
Insulin-Like Growth Factor-II and Ischemic Stroke—A Prospective Observational Study

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Vascular Risk Factors, Stroke Severity, and Subtypes
2.3. Functional Outcome and Mortality
2.4. Blood Sampling and Biochemical Analysis
2.5. Statistical Evaluation
2.6. Ethics Approval and Consent to Participate
3. Results
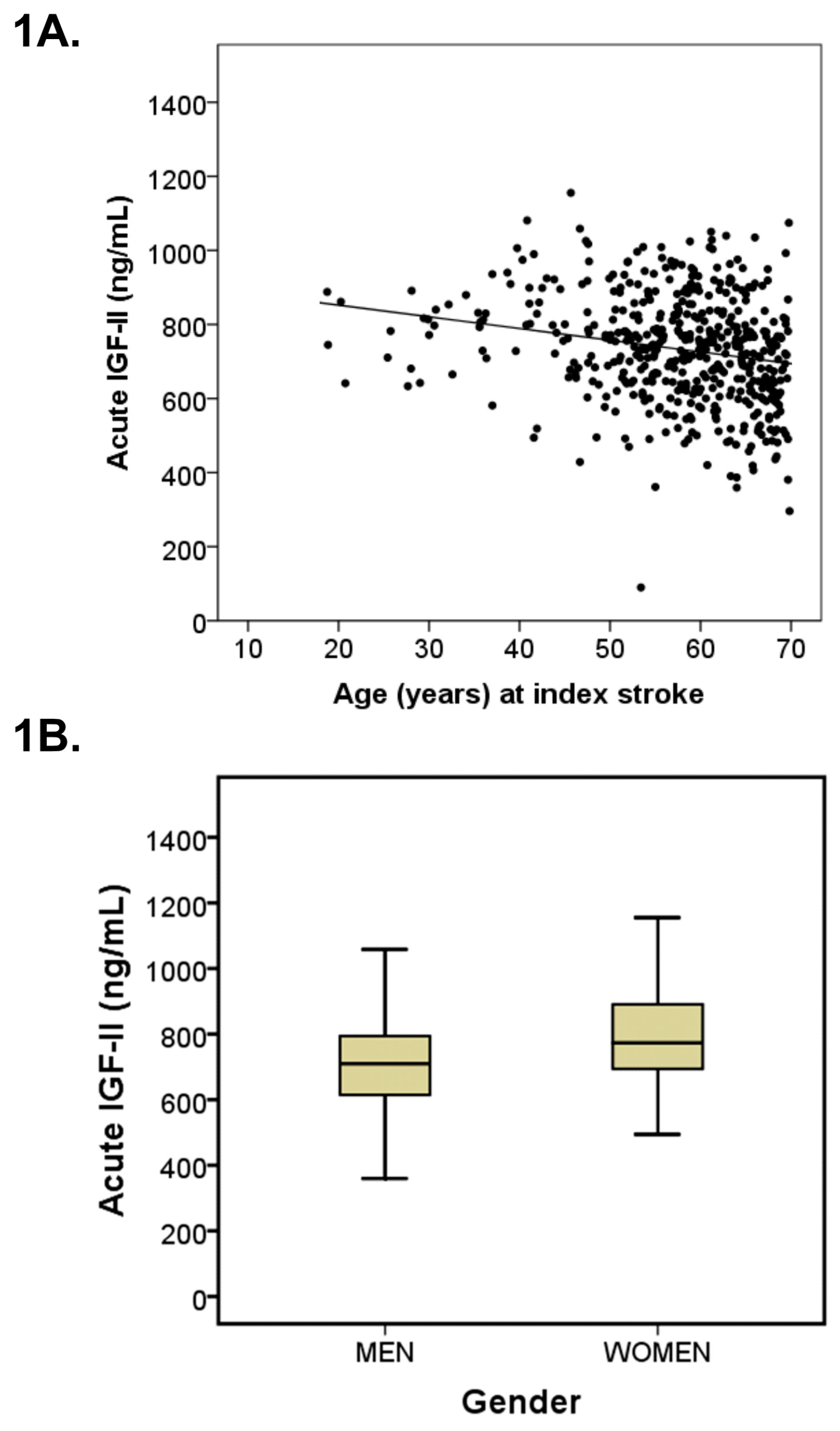
3.1. Baseline Characteristics
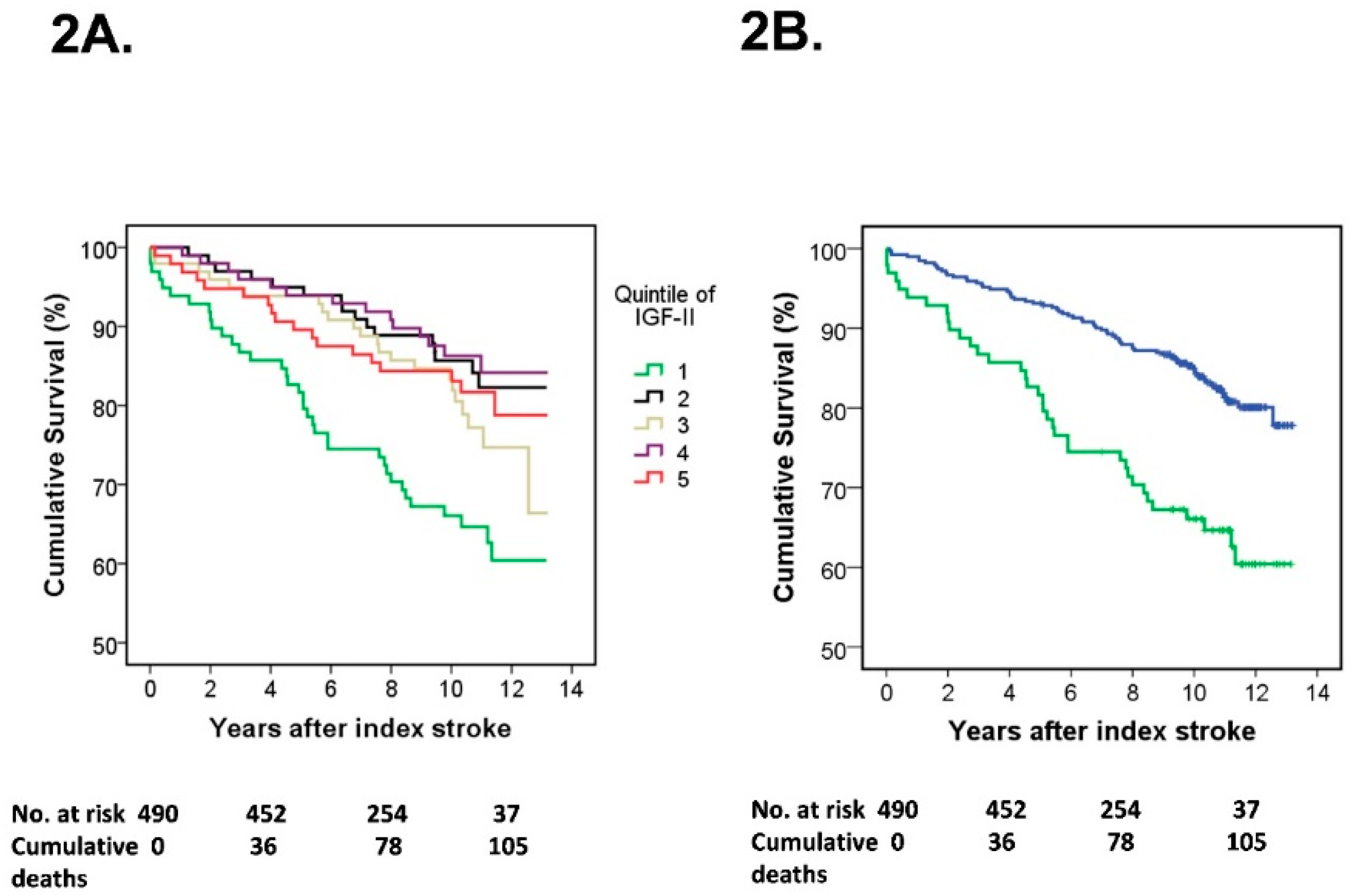
3.2. S-IGF-II and Risk of Mortality
3.3. Serum IGF-II and Functional Outcome
4. Discussion
4.1. IGF-II Levels in Normal Subjects and Stroke Patients
4.2. IGF-II and Mortality
4.3. Modulation of IGF-II Activity
4.4. IGF-II and Functional Outcome
4.5. Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Le Roith, D.; Butler, A.A. Insulin-like Growth Factors in Pediatric Health and Disease. J. Clin. Endocrinol. Metab. 1999, 84, 4355–4361. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, E.E. Insulin-like growth factors in the brain and their potential clinical implications. Neurology 2012, 79, 2148–2153. [Google Scholar] [CrossRef] [PubMed]
- Adamek, A.; Kasprzak, A. Insulin-Like Growth Factor (IGF) System in Liver Diseases. Int. J. Mol. Sci. 2018, 19, 1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohlsson, C.; Mohan, S.; Sjögren, K.; Tivesten, A.; Isgaard, J.; Isaksson, O.; Jansson, J.-O.; Svensson, J. The Role of Liver-Derived Insulin-Like Growth Factor-I. Endocr. Rev. 2009, 30, 494–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera, E.J.; Goldin, A.; Fulmer, N.; Tavares, R.; Wands, J.R.; De La Monte, S.M. Insulin and insulin-like growth factor expression and function deteriorate with progression of Alzheimer’s disease: Link to brain reductions in acetylcholine. J. Alzheimer’s Dis. 2005, 8, 247–268. [Google Scholar] [CrossRef]
- Reinhardt, R.R.; A Bondy, C. Insulin-like growth factors cross the blood-brain barrier. Endocrinology 1994, 135, 1753–1761. [Google Scholar] [CrossRef]
- Russo, V.C.; Gluckman, P.D.; Feldman, E.L.; Werther, G.A. The Insulin-Like Growth Factor System and Its Pleiotropic Functions in Brain. Endocr. Rev. 2005, 26, 916–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, A.M.; Torres-Alemán, I. The many faces of insulin-like peptide signalling in the brain. Nat. Rev. Neurosci. 2012, 13, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C.; Kar, S. The insulin-like growth factor-II/mannose-6-phosphate receptor: Structure, distribution and function in the central nervous system. Brain Res. Rev. 2004, 44, 117–140. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C.; Jhamandas, J.H.; Harris, K.H.; Fu, W.; Macdonald, R.G.; Kar, S. Single Transmembrane Domain Insulin-Like Growth Factor-II/Mannose-6-Phosphate Receptor Regulates Central Cholinergic Function by Activating a G-Protein-Sensitive, Protein Kinase C-Dependent Pathway. J. Neurosci. 2006, 26, 585–596. [Google Scholar] [CrossRef]
