
The Effect of a Total Fishmeal Replacement by Arthrospira platensis on the Microbiome of African Catfish (Clarias gariepinus)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Rearing Conditions
2.2. Microbiome
2.2.1. Sampling
2.2.2. DNA Extraction
2.2.3. 16S rRNA Gene Amplification and Sequencing
2.2.4. Sequence Processing and Analyses
2.3. Data Processing
2.4. Sequence Data Deposition
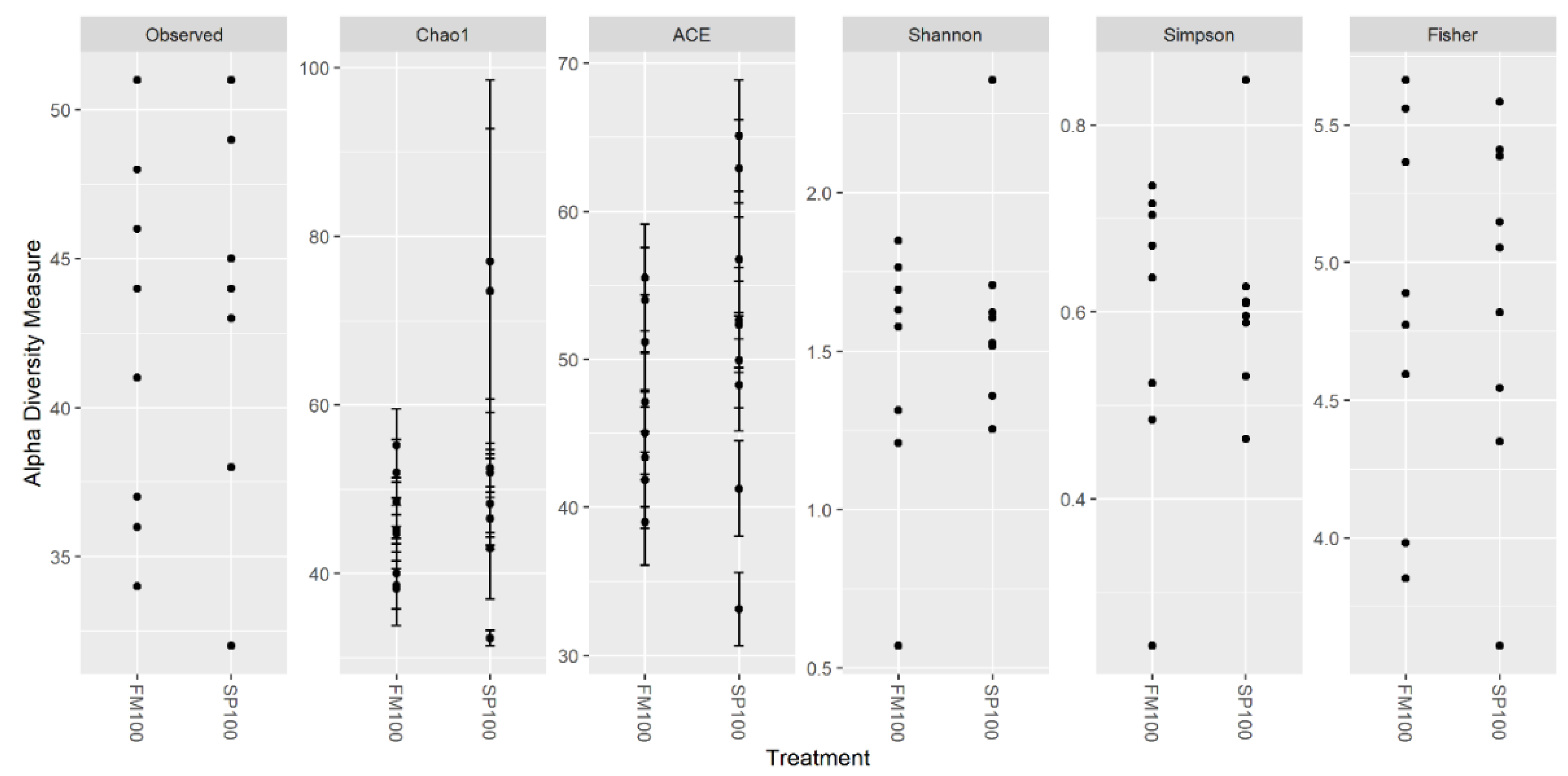
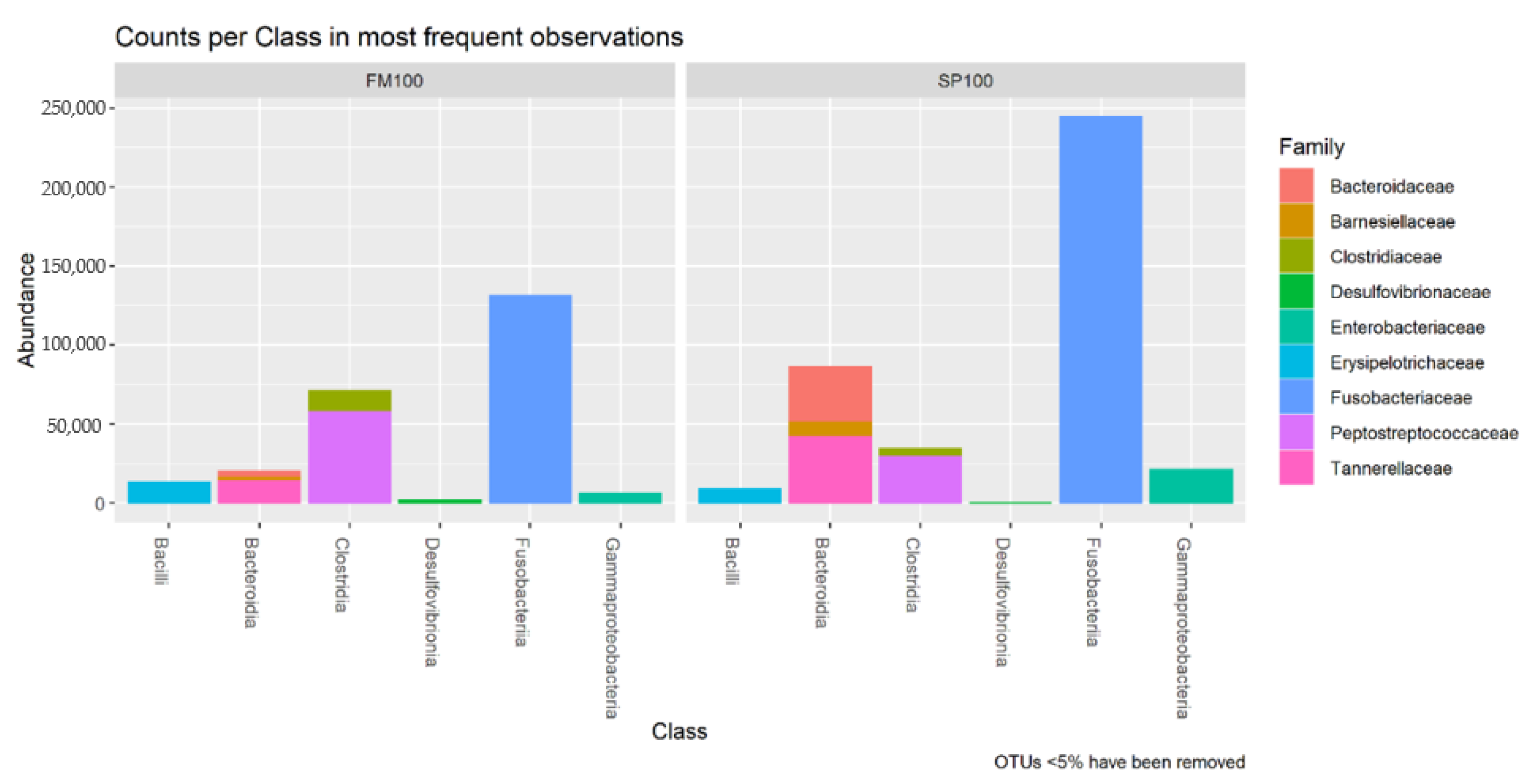
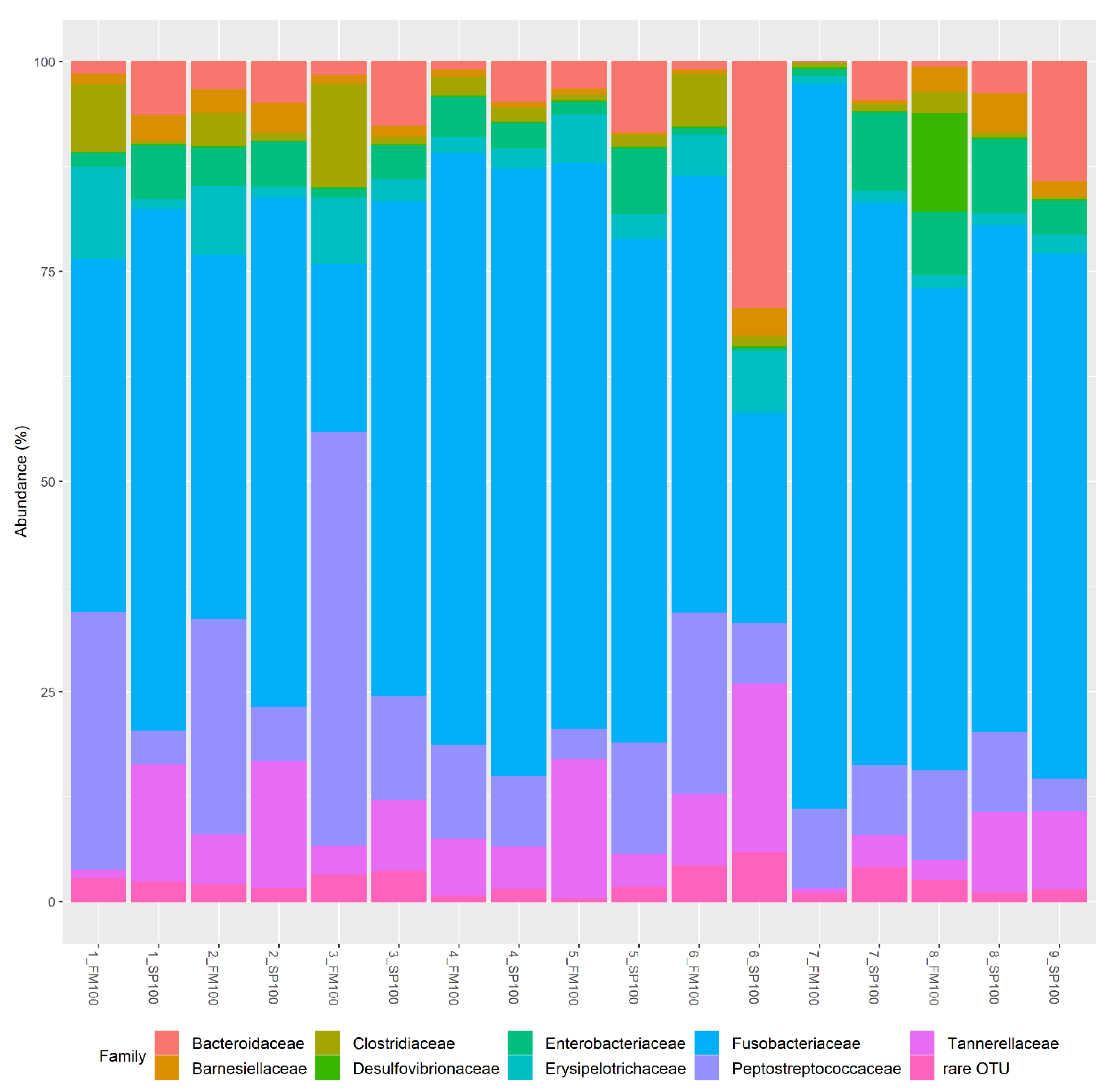
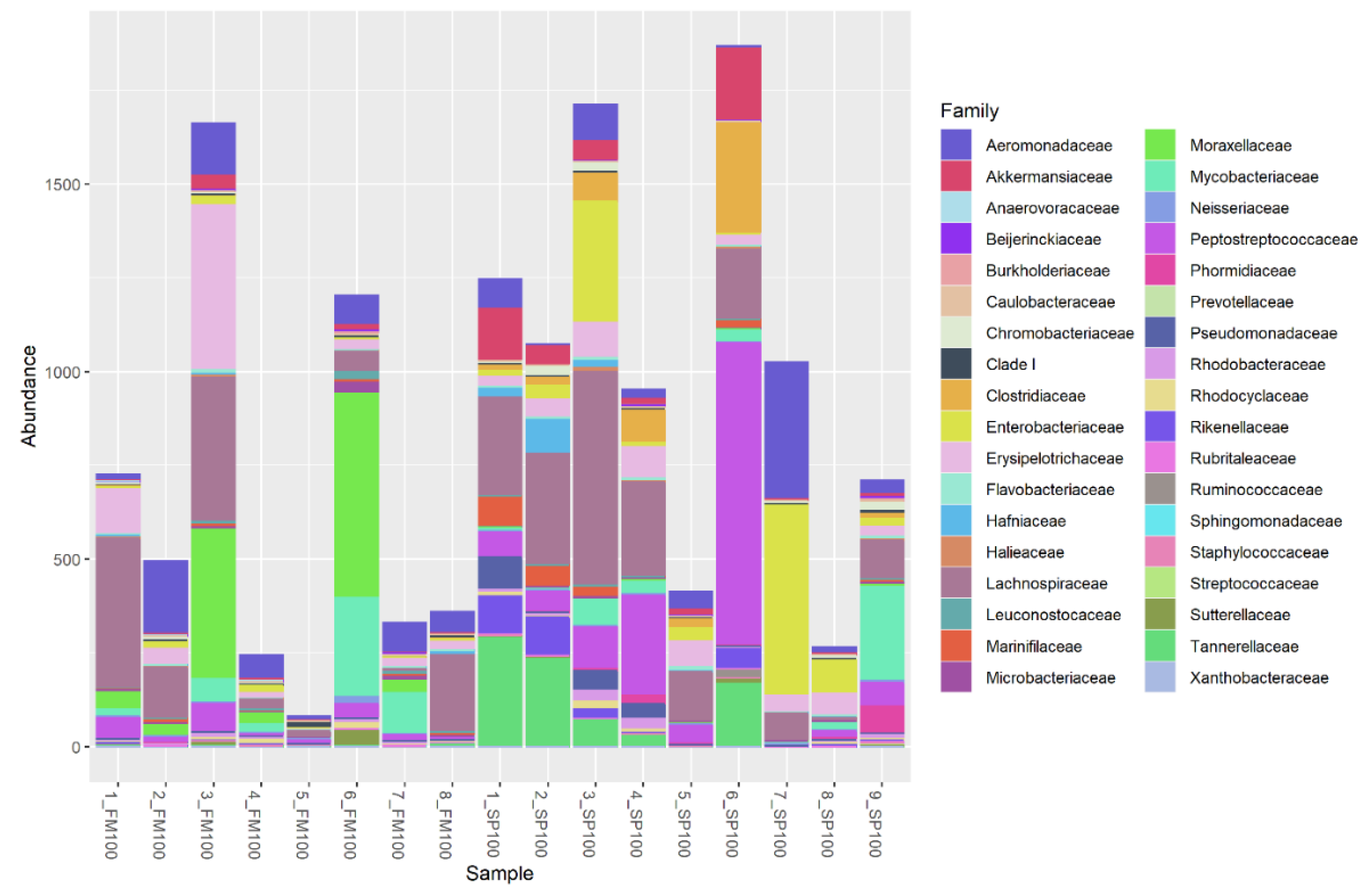
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. State of World Fisheries and Aquaculture 2020: Sustainability in Action; FOOD & AGRICULTURE ORG: Rome, Italy, 2016; ISBN 978-92-5-132692-3. [Google Scholar]
- World Bank. Fish to 2030: Prospects for Fisheries and Aquaculture; World Bank: Washington, DC, USA, 2013. [Google Scholar]
- García, S.M. The Ecosystem Approach to Fisheries: Issues, Terminology, Principles, Institutional Foundations, Implementation and Outlook; Food and Agriculture Organization of the United Nations: Rome, Italy, 2003; ISBN 9251049602. [Google Scholar]
- Becker, E.W. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Rana, K.J.; Siriwardena, S.; Hasan, M.R. Impact of Rising Feed Ingredient Prices on Aquafeeds and Aquaculture Production; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009; ISBN 978-92-5-106422-1. [Google Scholar]
- Ytrestøyl, T.; Aas, T.S.; Åsgård, T. Utilisation of feed resources in production of Atlantic salmon (Salmo salar) in Norway. Aquaculture 2015, 448, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Ragaza, J.A.; Hossain, M.S.; Meiler, K.A.; Velasquez, S.F.; Kumar, V. A review on Spirulina: Alternative media for cultivation and nutritive value as an aquafeed. Rev. Aquac. 2020, 12, 2371–2395. [Google Scholar] [CrossRef]
- Yarnold, J.; Karan, H.; Oey, M.; Hankamer, B. Microalgal Aquafeeds As Part of a Circular Bioeconomy. Trends Plant Sci. 2019, 24, 959–970. [Google Scholar] [CrossRef] [Green Version]
