
Integrated Biomarker Response for Environmental Assessment Using the Gastropod Phorcus turbinatus along the Northern and the Northeastern Coasts of Tunisia

Abstract
:1. Introduction
2. Material and Methods
2.1. Samples Grinding
2.2. Determination of Total Protein
2.3. Catalase
2.4. Glutathione-S-Transferase
2.5. Acethylcholinesterase
2.6. Integrated Biological Response IBR
3. Results
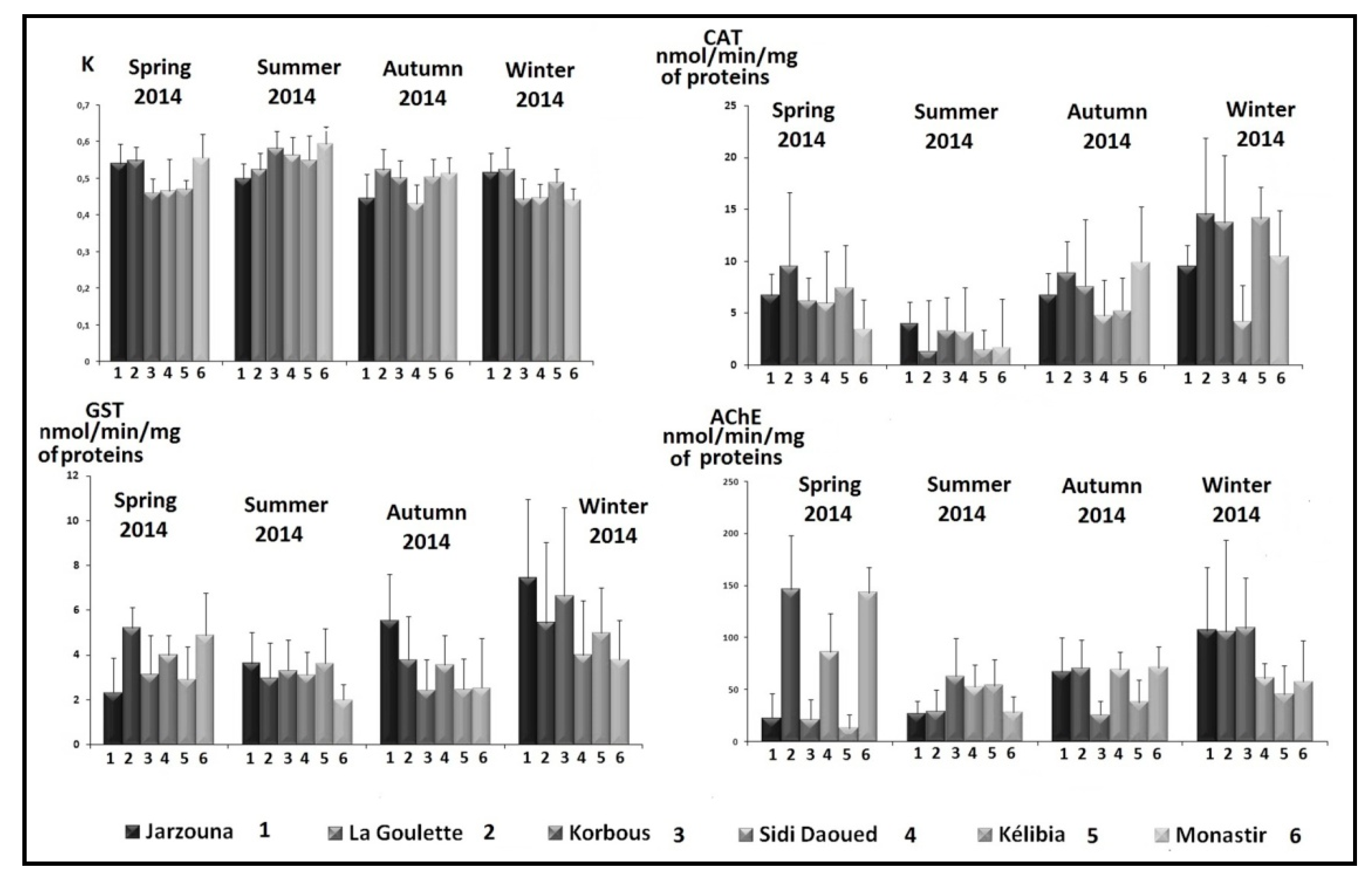
3.1. Fulton Condition Factor (K)
3.2. Catalase
3.3. Glutathione S-Transferase (GST)
3.4. Acetylcholinesterase (AChE)
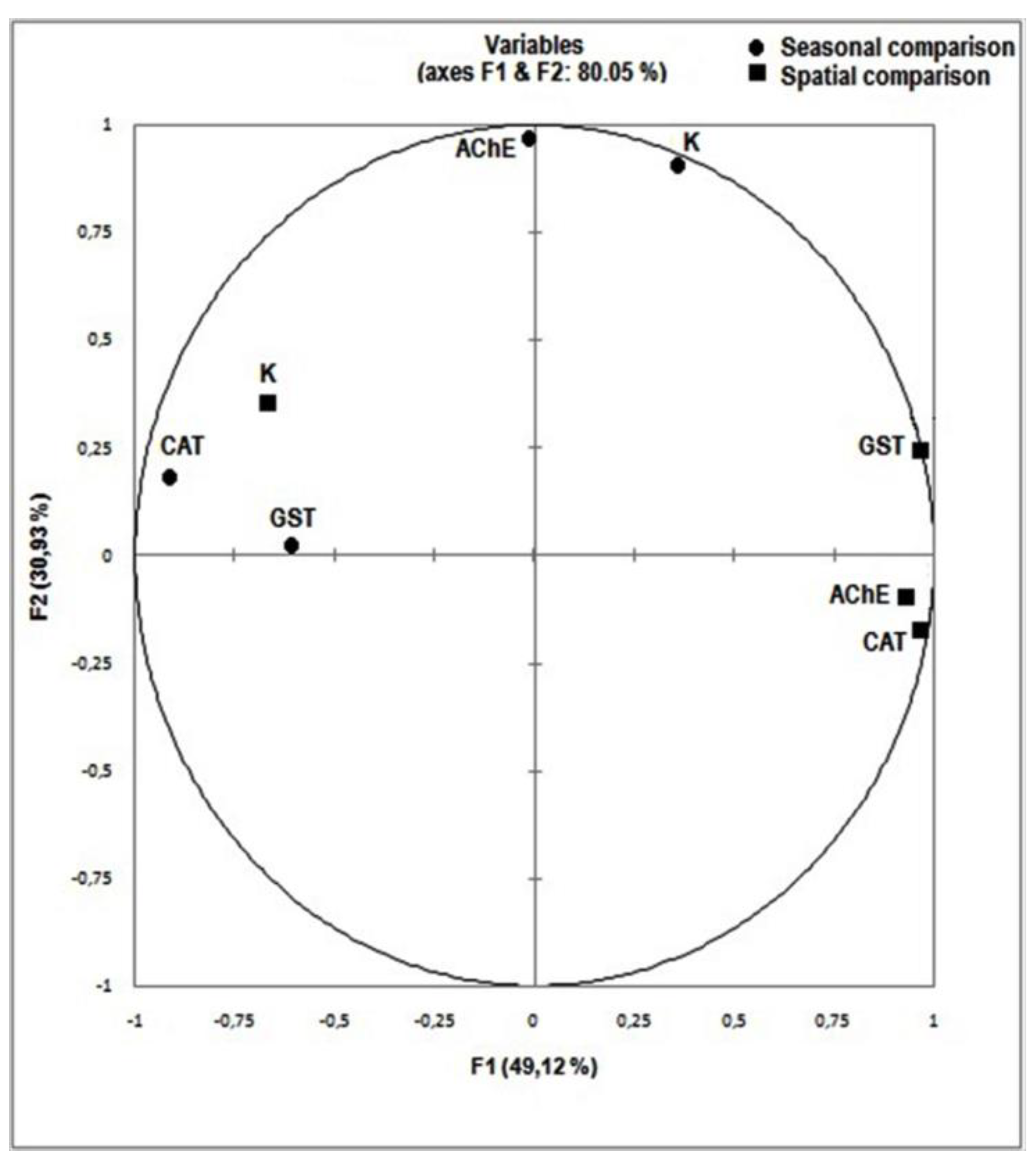
3.5. Integrated Biological Response IB
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Coll, M.; Piroddi, C.; Albouy, C.; Ben Rais-Lasram, F.; Cheung, W.W.L.; Christensen, V.; Karpouzi, V.S.; Guilhaumon, F.; Mouillot, D.; Paleczny, M.; et al. The Mediterranean Sea under siege: Spatial overlap between marine biodiversity, cumulative threats and marine reserves: The Mediterranean Sea under siege. Glob. Ecol. Biogeogr. 2012, 21, 465–480. [Google Scholar] [CrossRef]
- Zorita, I.; Apraiz, I.; Ortiz-Zarragoitia, M.; Orbea, A.; Cancio, I.; Soto, M.; Marigómez, I.; Cajaraville, M.P. Assessment of biological effects of environmental pollution along the NW Mediterranean Sea using mussels as sentinel organisms. Environ. Pollut. 2007, 148, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Serafim, A.; Company, R.; Lopes, B.; Fonseca, V.F.; França, S.; Vasconcelos, R.P.; Bebianno, M.J.; Cabral, H.N. Application of an integrated biomarker response index (IBR) to assess temporal variation of environmental quality in two Portuguese aquatic systems. Ecol. Indic. 2012, 19, 215–225. [Google Scholar] [CrossRef]
- Eduljee, G.H. Budget and Source Inventories. In Persistent Organic Pollutants; Springer: Boston, MA, USA, 2001; p. 128. [Google Scholar]
- Benlahcen, K.T.; Chaoui, A.; Budzinski, H.; Bellocq, J.; Garrigues, P. Distribution and sources of polycyclic aromatic hydrocarbons in some Mediterranean coastal sediments. Mar. Pollut. Bull. 1997, 34, 298–305. [Google Scholar] [CrossRef]
- Galgani, F.; Martínez-Gómez, C.; Giovanardi, F.; Romanelli, G.; Caixach, J.; Cento, A.; Scarpato, A.; BenBrahim, S.; Messaoudi, S.; Deudero, S.; et al. Assessment of polycyclic aromatic hydrocarbon concentrations in mussels (Mytilus galloprovincialis) from the Western basin of the Mediterranean Sea. Environ. Monit. Assess. 2011, 172, 301–317. [Google Scholar] [CrossRef] [Green Version]