- Masunaga, Y.; Inoue, T.; Yamoto, K.; Fujisawa, Y.; Sato, Y.; Kawashima-Sonoyama, Y.; Morisada, N.; Iijima, K.; Ohata, Y.; Namba, N.; et al. IGF2 Mutations. J. Clin. Endocrinol. Metab. 2019, 105, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Hertze, J.; Nägga, K.; Minthon, L.; Hansson, O. Changes in cerebrospinal fluid and blood plasma levels of IGF-II and its binding proteins in Alzheimer’s disease: An observational study. BMC Neurol. 2014, 14, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.Y.; Stern, S.A.; Garcia-Osta, A.; Saunier-Rebori, B.; Pollonini, G.; Bambah-Mukku, D.; Blitzer, R.D.; Alberini, C.M. A critical role for IGF-II in memory consolidation and enhancement. Nat. Cell Biol. 2011, 469, 491–497. [Google Scholar] [CrossRef]
- Shahmoradi, A.; Radyushkin, K.; Rossner, M.J. Enhanced memory consolidation in mice lacking the circadian modulators Sharp1 and -2 caused by elevated Igf2 signaling in the cortex. Proc. Natl. Acad. Sci. USA 2015, 112, E3582–E3589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinmetz, A.B.; Johnson, S.A.; Iannitelli, D.E.; Pollonini, G.; Alberini, C.M. Insulin-like growth factor 2 rescues aging-related memory loss in rats. Neurobiol. Aging 2016, 44, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Castilla-Cortázar, I.; García-Fernández, M.; Delgado, G.; E Puche, J.; Sierra, I.; Barhoum, R.; González-Barón, S. Hepatoprotection and neuroprotection induced by low doses of IGF-II in aging rats. J. Transl. Med. 2011, 9, 103. [Google Scholar] [CrossRef] [Green Version]
- Montañez, E.M.; Pavia, J.; Santin, L.J.; Boraldi, F.; Estivill-Torrus, G.; Aguirre, J.A.; Garcia-Fernandez, M. Involvement of IGF-II receptors in the antioxidant and neuroprotective effects of IGF-II on adult cortical neuronal cultures. Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 1041–1051. [Google Scholar] [CrossRef] [Green Version]
- Montañez, E.M.; Millon, C.; Boraldi, F.; Garcia-Guirado, F.; Pedraza, C.; Lara, E.; Santin, L.; Pavia, J.; Garcia-Fernandez, M. IGF-II promotes neuroprotection and neuroplasticity recovery in a long-lasting model of oxidative damage induced by glucocorticoids. Redox Biol. 2017, 13, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-H.; Singh, P.; Tsark, W.M.K.; Szabó, P.E. Complete Biallelic Insulation at the H19/Igf2 Imprinting Control Region Position Results in Fetal Growth Retardation and Perinatal Lethality. PLoS ONE 2010, 5, e12630. [Google Scholar] [CrossRef] [Green Version]