- Dernekbasi, S.; Una, H.; Karayucel, I.; Aral, O. Effect of Dietary Supplementation of Different Rates of Spirulina (Spirulina platensis) on Growth and Feed Conversion in Guppy (Poecilia reticulata Peters, 1860). J. Anim. Vet. Adv. 2010, 9, 1395–1399. [Google Scholar] [CrossRef] [Green Version]
- Raji, A.A.; Alaba, P.A.; Yusuf, H.; Abu Bakar, N.H.; Mohd Taufek, N.; Muin, H.; Alias, Z.; Milow, P.; Abdul Razak, S. Fishmeal replacement with Spirulina Platensis and Chlorella vulgaris in African catfish (Clarias gariepinus) diet: Effect on antioxidant enzyme activities and haematological parameters. Res. Vet. Sci. 2018, 119, 67–75. [Google Scholar] [CrossRef]
- Promya, J.; Chitmanat, C. The effects of Spirulina platensis and Cladophora Algae on the Growth Performance, Meat Quality and Immunity Stimulating Capacity of the African Sharptooth Catfish (Clarias gariepinus). Int. J. Agric. Biol. 2011, 13, 77–82. [Google Scholar]
- Jafari, S.M.A.; Rabbani, M.; Emtyazjoo, M.; Piryaei, F. Effect of dietary Spirulina platensis on fatty acid composition of rainbow trout (Oncorhynchus mykiss) fillet. Aquac. Int. 2014, 22, 1307–1315. [Google Scholar] [CrossRef]
- Roohani, A.M.; Abedian Kenari, A.; Fallahi Kapoorchali, M.; Borani, M.S.; Zoriezahra, S.J.; Smiley, A.H.; Esmaeili, M.; Rombenso, A.N. Effect of spirulina Spirulina platensis as a complementary ingredient to reduce dietary fish meal on the growth performance, whole-body composition, fatty acid and amino acid profiles, and pigmentation of Caspian brown trout (Salmo trutta caspius) juveniles. Aquac. Nutr. 2019, 25, 633–645. [Google Scholar] [CrossRef]
- Teimouri, M.; Yeganeh, S.; Amirkolaie, A.K. The effects of Spirulina platensis meal on proximate composition, fatty acid profile and lipid peroxidation of rainbow trout (Oncorhynchus mykiss) muscle. Aquac. Nutr. 2016, 22, 559–566. [Google Scholar] [CrossRef]