- Ben Said, O.; Goñi-Urriza, M.S.; El Bour, M.; Dellali, M.; Aissa, P.; Duran, R. Characterization of aerobic polycyclic aromatic hydrocarbon-degrading bacteria from Bizerte lagoon sediments, Tunisia. J. Appl. Microbiol. 2008, 104, 987–997. [Google Scholar] [CrossRef]
- Nouira, T.; Tagorti, M.A.; Budzinski, H.; Etchebert, H.; Boussetta, H. Polycyclic aromatic hydrocarbons (PAHs) in surface sediments of Monastir Bay (Tunisia, Central Mediterranean): Distribution, origin and seasonal variations. Int. J. Environ. Anal. Chem. 2013, 93, 1470–1483. [Google Scholar] [CrossRef]
- Boulajfene, W.; Boukhicha, J.; Deidun, A.; Berto, D.; Romeo, T.; Ben Hassine, O.K.; Tlig-Zouari, S. Biomonitoring of the environmental contamination by organotins in the Gulf of Tunis: Occurrence of imposex in Stramonita haemastoma (Linnaeus, 1767). Mar. Freshw. Res. 2015, 66, 778–785. [Google Scholar] [CrossRef]
- Boulajfene, W.; Strogyloudi, E.; Catsiki, V.A.; El Mlayah, A.; Tlig-Zouari, S. Bio-monitoring of metal impact on metallothioneins levels in the gastropod Phorcus turbinatus (Born, 1778) in the northeastern and the eastern coasts of Tunisia. Mar. Pollut. Bull. 2017, 120, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Trabelsi, S.; Driss, M.R. Polycyclic aromatic hydrocarbons in superficial coastal sediments from Bizerte Lagoon, Tunisia. Mar. Pollut. Bull. 2005, 50, 344–348. [Google Scholar] [CrossRef]
- Zrafi-Nouira, I.; Khedir-Ghenim, Z.; Zrafi, F.; Bahri, R.; Cheraeif, I.; Rouabhia, M.; Saidane-Mosbahi, D. Hydrocarbon Pollution in the Sediment from the Jarzouna-Bizerte Coastal Area of Tunisia (Mediterranean Sea). Bull. Environ. Contam. Toxicol. 2008, 80, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Ben Ameur, W.; Trabelsi, S.; Driss, M.R. Polycyclic Aromatic Hydrocarbons in Superficial Sediments from Ghar El Melh Lagoon, Tunisia. Bull. Environ. Contam. Toxicol. 2010, 85, 184–189. [Google Scholar] [CrossRef]
- Mzoughi, N.; Chouba, L. Distribution of trace metals, aliphatic hydrocarbons and polycyclic aromatic hydrocarbons in sediment cores from the Sicily Channel and the Gulf of Tunis (south-western Mediterranean Sea). Environ. Technol. 2011, 32, 43–54. [Google Scholar] [CrossRef]
- Zrafi, I.; Hizem, L.; Chalghmi, H.; Ghrabi, A.; Rouabhia, M.; Saidane-Mosbahi, D. Aliphatic and Aromatic Biomarkers for Petroleum Hydrocarbon Investigation in Marine Sediment. J. Pet. Sci. Eng. 2013, 2, 145–155. [Google Scholar] [CrossRef]
- Zaghden, H.; Tedetti, M.; Sayadi, S.; Serbaji, M.M.; Elleuch, B.; Saliot, A. Origin and distribution of hydrocarbons and organic matter in the surficial sediments of the Sfax-Kerkennah channel (Tunisia, Southern Mediterranean Sea). Mar. Pollut. Bull. 2017, 117, 414–428. [Google Scholar] [CrossRef]