- O’Dell, S.; Miller, G.; Cooper, J.; Hindmarsh, P.; Pringle, P.; Ford, H.; Humphries, S.; Day, I. Apal polymorphism in insulin-like growth factor II (IGF2) gene and weight in middle-aged males. Int. J. Obes. 1997, 21, 822–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Dell, S.D.; Day, I.N. Molecules in focus Insulin-like growth factor II (IGF-II). Int. J. Biochem. Cell Biol. 1998, 30, 767–771. [Google Scholar] [CrossRef]
- Stessman, J.; Maaravi, Y.; Hammerman-Rozenberg, R.; Cohen, A.; Nemanov, L.; Gritsenko, I.; Gruberman, N.; Ebstein, R.P. Candidate genes associated with ageing and life expectancy in the Jerusalem longitudinal study. Mech. Ageing Dev. 2005, 126, 333–339. [Google Scholar] [CrossRef] [PubMed]
- De Luca, M.; Rose, G.; Bonafè, M.; Garasto, S.; Greco, V.; Weir, B.; Franceschi, C.; De Benedictis, G. Sex-specific longevity associations defined by Tyrosine Hydroxylase–Insulin–Insulin Growth Factor 2 haplotypes on the 11p15.5 chromosomal region. Exp. Gerontol. 2001, 36, 1663–1671. [Google Scholar] [CrossRef]
- Beilharz, E.J.; Bassett, N.S.; Sirimanne, E.S.; Williams, C.E.; Gluckman, P.D. Insulin-like growth factor II is induced during wound repair following hypoxic-ischemic injury in the developing rat brain. Mol. Brain Res. 1995, 29, 81–91. [Google Scholar] [CrossRef]
- Lee, W.-H.; Clemens, J.A.; Bondy, C.A. Insulin-like growth factors in the response to cerebral ischemia. Mol. Cell. Neurosci. 1992, 3, 36–43. [Google Scholar] [CrossRef]
- Stephenson, D.T.; Rash, K.; Clemens, J.A. Increase in Insulin-like Growth Factor II Receptor within Ischemic Neurons following Focal Cerebral Infarction. Br. J. Pharmacol. 1995, 15, 1022–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Yan, J.; Bregere, C.; Zelmer, A.; Goerne, T.; Kapfhammer, J.P.; Guzman, R.; Wellmann, S. RBM3 promotes neurogenesis in a niche-dependent manner via IMP2-IGF2 signaling pathway after hypoxic-ischemic brain injury. Nat. Commun. 2019, 10, 3983. [Google Scholar] [CrossRef] [Green Version]
- Johnsen, S.P.; Hundborg, H.H.; Sørensen, H.T.; Ørskov, H.; Tjonneland, A.; Overvad, K.; Jørgensen, J.O.L. Insulin-Like Growth Factor (IGF) I, -II, and IGF Binding Protein-3 and Risk of Ischemic Stroke. J. Clin. Endocrinol. Metab. 2005, 90, 5937–5941. [Google Scholar] [CrossRef]