- Desai, A.R.; Links, M.G.; Collins, S.A.; Mansfield, G.S.; Drew, M.D.; van Kessel, A.G.; Hill, J.E. Effects of plant-based diets on the distal gut microbiome of rainbow trout (Oncorhynchus mykiss). Aquaculture 2012, 350–353, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Estruch, G.; Collado, M.C.; Peñaranda, D.S.; Tomás Vidal, A.; Jover Cerdá, M.; Pérez Martínez, G.; Martinez-Llorens, S. Impact of Fishmeal Replacement in Diets for Gilthead Sea Bream (Sparus aurata) on the Gastrointestinal Microbiota Determined by Pyrosequencing the 16S rRNA Gene. PLoS ONE 2015, 10, e0136389. [Google Scholar] [CrossRef]
- Ingerslev, H.-C.; von Gersdorff Jørgensen, L.; Lenz Strube, M.; Larsen, N.; Dalsgaard, I.; Boye, M.; Madsen, L. The development of the gut microbiota in rainbow trout (Oncorhynchus mykiss) is affected by first feeding and diet type. Aquaculture 2014, 424-425, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Ringø, E.; Sperstad, S.; Myklebust, R.; Refstie, S.; Krogdahl, Å. Characterisation of the microbiota associated with intestine of Atlantic cod (Gadus morhua L.). Aquaculture 2006, 261, 829–841. [Google Scholar] [CrossRef]
- Schmidt, V.; Amaral-Zettler, L.; Davidson, J.; Summerfelt, S.; Good, C. Influence of Fishmeal-Free Diets on Microbial Communities in Atlantic Salmon (Salmo salar) Recirculation Aquaculture Systems. Appl. Environ. Microbiol. 2016, 82, 4470–4481. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.H.; Lin, G.; Fu, G.H.; Wan, Z.Y.; Lee, M.; Wang, L.; Liu, X.J.; Yue, G.H. The intestinal microbiome of fish under starvation. BMC Genom. 2014, 15, 266. [Google Scholar] [CrossRef] [Green Version]
- Zarkasi, K.Z.; Taylor, R.S.; Abell, G.C.J.; Tamplin, M.L.; Glencross, B.D.; Bowman, J.P. Atlantic Salmon (Salmo salar L.) Gastrointestinal Microbial Community Dynamics in Relation to Digesta Properties and Diet. Microb. Ecol. 2016, 71, 589–603. [Google Scholar] [CrossRef]
- de Souza, F.P.; de Lima, E.C.S.; Urrea-Rojas, A.M.; Suphoronski, S.A.; Facimoto, C.T.; da Silva Bezerra Júnior, J.; de Oliveira, T.E.S.; de Pádua Pereira, U.; Di Santis, G.W.; de Oliveira, C.A.L.; et al. Effects of dietary supplementation with a microalga (Schizochytrium sp.) on the hemato-immunological, and intestinal histological parameters and gut microbiota of Nile tilapia in net cages. PLoS ONE 2020, 15, e0226977. [Google Scholar] [CrossRef] [Green Version]