- Melila, M.; Poutouli, W.; Amouzou, K.S.; Gado, T.; Tchao, M.; Doh, A. Évaluation de l’impact du rejet des déchets phosphates dans la mer sur la biodiversité marine dans trois localités côtières au Togo à partir des biomarqueurs du stress oxydatif chez Sphyraena barracuda (HECKEL, 1843). Int. J. Biol. Chem. Sci. 2012, 6, 820–831. [Google Scholar] [CrossRef] [Green Version]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Dellali, M.; Romeo, M.; Aissa, P. Suivi annuel de l’activité catalase chez des moules et des palourdes originaires de la lagune de Bizerte. Oceanol. Acta 2001, 24, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Yİldİrİm, N.C.; Benzer, F.; Danabas, D. Evaluation of environmental pollution at Munzur River of Tunceli applying oxidative stress biomarkers in Capoeta trutta (Heckel, 1843). J. Anim. Plant. Sci. 2011, 21, 66–71. [Google Scholar]
- Farombi, E.O.; Adelowo, O.A.; Ajimoko, Y.R. Biomarkers of oxidative stress and heavy metal levels as indicators of environmental pollution in African cat fish (Clarias gariepinus) from Nigeria Ogun River. Int. J. Environ. Res. Public Health 2007, 4, 158–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velkova-Jordanoska, L.; Kostoski, G.; Jordanoska, B. Antioxidative enzymes in fish as biochemical indicators of aquatic pollution. Bulg. J. Agric. Sci. 2008, 14, 235–237. [Google Scholar]
- Angel, D.L.; Fiedler, U.; Eden, N.; Kress, N.; Adelung, D.; Herut, B. Catalase activity in macro-and microorganisms as an indicator of biotic stress in coastal waters of the eastern Mediterranean Sea. Helgol. Mar. Res. 1999, 53, 209–218. [Google Scholar]
- Hoarau, P.; Gnassia-Barelli, M.; Romeo, M.; Girard, J.P. Differential induction of glutathione S-transferases in the clam Ruditapes decussatus exposed to organic compounds. Environ. Toxicol. Chem. 2001, 20, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Hoarau, P.; Garello, G.; Gnassia-Barelli, M.; Roméo, M.; Girard, J.P. Effect of three xenobiotic compounds on Glutathione S-Transferase in the clam Ruditapes decussatus. Aquat. Toxicol. 2004, 68, 87–94. [Google Scholar] [CrossRef]
- Binelli, A.; Ricciardi, F.; Riva, C.; Provini, A. New evidences for old biomarkers: Effects of several xenobiotics on EROD and AChE activities in Zebra mussel (Dreissena polymorpha). Chemosphere 2006, 62, 510–519. [Google Scholar] [CrossRef]
- Galgani, F.; Bocquene, G. Semi-automated colorimetric and enzymatic assays for aquatic organisms using microplate readers. Water Res. 1991, 25, 147–150. [Google Scholar] [CrossRef]