- Åberg, D.; Johansson, P.; Isgaard, J.; Wallin, A.; Johansson, J.-O.; Andreasson, U.; Blennow, K.; Zetterberg, H.; Åberg, N.D.; Svensson, J. Increased Cerebrospinal Fluid Level of Insulin-like Growth Factor-II in Male Patients with Alzheimer’s Disease. J. Alzheimer’s Dis. 2015, 48, 637–646. [Google Scholar] [CrossRef]
- Tham, A.; Nordberg, A.; Grissom, F.E.; Carlsson-Skwirut, C.; Viitanen, M.; Sara, V.R. Insulin-like growth factors and insulin-like growth factor binding proteins in cerebrospinal fluid and serum of patients with dementia of the Alzheimer type. J. Neural Transm. 1993, 5, 165–176. [Google Scholar] [CrossRef]
- Jood, K.; Ladenvall, C.; Rosengren, A.; Blomstrand, C.; Jern, C. Family History in Ischemic Stroke before 70 Years of Age. Stroke 2005, 36, 1383–1387. [Google Scholar] [CrossRef] [Green Version]
- Åberg, D.; Jood, K.; Blomstrand, C.; Jern, C.; Nilsson, M.; Isgaard, J.; Åberg, N.D. Serum IGF-I Levels Correlate to Improvement of Functional Outcome after Ischemic Stroke. J. Clin. Endocrinol. Metab. 2011, 96, E1055–E1064. [Google Scholar] [CrossRef] [Green Version]
- Åberg, D.; Åberg, N.D.; Jood, K.; Holmegaard, L.; Redfors, P.; Blomstrand, C.; Isgaard, J.; Jern, C.; Svensson, J. Homeostasis model assessment of insulin resistance and outcome of ischemic stroke in non-diabetic patients—A prospective observational study. BMC Neurol. 2019, 19, 177. [Google Scholar] [CrossRef]
- Åberg, N.D.; Åberg, D.; Jood, K.; Nilsson, M.; Blomstrand, C.; Kuhn, H.G.; Svensson, J.; Jern, C.; Isgaard, J. Altered levels of circulating insulin-like growth factor I (IGF-I) following ischemic stroke are associated with outcome—A prospective observational study. BMC Neurol. 2018, 18, 106. [Google Scholar] [CrossRef] [PubMed]
- Åberg, N.D.; Åberg, D.; Lagging, C.; Holmegaard, L.; Redfors, P.; Jood, K.; Nilsson, M.; Aberg, M.; Blomstrand, C.; Svensson, J.; et al. Association Between Levels of Serum Insulin-like Growth Factor I and Functional Recovery, Mortality, and Recurrent Stroke at a 7-year Follow-up. Exp. Clin. Endocrinol. Diabetes 2019, 128, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Watts, E.L.; Perez-Cornago, A.; Appleby, P.N.; Albanes, D.; Ardanaz, E.; Black, A.; Bueno-De-Mesquita, H.B.; Chan, J.M.; Chen, C.; Chubb, S.P.; et al. The associations of anthropometric, behavioural and sociodemographic factors with circulating concentrations of IGF-I, IGF-II, IGFBP-1, IGFBP-2 and IGFBP-3 in a pooled analysis of 16,024 men from 22 studies. Int. J. Cancer 2019, 145, 3244–3256. [Google Scholar] [CrossRef] [PubMed]