- Cerezuela, R.; Fumanal, M.; Tapia-Paniagua, S.T.; Meseguer, J.; Moriñigo, M.A.; Esteban, M.A. Histological alterations and microbial ecology of the intestine in gilthead seabream (Sparus aurata L.) fed dietary probiotics and microalgae. Cell Tissue Res. 2012, 350, 477–489. [Google Scholar] [CrossRef]
- Lyons, P.P.; Turnbull, J.F.; Dawson, K.A.; Crumlish, M. Effects of low-level dietary microalgae supplementation on the distal intestinal microbiome of farmed rainbow trout Oncorhynchus mykiss (Walbaum). Aquac. Res. 2017, 48, 2438–2452. [Google Scholar] [CrossRef]
- Sullam, K.E.; Essinger, S.D.; Lozupone, C.A.; O’Connor, M.P.; Rosen, G.L.; Knight, R.; Kilham, S.S.; Russell, J.A. Environmental and ecological factors that shape the gut bacterial communities of fish: A meta-analysis. Mol. Ecol. 2012, 21, 3363–3378. [Google Scholar] [CrossRef] [Green Version]
- Roeselers, G.; Mittge, E.K.; Stephens, W.Z.; Parichy, D.M.; Cavanaugh, C.M.; Guillemin, K.; Rawls, J.F. Evidence for a core gut microbiota in the zebrafish. ISME J. 2011, 5, 1595–1608. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, V.T.; Smith, K.F.; Melvin, D.W.; Amaral-Zettler, L.A. Community assembly of a euryhaline fish microbiome during salinity acclimation. Mol. Ecol. 2015, 24, 2537–2550. [Google Scholar] [CrossRef]
- Zhang, M.; Sun, Y.; Liu, Y.; Qiao, F.; Chen, L.; Liu, W.-T.; Du, Z.; Li, E. Response of gut microbiota to salinity change in two euryhaline aquatic animals with reverse salinity preference. Aquaculture 2016, 454, 72–80. [Google Scholar] [CrossRef]
- Ray, C. Characterization of the Gut and Skin Microbiomes of Wild-Caught Fishes from Lake Guntersville, , Alabama. Master’s Thesis, Auburn University, Auburn, AL, USA, 2016. [Google Scholar]
- Zarkasi, K.Z.; Abell, G.C.J.; Taylor, R.S.; Neuman, C.; Hatje, E.; Tamplin, M.L.; Katouli, M.; Bowman, J.P. Pyrosequencing-based characterization of gastrointestinal bacteria of Atlantic salmon (Salmo salar L.) within a commercial mariculture system. J. Appl. Microbiol. 2014, 117, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.C.R.; Snowberg, L.K.; Gregory Caporaso, J.; Knight, R.; Bolnick, D.I. Dietary input of microbes and host genetic variation shape among-population differences in stickleback gut microbiota. ISME J. 2015, 9, 2515–2526. [Google Scholar] [CrossRef]