- Belhaouari, B.; Rouane-Hacene, O.; Bouhadiba, S.; Boutiba, Z. Utilisation d’un Gastéropode marin Osilinus turbinatus en biosurveillance marine: Application aux métaux lourds du littoral algérien occidental. J. Sci. Hal. Aquat. 2011, 3, 89–96. [Google Scholar]
- Boucetta, S.; Beldi, H.; Draredja, B. Seasonal Variation of Heavy Metals in Phorcus (Osilinus) turbinatus (Gastropod, Trochidae) in the Eastern Algerian Coast. Glob. Vet. 2016, 17, 25–41. [Google Scholar]
- Bode, A.; Lombas, I.; Anadon, N. Preliminary studies on the reproduction and population dynamics of Monodonta lineata and Gibbula umbilicalis (Mollusca, Gastropoda) on the central coast of Asturias (N. Spain). Hydrobiologia 1986, 142, 31–39. [Google Scholar] [CrossRef]
- Garwood, P.R.; Kendall, M.A. The Reproductive Cycles of Monodonta Lineata and Gibbula Umbilicalis on the Coast of Mid-Wales. J. Mar. Biol. Assoc. United Kingd. 1985, 65, 993–1008. [Google Scholar] [CrossRef]
- Fulton, T.W. The Sovereignty of the Sea: An Historical Account of the Claims of England to the Dominion of the British Seas, and of the Evolution of the Territorial Waters, with Special Reference to the Rights of Fishing and the Naval Salute; Blackwood and Sons: Edinburgh, UK, 1911. [Google Scholar]
- Annabi, A.; Dhouib Bini, I.; Lasram, M.; Gharbi, N.; El Fazâa, S. Oxidative stress, biochemical alterations, and hyperlipidemia in female rats induced by lead chronic toxicity during puberty and post puberty periods. Iran. J. Basic Med. Sci. 2015, 18, 1034–1043. [Google Scholar]
- Gornall, A.G.; Bardawill, C.J.; David, M.M. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem. 1949, 177, 751–766. [Google Scholar] [CrossRef]
- Cohen, G.; Dembiec, D.; Marcus, J. Measurement of catalase activity in tissue extracts. Anal. Biochem. 1970, 34, 30–38. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Arzate-Cárdenas, M.A.; Martínez-Jerónimo, F. Age-altered susceptibility in hexavalent chromium-exposed Daphnia schodleri (Anomopoda: Daphniidae): Integrated Biomarker Response implementation. Aquat. Toxicol. 2011, 105, 528–534. [Google Scholar] [CrossRef]
- Damiens, G.; His, E.; Gnassia-Barelli, M.; Quiniou, F.; Roméo, M. Evaluation of biomarkers in oyster larvae in natural and polluted conditions. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2004, 138, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Jebali, J.; Ben-Khedher, S.; Ghedira, J.; Kamel, N.; Boussetta, H. Integrated assessment of biochemical responses in Mediterranean crab (Carcinus maenas) collected from Monastir Bay, Tunisia. J. Environ. Sci. 2011, 23, 1714–1720. [Google Scholar] [CrossRef]