- Ceda, G.P.; Dall’Aglio, E.; Magnacavallo, A.; Vargas, N.; Fontana, V.; Maggio, M.; Valenti, G.; Lee, P.D.K.; Hintz, R.L.; Hoffman, A.R. The Insulin-Like Growth Factor Axis and Plasma Lipid Levels in the Elderly1. J. Clin. Endocrinol. Metab. 1998, 83, 499–502. [Google Scholar] [CrossRef] [Green Version]
- Zaina, S.; Pettersson, L.; Thomsen, A.B.; Chai, C.-M.; Qi, Z.; Thyberg, J.; Nilsson, J. Shortened Life Span, Bradycardia, and Hypotension in Mice with Targeted Expression of anIgf2Transgene in Smooth Muscle Cells. Endocrinology 2003, 144, 2695–2703. [Google Scholar] [CrossRef] [Green Version]
- Husted, C.I.; Valencik, M. Insulin-like growth factors and their potential role in cardiac epigenetics. J. Cell. Mol. Med. 2016, 20, 1589–1602. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, S.; Gaunt, T.R.; O’Dell, S.D.; Chen, X.-H.; Gu, N.; Hawe, E.; Miller, G.J.; Humphries, S.E.; Day, I.N. Haplotypic analyses of the IGF2-INS-TH gene cluster in relation to cardiovascular risk traits. Hum. Mol. Genet. 2004, 13, 715–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiansen, J.; Kolte, A.M.; O Hansen, T.V.; Nielsen, F.C. IGF2 mRNA-binding protein 2: Biological function and putative role in type 2 diabetes. J. Mol. Endocrinol. 2009, 43, 187–195. [Google Scholar] [CrossRef]
- Ng, M.C.Y.; Shriner, D.; Chen, B.H.; Li, M.; Chen, W.-M.; Guo, X.; Liu, J.; Bielinski, S.J.; Yanek, L.R.; Nalls, M.A.; et al. Meta-Analysis of Genome-Wide Association Studies in African Americans Provides Insights into the Genetic Architecture of Type 2 Diabetes. PLoS Genet. 2014, 10, e1004517. [Google Scholar] [CrossRef]
- Mercader, J.M.; Liao, R.G.; Bell, A.D.; Dymek, Z.; Estrada, K.; Tukiainen, T.; Huerta-Chagoya, A.; Moreno-Macías, H.; Jablonski, K.A.; Hanson, R.L.; et al. A Loss-of-Function Splice Acceptor Variant in IGF2 Is Protective for Type 2 Diabetes. Diabetes 2017, 66, 2903–2914. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, M.S.; Gibson, J.M.; Heald, A.H.; Dunger, D.B.; Wareham, N.J. Low Circulating IGF-II Concentrations Predict Weight Gain and Obesity in Humans. Diabetes 2003, 52, 1403–1408. [Google Scholar] [CrossRef] [Green Version]
- Inzaghi, E.; Ferroli, B.B.; Fintini, D.; Grossi, A.; Nobili, V.; Cianfarani, S. Insulin-Like Growth Factors and Metabolic Syndrome in Obese Children. Horm. Res. Paediatr. 2017, 87, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Pouriamehr, S.; Barmaki, H.; Rastegary, M.; Lotfi, F.; Afjadi, M.N. Investigation of insulin-like growth factors/insulin-like growth factor binding proteins regulation in metabolic syndrome patients. BMC Res. Notes 2019, 12, 653. [Google Scholar] [CrossRef] [Green Version]