- Ye, L.; Amberg, J.; Chapman, D.; Gaikowski, M.; Liu, W.-T. Fish gut microbiota analysis differentiates physiology and behavior of invasive Asian carp and indigenous American fish. ISME J. 2014, 8, 541–551. [Google Scholar] [CrossRef] [Green Version]
- Ringø, E.; Zhou, Z.; Vecino, J.; Wadsworth, S.; Romero, J.; Krogdahl, Å.; Olsen, R.E.; Dimitroglou, A.; Foey, A.; Davies, S.; et al. Effect of dietary components on the gut microbiota of aquatic animals. A never-ending story? Aquac. Nutr. 2016, 22, 219–282. [Google Scholar] [CrossRef] [Green Version]
- Romero, J.; Ringø, E.; Merrifield, D.L. The Gut Microbiota of Fish. In Aquaculture Nutrition: Gut Health, Probiotics, and Prebiotics; Merrifield, D., Ringø, E., Eds.; John Wiley & Sons Inc.: Chichester, UK, 2014; pp. 75–100. ISBN 9781118897263. [Google Scholar]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost microbiomes: The state of the art in their characterization, manipulation and importance in aquaculture and fisheries. Front. Microbiol. 2014, 5, 207. [Google Scholar] [CrossRef] [Green Version]
- Palm, H.W.; Knaus, U.; Wasenitz, B.; Bischoff, A.A.; Strauch, S.M. Proportional up scaling of African catfish (Clarias gariepinus Burchell, 1822) commercial recirculating aquaculture systems disproportionally affects nutrient dynamics. Aquaculture 2018, 491, 155–168. [Google Scholar] [CrossRef]
- Bovendeur, J.; Eding, E.H.; Henken, A.M. Design and performance of a water recirculation system for high-density culture of the African catfish, Clarias gariepinus (Burchell 1822). Aquaculture 1987, 63, 329–353. [Google Scholar] [CrossRef]
- Rosenau, S.; Oertel, E.; Dietz, C.; Wessels, S.; Tetens, J.; Mörlein, D.; Ciulu, M. Total Replacement of Fishmeal by Spirulina (Arthrospira platensis) and its Effect on Growth Performance and Product Quality of African catfish (Clarias gariepinus). Sustainability. submitted.