- Kim, W.K.; Lee, S.K.; Jung, J. Integrated assessment of biomarker responses in common carp (Cyprinus carpio) exposed to perfluorinated organic compounds. J. Hazard. Mater. 2010, 180, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.; Ahmad, I.; Maria, V.L.; Pacheco, M.; Santos, M.A. Monitoring pollution of coastal lagoon using Liza aurata kidney oxidative stress and genetic endpoints: An integrated biomarker approach. Ecotoxicology 2010, 19, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lu, G.H.; Song, W.T.; Xu, S.; Wang, P.F. Integrated biomarker response index for the assessment of environmental stress of the Yangtze River (Nanjing section). Fish. Physiol. Biochem. 2010, 36, 1069–1078. [Google Scholar] [CrossRef]
- Sanchez, W.; Burgeot, T.; Porcher, J.M. A novel “Integrated Biomarker Response” calculation based on reference deviation concept. Environ. Sci. Pollut. Res. 2013, 20, 2721–2725. [Google Scholar] [CrossRef]
- Richardson, B.J.; Mak, E.; De Luca-Abbott, S.B.; Martin, M.; McClellan, K.; Lam, P.K.S. Antioxidant responses to polycyclic aromatic hydrocarbons and organochlorine pesticides in green-lipped mussels (Perna viridis): Do mussels “integrate” biomarker responses? Mar. Pollut. Bull. 2008, 57, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Van der Oost, R.; Beyerb, J.; Vermeulen, N. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Boudouresque, C.F. Excursion au Cap-Croisette (Marseille): Le Milieu Marin, 13th ed.; GIS Posidonie Publishers: Marseilles, France, 2013; p. 152. [Google Scholar]
- Boulajfene, W.; Strogyloudi, E.; Lasram, M.; El Mlayah, A.; Vassiliki-Angelique, C.; Zouari-Tlig, S. Biological and biochemical assessment in Phorcus articulatus (Lamarck 1822): Contamination and seasonal effect. Environ. Monit. Assess. 2019, 191, 555. [Google Scholar] [CrossRef] [PubMed]
- Bejaoui, S.; Boussoufa, D.; Tir, M.; Haouas-gharsallah, I.; Boudawara, T.; Ghram, A.; El-Cafsi, M.; Soudani, N. DNA damage and oxidative stress in digestive gland of Venerupis decussata collected from two contrasting habitats in the southern Tunisian coast: Biochemical and histopathological studies. Cah. Biol. Mar. 2017, 58, 123–135. [Google Scholar]
- Rodríguez-Ariza, A.; Peinado, J.; Pueyo, C.; López-Barea, J. Biochemical Indicators of Oxidative Stress in Fish from Polluted Littoral Areas. Can. Spec. Publ. Fish. Aquat. Sci. 1993, 50, 2568–2573. [Google Scholar] [CrossRef]