- Hjortebjerg, R.; Tarnow, L.; Jorsal, A.; Parving, H.-H.; Rossing, P.; Bjerre, M.; Frystyk, J. IGFBP-4 Fragments as Markers of Cardiovascular Mortality in Type 1 Diabetes Patients with and without Nephropathy. J. Clin. Endocrinol. Metab. 2015, 100, 3032–3040. [Google Scholar] [CrossRef] [Green Version]
- Holly, J.M.P.; Biernacka, K.; Perks, C.M. The Neglected Insulin: IGF-II, a Metabolic Regulator with Implications for Diabetes, Obesity, and Cancer. Cells 2019, 8, 1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallagher, E.J.; Leroith, D. Minireview: IGF, Insulin, and Cancer. Endocrinology 2011, 152, 2546–2551. [Google Scholar] [CrossRef] [Green Version]
- Murphy, S.K.; Huang, Z.; Wen, Y.; Spillman, M.A.; Whitaker, R.S.; Simel, L.R.; Nichols, T.D.; Marks, J.R.; Berchuck, A. Frequent IGF2/H19 Domain Epigenetic Alterations and Elevated IGF2 Expression in Epithelial Ovarian Cancer. Mol. Cancer Res. 2006, 4, 283–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, R.; DeCoteau, J.F.; Geyer, C.; Gao, M.; Cui, H.; Casson, A.G. Loss of imprinting of the insulin-like growth factor II (IGF2) gene in esophageal normal and adenocarcinoma tissues. Carcinogenesis 2009, 30, 2117–2122. [Google Scholar] [CrossRef] [PubMed]
- Liou, J.-M.; Shun, C.-T.; Liang, J.-T.; Chiu, H.-M.; Chen, M.-J.; Chen, C.; Wang, H.-P.; Wu, M.-S.; Lin, J.-T. Plasma Insulin-Like Growth Factor-Binding Protein-2 Levels as Diagnostic and Prognostic Biomarker of Colorectal Cancer. J. Clin. Endocrinol. Metab. 2010, 95, 1717–1725. [Google Scholar] [CrossRef] [Green Version]
- Dai, N. The Diverse Functions of IMP2/IGF2BP2 in Metabolism. Trends Endocrinol. Metab. 2020, 31, 670–679. [Google Scholar] [CrossRef]
- Coderch, C.; de Cerio, M.D.; Zapico, J.M.; Peláez, R.; Larrayoz, I.M.; Ramos, A.; Martínez, A.; de Pascual-Teresa, B. In silico identification and in vivo characterization of small molecule therapeutic hypothermia mimetics. Bioorg. Med. Chem. 2017, 25, 6597–6604. [Google Scholar] [CrossRef]
- Peduzzi, P.; Concato, J.; Kemper, E.; Holford, T.R.; Feinstein, A.R. A simulation study of the number of events per variable in logistic regression analysis. J. Clin. Epidemiol. 1996, 49, 1373–1379. [Google Scholar] [CrossRef]
- Heald, A.H.; Anderson, S.G.; Ivison, F.; Laing, I.; Gibson, J.M.; Cruickshank, K. C-reactive protein and the insulin-like growth factor (IGF)-system in relation to risk of cardiovascular disease in different ethnic groups. Atherosclerosis 2003, 170, 79–86. [Google Scholar] [CrossRef]