- Dietz, C.; Sünder, A.; Liebert, F. Does Genetic Background of Rainbow Trout Impact on Growth and Feed Utilization Following Fishmeal Substitution by Partly Defatted Insect Meal (Hermetia illucens) or Microalgae Powder (Arthrospira platensis)?: 74rd Conference 3rd–5th March 2020 in Göttingen; DLG-Verlag: Frankfurt, Germany, 2020; ISBN 978-3-7690-4113-2. [Google Scholar]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: Cham, Germany, 2016; ISBN 978-0-387-98140-6. [Google Scholar]
- Yukgehnaish, K.; Kumar, P.; Sivachandran, P.; Marimuthu, K.; Arshad, A.; Paray, B.A.; Arockiaraj, J. Gut microbiota metagenomics in aquaculture: Factors influencing gut microbiome and its physiological role in fish. Rev. Aquac. 2020, 12, 1903–1927. [Google Scholar] [CrossRef]
- Eichmiller, J.J.; Hamilton, M.J.; Staley, C.; Sadowsky, M.J.; Sorensen, P.W. Environment shapes the fecal microbiome of invasive carp species. Microbiome 2016, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Chen, J.; Meng, S.; Song, C.; Qiu, L.; Hu, G.; Xu, P. Characterization of microbial communities in intensive GIFT tilapia (Oreochromis niloticus) pond systems during the peak period of breeding. Aquac. Res. 2017, 48, 459–472. [Google Scholar] [CrossRef]
- Minich, J.J.; Zhu, Q.; Xu, Z.Z.; Amir, A.; Ngochera, M.; Simwaka, M.; Allen, E.E.; Zidana, H.; Knight, R. Microbial effects of livestock manure fertilization on freshwater aquaculture ponds rearing tilapia (Oreochromis shiranus) and North African catfish (Clarias gariepinus). Microbiologyopen 2018, 7, e00716. [Google Scholar] [CrossRef]
- Bledsoe, J.W.; Peterson, B.C.; Swanson, K.S.; Small, B.C. Ontogenetic Characterization of the Intestinal Microbiota of Channel Catfish through 16S rRNA Gene Sequencing Reveals Insights on Temporal Shifts and the Influence of Environmental Microbes. PLoS ONE 2016, 11, e0166379. [Google Scholar] [CrossRef]
- Tsuchiya, C.; Sakata, T.; Sugita, H. Novel ecological niche of Cetobacterium somerae, an anaerobic bacterium in the intestinal tracts of freshwater fish. Lett. Appl. Microbiol. 2008, 46, 43–48. [Google Scholar] [CrossRef]
- Gerritsen, J.; Hornung, B.; Ritari, J.; Paulin, L.; Rijkers, G.T.; Schaap, P.J.; de Vos, W.M.; Smidt, H. A Comparative and Functional Genomics Analysis of the Genus Romboutsia Provides Insight into Adaptation to an Intestinal Lifestyle. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.-J.; Qiang, J.; Tao, Y.-F.; Ngoepe, T.K.; Bao, J.-W.; Chen, D.-J.; Xu, P. Physiological and gut microbiome changes associated with low dietary protein level in genetically improved farmed tilapia (GIFT, Oreochromis niloticus) determined by 16S rRNA sequence analysis. Microbiologyopen 2020, 9, e1000. [Google Scholar] [CrossRef] [Green Version]
- Michail, S.; Lin, M.; Frey, M.R.; Fanter, R.; Paliy, O.; Hilbush, B.; Reo, N.V. Altered gut microbial energy and metabolism in children with non-alcoholic fatty liver disease. FEMS Microbiol. Ecol. 2015, 91, 1–9. [Google Scholar] [CrossRef]
- Ren, N.; Xing, D.; Rittmann, B.E.; Zhao, L.; Xie, T.; Zhao, X. Microbial community structure of ethanol type fermentation in bio-hydrogen production. Environ. Microbiol. 2007, 9, 1112–1125. [Google Scholar] [CrossRef]