- Barhoumi, B.; Menach, K.L.; Clérandeau, C.; Ameur, W.B.; Budzinski, H.; Driss, M.R.; Cachot, J. Assessment of pollution in the Bizerte lagoon (Tunisia) by the combined use of chemical and biochemical markers in mussels, Mytilus galloprovincialis. Mar. Pollut. Bull. 2014, 84, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumar, P.K.; Casillas, E.; Varanasi, U. Cytochemical responses in the digestive tissue of Mytilus edulis complex exposed to microencapsulated PAHs or PCBs. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1997, 118, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Wenning, R.J.; Di Giulio, R.T.; Gallagher, E.P. Oxidant-mediated biochemical effects of paraquat in the ribbed mussel, Geukensia demissa. Aquat. Toxicol. 1988, 12, 157–170. [Google Scholar] [CrossRef]
- Livingstone, D.R.; Martinez, P.G.; Michel, X.; Narbonne, J.F.; O’Hara, S.; Ribera, D.; Winston, G.W. Oxyradical Production as a Pollution-Mediated Mechanism of Toxicity in the Common Mussel, Mytilus edulis L., and Other Molluscs. Funct. Ecol. 1990, 4, 415–424. [Google Scholar] [CrossRef]
- Stanic, B.; Andric, N.; Zoric, S.; Grubor-Lajsic, G.; Kovacevic, R. Assessing pollution in the Danube River near Novi Sad (Serbia) using several biomarkers in sterlet (Acipenser ruthenus L.). Ecotoxicol. Environ. Saf. 2006, 65, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Jebali, J.; Banni, M.; Almeida, E.A.D.; Boussetta, H. Oxidative DNA damage levels and catalase activity in the clam Ruditapes decussatus as pollution biomarkers of Tunisian marine environment. Environ. Monit. Assess. 2007, 124, 195–200. [Google Scholar] [CrossRef]
- Kamel, N.; Jebali, J.; Banni, M.; Ben Khedher, S.; Chouba, L.; Boussetta, H. Biochemical responses and metals levels in Ruditapes decussatus after exposure to treated municipal effluents. Ecotoxicol. Environ. Saf. 2012, 82, 40–46. [Google Scholar] [CrossRef]
- Kádár, E.; Costa, V.; Santos, R.S. Distribution of micro-essential (Fe, Cu, Zn) and toxic (Hg) metals in tissues of two nutritionally distinct hydrothermal shrimps. Sci. Total Environ. 2006, 358, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Srarfi, F.; Slim-Shimi, N. Evaluation simplifiée des risques: Une approche qui s’applique au nord de Tunisie. Larhyss J. 2004, 186, 185–196. [Google Scholar]
- Kaaya, A.; Najimi, S.; Ribera, D.; Narbonne, J.F.; Moukrim, A. Characterization of glutathione S-transferases (GST) activities in Perna perna and Mytilus galloprovincialis used as a biomarker of pollution in the Agadir Marine Bay (South of Morocco). Bull. Environ. Contam. Toxicol. 1999, 62, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Damiens, G.; Gnassia-Barelli, M.; Loquès, F.; Roméo, M.; Salbert, V. Integrated biomarker response index as a useful tool for environmental assessment evaluated using transplanted mussels. Chemosphere 2007, 66, 574–583. [Google Scholar] [CrossRef]