- Ali, K.; Cheek, E.; Sills, S.; Crome, P.; Roffe, C. Development of a Conversion Factor to Facilitate Comparison of National Institute of Health Stroke Scale Scores with Scandinavian Stroke Scale Scores. Cerebrovasc. Dis. 2007, 24, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Gray, L.J.; Ali, M.; Lyden, P.D.; Bath, P.M. Interconversion of the National Institutes of Health Stroke Scale and Scandinavian Stroke Scale in Acute Stroke. J. Stroke Cerebrovasc. Dis. 2009, 18, 466–468. [Google Scholar] [CrossRef] [Green Version]
- Adams, H.P.; Bendixen, B.H.; Kappelle, L.J.; Biller, J.; Love, B.B.; Gordon, D.L.; E Marsh, E. Classification of subtype of acute ischemic stroke. Definitions for use in a multicenter clinical trial. TOAST. Trial of Org 10172 in Acute Stroke Treatment. Stroke 1993, 24, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladenvall, C.; Jood, K.; Blomstrand, C.; Nilsson, S.; Jern, C.; Ladenvall, P. Serum C-Reactive Protein Concentration and Genotype in Relation to Ischemic Stroke Subtype. Stroke 2006, 37, 2018–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondanelli, M.; Ambrosio, M.R.; Zatelli, M.C.; Basaglia, N.; Degli Uberti, E.C. Prevalence of hypopituitarism in patients with cerebrovascular diseases. J. Endocrinol. Investig. 2008, 31, 16–20. [Google Scholar]



{kind=link}
{kind=link}
| Controls (n = 514) | Patients (n = 492) | Large Vessel Disease (n = 54) | Small Vessel Disease (n = 106) | Cardioembolic Stroke (n = 81) Cryptogenic Stroke (n = 131) | ||
|---|---|---|---|---|---|---|
| Age (years) | 57.2 ± 0.44 | 57.2 ± 0.44 | 58.5 ± 1.00 | 59.3 ± 0.66 * | 58.9 ± 1.14 | 54.1 ± 0.94 ** |
| Male sex, n (%) | 329 (64) | 315 (64) | 39 (72) | 66 (62) | 66 (68) | 77 (59) |
| Hypertension, n (%) | 199 (39) | 299 (62) *** | 30 (60) *** | 78 (74) *** | 44 (56) ** | 73 (57) *** |
| Diabetes mellitus, n (%) | 30 (6) | 94 (19) *** | 19 (38) *** | 25 (24) *** | 15 (19) *** | 17 (13) *** |
| Current smoker, n (%) | 89 (17) | 193 (39) *** | 27 (54) *** | 46 (43) *** | 29 (37) *** | 50 (39) *** |
| hsCRP (mg/L) | 3.06 ± 0.26 | 10.8 ± 1.00 *** | 12.1 ± 2.51 *** | 4.95 ± 0.66 ** | 16.7 ±3.18 *** | 7.98 ± 1.52 ** |
| Imputed LDL (ng/nL) | 3.33 ± 0.04 | 3.35 ± 0.04 | 3.54 ± 0.15 | 3.48 ± 0.08 | 3.08 ± 0.11 * | 3.32 ± 0.07 |
| HOMA-IR | 2.13 ± 0.12 | 4.80 ± 0.25 *** | 6.14 ± 0.88 *** | 5.06 ± 0.71 *** | 4.59 ± 0.53 *** | 3.78 ± 0.27 *** |
| BMI (kg/m2) | 26.5 ± 0.18 | 26.7 ± 0.20 | 26.8 ± 0.65 | 27.0 ± 0.42 | 26.8 ± 0.51 | 26.4 ± 0.34 |
| NIHSS score baseline | NA | 5.32 ± 0.25 | 6.49 ± 0.85 | 3.32 ± 0.26 | 6.48 ± 0.78 | 5.24 ± 0.48 |