- Ahmad, M.; Aggarwal, M.; Ahmed, A. Bloody diarrhea caused by Plesiomonas shigelloides proctitis in a human immunodeficiency virus-infected patient. Clin. Infect. Dis. 1998, 27, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenden, R.A.; Miller, M.A.; Janda, M.J. Clinical Disease Spectrum and Pathogenic Factors Associated with Plesiomonas shigelloides Infections in Humans. Rev. Infect. Dis. 1988, 10, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Sugita, H.; Shibuya, K.; Shimooka, H.; Deguchi, Y. Antibacterial abilities of intestinal bacteria in freshwater cultured fish. Aquaculture 1996, 145, 195–203. [Google Scholar] [CrossRef]
- Suphoronski, S.A.; Chideroli, R.T.; Facimoto, C.T.; Mainardi, R.M.; Souza, F.P.; Lopera-Barrero, N.M.; Jesus, G.F.A.; Martins, M.L.; Di Santis, G.W.; de Oliveira, A.; et al. Effects of a phytogenic, alone and associated with potassium diformate, on tilapia growth, immunity, gut microbiome and resistance against francisellosis. Sci. Rep. 2019, 9, 6045. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wu, S.; Wirth, S.; Hao, Y.; Wang, W.; Zou, H.; Li, W.; Wang, G. Diversity and activity of cellulolytic bacteria, isolated from the gut contents of grass carp (Ctenopharyngodon idellus) (Valenciennes) fed on Sudan grass (Sorghum sudanense) or artificial feedstuffs. Aquac. Res. 2016, 47, 153–164. [Google Scholar] [CrossRef]
- Karachle, P.K.; Stergiou, K.I. Intestine Morphometrics of Fishes: A Compilation and Analysis of Bibliographic Data. Acta Ichthyol. Piscat. 2010, 40, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Escalas, A.; Auguet, J.-C.; Avouac, A.; Seguin, R.; Gradel, A.; Borrossi, L.; Villéger, S. Ecological Specialization Within a Carnivorous Fish Family Is Supported by a Herbivorous Microbiome Shaped by a Combination of Gut Traits and Specific Diet. Front. Mar. Sci. 2021, 8, 91. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Wang, G.; Angert, E.R.; Wang, W.; Li, W.; Zou, H. Composition, diversity, and origin of the bacterial community in grass carp intestine. PLoS ONE 2012, 7, e30440. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zheng, Z.; Cong-xin, X.; Bo, H.; Chao-yuan, W.; Gang, H. Isolation of cellulose—producing microbes from the intestine of grass carp (Ctenopharyngodon idellus). In Chinese Fishes; Noakes, D.L.G., Romero, A., Zhao, Y., Zhou, Y., Eds.; Springer Science+Business Media B.V: Dordrecht, The Netherlands, 2009; pp. 131–135. ISBN 978-90-481-3457-1. [Google Scholar]
- Nayak, S.K. Role of gastrointestinal microbiota in fish. Aquac. Res. 2010, 41, 1553–1573. [Google Scholar] [CrossRef]
- Santigosa, E.; Sánchez, J.; Médale, F.; Kaushik, S.; Pérez-Sánchez, J.; Gallardo, M.A. Modifications of digestive enzymes in trout (Oncorhynchus mykiss) and sea bream (Sparus aurata) in response to dietary fish meal replacement by plant protein sources. Aquaculture 2008, 282, 68–74. [Google Scholar] [CrossRef]