- Aouini, F.; Boussoufa, D.; Ghribi, F.; Bejaoui, S.; Navarro, J.; El Cafsi, M. Short term exposure to permethrin alters lipid classes and fatty acid composition of the unfed clam Donax trunculus from Tunisian coast (Southern Mediterranean). Cah. Biol. Mar. 2017, 58, 163–172. [Google Scholar]
- Matozzo, V.; Tomei, A.; Marin, M.G. Acetylcholinesterase as a biomarker of exposure to neurotoxic compounds in the clam Tapes philippinarum from the Lagoon of Venice. Mar. Pollut. Bull. 2005, 50, 1686–1693. [Google Scholar] [CrossRef]
- Chalghmi, H.; Bourdineaud, J.P.; Haouas, Z.; Gourves, P.Y.; Zrafi, I.; Saidane-Mosbahi, D. Transcriptomic, Biochemical, and Histopathological Responses of the Clam Ruditapes decussatus from a Metal-Contaminated Tunis Lagoon. Arch. Environ. Contam. Toxicol. 2016, 70, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Beliaeff, B.; Burgeot, T. Integrated biomarker response: A useful tool for ecological risk assessment. Environ. Toxicol. Chem. 2002, 21, 1316–1322. [Google Scholar] [CrossRef] [PubMed]
- Broeg, K.; Westernhagen, H.; Zander, S.; Körting, W.; Koehler, A. The “bioeffect assessment index” (BAI). Mar. Pollut. Bull. 2005, 50, 495–503. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stations | Spring 2014 | Summer 2014 | Autumn 2014 | Winter 2015 |
|---|---|---|---|---|
| Jarzouna | 2.1235 | 7.1014 | 4.2104 | 3.4296 |
| La Goulette | 5.8597 | 3.1587 | 4.0036 | 3.9436 |
| Sidi Daoued | 2.6256 | 4.0106 | 4.4448 | 7.1307 |
| Kélibia | 2.0367 | 3.5005 | 2.5134 | 5.6174 |
| Monastir | 5.5657 | 4.0106 | 2.4611 | 5.7923 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boulajfene, W.; Lasram, M.; Zouari-Tlig, S. Integrated Biomarker Response for Environmental Assessment Using the Gastropod Phorcus turbinatus along the Northern and the Northeastern Coasts of Tunisia. Life 2021, 11, 529. https://doi.org/10.3390/life11060529
Boulajfene W, Lasram M, Zouari-Tlig S. Integrated Biomarker Response for Environmental Assessment Using the Gastropod Phorcus turbinatus along the Northern and the Northeastern Coasts of Tunisia. Life. 2021; 11(6):529. https://doi.org/10.3390/life11060529
Chicago/Turabian StyleBoulajfene, Wafa, Montassar Lasram, and Sabiha Zouari-Tlig. 2021. "Integrated Biomarker Response for Environmental Assessment Using the Gastropod Phorcus turbinatus along the Northern and the Northeastern Coasts of Tunisia" Life 11, no. 6: 529. https://doi.org/10.3390/life11060529