| mRS score 3 months | NA | 1.79 ± 0.05 | 2.14 ± 0.16 | 1.35 ± 0.10 | 2.01 ± 0.15 | 1.73 ± 0.10 |
| mRS score 2 years | NA | 1.88 ± 0.06 | 1.77 ± 0.24 | 1.49 ± 0.11 | 2.17 ± 0.17 | 1.68 ± 0.10 |
| s-IGF-II acute (ng/mL) | 712.1 ± 5.58 | 734.7 ± 7.08 * | 747.0 ± 18.8 | 734.6 ± 13.7 | 667.9 ± 17.1 ** | 764.4 ± 12.2 *** |
| s-IGF-II 3 months (ng/mL) | NA | 736.8 ± 6.91 ** | 751.9 ± 23.3 | 739.5 ± 14.8 | 679.4 ± 14.5 * | 748.4 ± 11.0 ** |
| Total Stroke Population | Acute Serum IGF-II Quintile 1 | Quintiles 2–5 | p-Value |
|---|---|---|---|
| Deaths, n (%) | 36 (36.7%) | 70 (17.9%) | |
| Crude | 2.34 (1.56–3.49) | 1.0 referent | <0.001 |
| Multivariate model A | 1.96 (1.29–2.95) | 1.0 referent | 0.001 |
| Multivariate model B | 1.93 (1.23–3.03) | 1.0 referent | 0.004 |
| Multivariate model C | 1.65 (1.04–2.63) | 1.0 referent | 0.035 |
| Multivariate model D | 1.64 (1.02–2.61) | 1.0 referent | 0.039 |
| Total Stroke Population | Quintile 1 | Quintile 2–5 | p-Value | n |
|---|---|---|---|---|
| 3-month mRS | ||||
| Crude | 2.30 (1.39–3.82) | 1.0 referent | 0.001 | 463 |
| Multivariate model A | 2.24 (1.33–3.79) | 1.0 referent | 0.002 | 463 |
| Multivariate model B | 1.95 (1.10–3.45) | 1.0 referent | 0.023 | 446 |
| Multivariate model C | 0.99 (0.46–2.13) | 1.0 referent | 0.98 | 446 |
| Multivariate model D | 0.71 (0.30–1.64) | 1.0 referent | 0.42 | 445 |
| 2-year mRS | ||||
| Crude | 1.93 (1.17–3.18) | 1.0 referent | 0.010 | 485 |
| Multivariate model A | 1.76 (1.04–2.95) | 1.0 referent | 0.034 | 485 |
| Multivariate model B | 1.59 (0.90–2.80) | 1.0 referent | 0.11 | 463 |
| Multivariate model C | 0.90 (0.46–1.77) | 1.0 referent | 0.76 | 463 |
| Multivariate model D | 0.79 (0.39–1.58) | 1.0 referent | 0.50 | 462 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Åberg, D.; Åberg, N.D.; Jood, K.; Redfors, P.; Blomstrand, C.; Isgaard, J.; Jern, C.; Svensson, J. Insulin-Like Growth Factor-II and Ischemic Stroke—A Prospective Observational Study. Life 2021, 11, 499. https://doi.org/10.3390/life11060499
Åberg D, Åberg ND, Jood K, Redfors P, Blomstrand C, Isgaard J, Jern C, Svensson J. Insulin-Like Growth Factor-II and Ischemic Stroke—A Prospective Observational Study. Life. 2021; 11(6):499. https://doi.org/10.3390/life11060499
Chicago/Turabian StyleÅberg, Daniel, N. David Åberg, Katarina Jood, Petra Redfors, Christian Blomstrand, Jörgen Isgaard, Christina Jern, and Johan Svensson. 2021. "Insulin-Like Growth Factor-II and Ischemic Stroke—A Prospective Observational Study" Life 11, no. 6: 499. https://doi.org/10.3390/life11060499