- Ogunji, J.O.; Awoke, J. Effect of environmental regulated water temperature variations on survival, growth performance and haematology of African catfish, Clarias gariepinus. Our Nat. 2017, 15, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Peteri, A.; Nandi, S.; Chowdhury, S.N. Manual on Seed Production of African Catfish (Clarias gariepinus); FAO: Rome, Italy, 1992. [Google Scholar]
- Schram, E.; Roques, J.A.C.; Abbink, W.; Yokohama, Y.; Spanings, T.; de Vries, P.; Bierman, S.; van de Vis, H.; Flik, G. The impact of elevated water nitrate concentration on physiology, growth and feed intake of African catfish Clarias gariepinus (Burchell 1822). Aquac. Res. 2014, 45, 1499–1511. [Google Scholar] [CrossRef]
- Schram, E.; Roques, J.A.; Abbink, W.; Spanings, T.; de Vries, P.; Bierman, S.; de van Vis, H.; Flik, G. The impact of elevated water ammonia concentration on physiology, growth and feed intake of African catfish (Clarias gariepinus). Aquaculture 2010, 306, 108–115. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
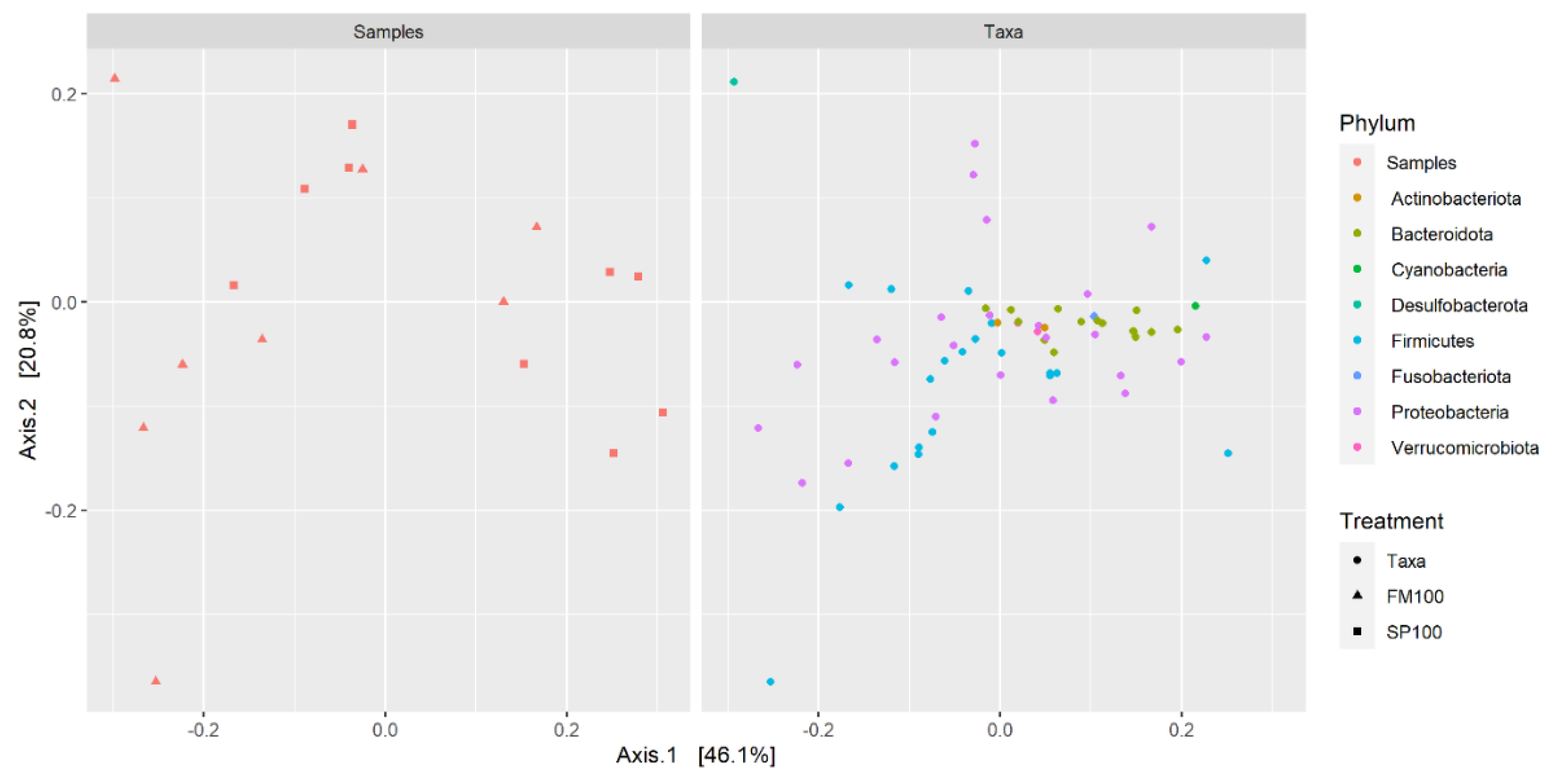{kind=link}
| Ingredient (% Dry Matter) | FM100 | SP100 |
|---|---|---|
| Fishmeal 1 | 20.00 | 0.00 |
| Spirulina 2 | 0.00 | 20.00 |
| Fish oil | 10.70 | 10.70 |
| Wheat meal | 14.00 | 12.50 |
| Wheat gluten | 20.00 | 21.50 |
| Soy protein concentrate 3 | 20.00 | 20.00 |
| Rapeseed oil | 10.70 | 10.70 |
| Vit./Min. Premix | 1.00 | 1.00 |
| CaHPO4 | 1.00 | 1.00 |
| CMC (Binder) | 1.29 | 1.08 |
| TiO2 (Marker) | 0.50 | 0.50 |
| Fe3O4–black (Dye) | 0.07 | 0.07 |
| L-Lysin (HCL-Lys, 78% Lys) | 0.70 | 0.90 |
| D,L-Methionine | 0.01 | 0.04 |
| L-Tryptophan | 0.03 | 0.01 |
| Approximate Composition (%) | FM100 | SP100 |
|---|---|---|
| Dry matter | 94.6 | 94.0 |
| Crude protein (N × 6.25) | 45.4 | 45.7 |
| Crude lipids | 24.6 | 23.9 |
| N-free extracts | 17.5 | 19.0 |
| Crude ash | 7.1 | 5.4 |
| Gross energy [MJ/kg] | 23.4 | 23.5 |
| Digestible energy [MJ/kg] | 20.0 | 20.0 |
| Essential amino acids 1 | 26.6 | 26.8 |
| Genus | FM100 | SP100 |
|---|---|---|
| Cetobacterium | 54.98 ± 20.43 | 58.90 ± 13.34 |
| Romboutsia | 20.30 ± 14.88 | 8.14 ± 3.26 |
| Macellibacteroides | 4.15 ± 4.10 | 5.05 ± 2.50 |
| Plesiomonas | 2.95 ± 2.47 | 5.66 ± 2.95 |
| Bacteroides | 1.50 ± 0.61 | 8.82 ± 5.72 |
| [Anaerorhabdus] furcosa group | 3.44 ± 2.96 | 1.88 ± 1.92 |
| [Barnesiellaceae] uncultured | 1.30 ± 1.15 | 2.17 ± 1.72 |
| Clostridium sensu stricto 1 | 2.39 ± 1.94 | 0.72 ± 0.38 |
| Turicibacter | 2.25 ± 2.30 | 0.21 ± 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosenau, S.; Oertel, E.; Mott, A.C.; Tetens, J. The Effect of a Total Fishmeal Replacement by Arthrospira platensis on the Microbiome of African Catfish (Clarias gariepinus). Life 2021, 11, 558. https://doi.org/10.3390/life11060558
Rosenau S, Oertel E, Mott AC, Tetens J. The Effect of a Total Fishmeal Replacement by Arthrospira platensis on the Microbiome of African Catfish (Clarias gariepinus). Life. 2021; 11(6):558. https://doi.org/10.3390/life11060558
Chicago/Turabian StyleRosenau, Simon, Elisa Oertel, Alexander Charles Mott, and Jens Tetens. 2021. "The Effect of a Total Fishmeal Replacement by Arthrospira platensis on the Microbiome of African Catfish (Clarias gariepinus)" Life 11, no. 6: 558. https://doi.org/10.3390/life11060558