00% | C vs. T | 1.11 (0.81–1.53) | 0.51 | |
| C | 95 | 13.69% | 81 | 15.00% | ||||
| CAPN10 rs3792267 genotype | ||||||||
| GG | 175 | 50.43% | 132 | 48.89% | 0.74 | AA+GA vs. GG | 1.06 (0.77–1.46) | 0.70 |
| GA | 129 | 37.18% | 108 | 40.00% | AA vs. GA+GG | 0.88 (0.54–1.45) | 0.63 | |
| AA | 43 | 12.39% | 30 | 11.11% | AA vs. GG | 0.92 (0.55–1.55) | 0.77 | |
| GA vs. GG | 1.11 (0.79–1.56) | 0.55 | ||||||
| AA vs. GA | 0.83 (0.49–1.42) | 0.50 | ||||||
| Allele | A vs. G | 1.01 (0.79–1.28) | 0.96 | |||||
| G | 479 | 69.02% | 372 | 68.89% | ||||
| A | 215 | 30.98% | 168 | 31.11% | ||||
| JAZF1 rs864745 genotype | ||||||||
| TT | 69 | 19.88% | 79 | 29.26% | 0.022 | CC+CT vs. TT | 0.60 (0.41–0.87) | 0.006 |
| CT | 191 | 55.04% | 136 | 50.37% | CC vs. CT+TT | 0.76 (0.52–1.12) | 0.17 | |
| CC | 87 | 25.07% | 55 | 20.37% | CC vs. TT | 0.55 (0.35–0.88) | 0.012 | |
| CT vs. TT | 0.62 (0.42–0.92) | 0.017 | ||||||
| CC vs. CT | 0.89 (0.59–1.33) | 0.56 | ||||||
| Allele | ||||||||
| T | 329 | 47.41% | 294 | 54.44% | C vs. T | 0.75 (0.60–0.95) | 0.014 | |
| C | 365 | 52.59% | 246 | 45.56% | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ustianowski, P.; Malinowski, D.; Kopytko, P.; Czerewaty, M.; Tarnowski, M.; Dziedziejko, V.; Safranow, K.; Pawlik, A. ADCY5, CAPN10 and JAZF1 Gene Polymorphisms and Placental Expression in Women with Gestational Diabetes. Life 2021, 11, 806. https://doi.org/10.3390/life11080806
Ustianowski P, Malinowski D, Kopytko P, Czerewaty M, Tarnowski M, Dziedziejko V, Safranow K, Pawlik A. ADCY5, CAPN10 and JAZF1 Gene Polymorphisms and Placental Expression in Women with Gestational Diabetes. Life. 2021; 11(8):806. https://doi.org/10.3390/life11080806
Chicago/Turabian StyleUstianowski, Przemysław, Damian Malinowski, Patrycja Kopytko, Michał Czerewaty, Maciej Tarnowski, Violetta Dziedziejko, Krzysztof Safranow, and Andrzej Pawlik. 2021. "ADCY5, CAPN10 and JAZF1 Gene Polymorphisms and Placental Expression in Women with Gestational Diabetes" Life 11, no. 8: 806. https://doi.org/10.3390/life11080806