
Tenderness of the Skin after Chemical Stimulation of Underlying Temporal and Thoracolumbar Fasciae Reveals Somatosensory Crosstalk between Superficial and Deep Tissues

 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Participants
2.2. Experimental Protocols
2.2.1. Electrical Stimulation
2.2.2. Chemical Stimulation
2.3. Quantitative Sensory Testing (QST)
2.3.1. Thermal Testing and Thermal Test Devices
2.3.2. Mechanical Testing
2.4. Statistics
3. Results
3.1. Somatosensory Profile of the Skin after High Frequency Electrical Stimulation (HFS) of the Thoracolumbar Fascia (TLF) or Underlying Multifidus Muscle (MM)
3.2. Mechanical Pain Thresholds (MPT) after Chemical Stimulation of Three Different Volumes of Hypertonic Saline into the Thoracolumbar Fascia (TLF) or Underlying Multifidus Muscle (MM)
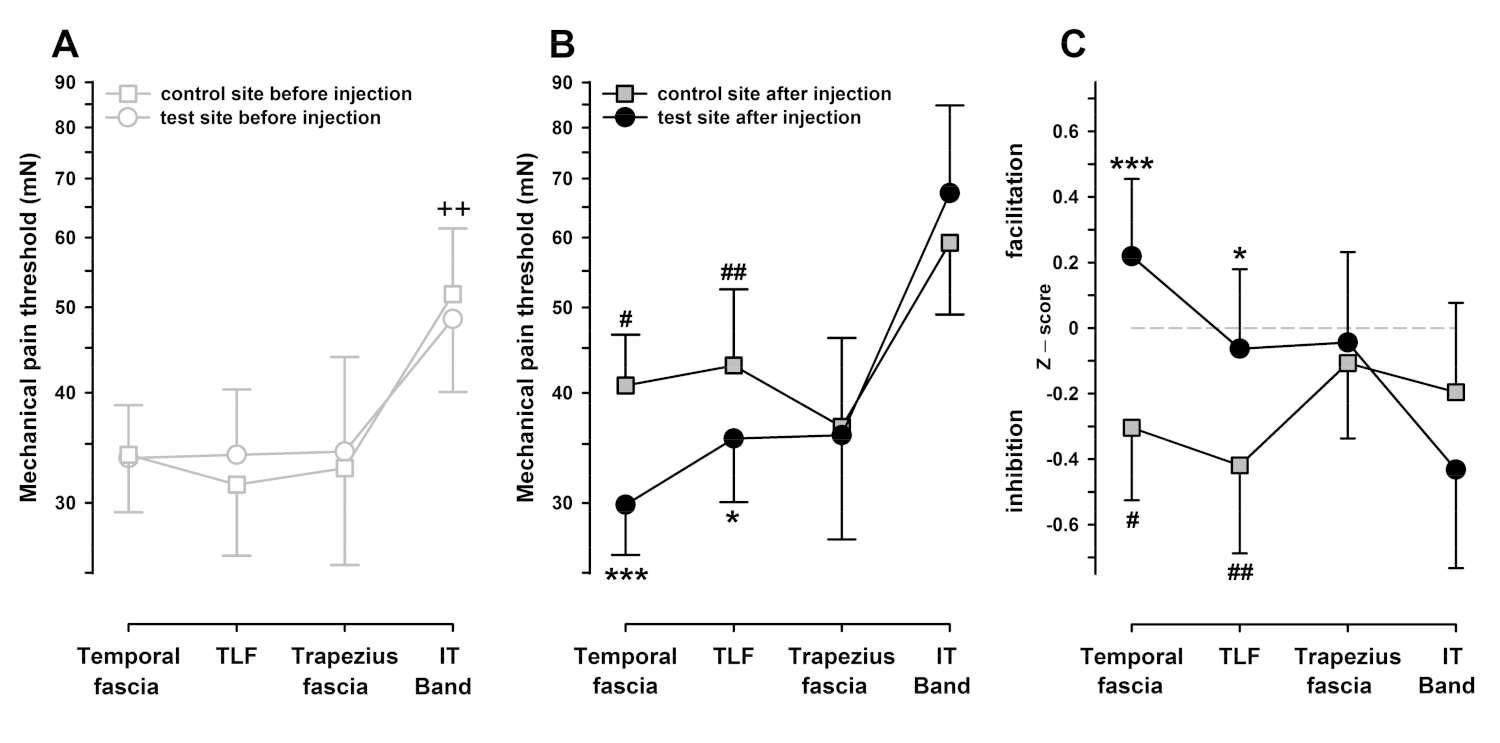
3.3. Mechanical Pain Thresholds (MPT) after Chemical Stimulation (Hypertonic Saline Injection) of the Temporal Fascia, Iliotibial Band, Trapezius Muscle Fascia, and Thoracolumbar Fascia
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergman, S. Management of musculoskeletal pain. Best Pract. Res. Clin. Rheumatol. 2007, 21, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Rice, A.S.C.; Smith, B.H.; Blyth, F.M. Pain and the global burden of disease. Pain 2016, 157, 791–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geneen, L.J.; Moore, R.A.; Clarke, C.; Martin, D.; Colvin, L.A.; Smith, B.H. Physical activity and exercise for chronic pain in adults: An overview of Cochrane Reviews. Cochrane Database Syst. Rev. 2017, 4, CD011279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayden, J.A.; Wilson, M.N.; Stewart, S.; Cartwright, J.L.; Smith, A.O.; Riley, R.D.; van Tulder, M.; Bendix, T.; Cecchi, F.; Costa, L.O.P.; et al. Exercise treatment effect modifiers in persistent low back pain: An individual participant data meta-analysis of 3514 participants from 27 randomised controlled trials. Br. J. Sports Med. 2020, 54, 1277–1278. [Google Scholar] [CrossRef]
- Polaski, A.M.; Phelps, A.L.; Kostek, M.C.; Szucs, K.A.; Kolber, B.J. Exercise-induced hypoalgesia: A meta-analysis of exercise dosing for the treatment of chronic pain. PLoS ONE 2019, 14, e0210418. [Google Scholar] [CrossRef] [Green Version]
- Kemp, H.I.; Corner, E.; Colvin, L.A. Chronic pain after COVID-19: Implications for rehabilitation. Br. J. Anaesth. 2020, 125, 436–440. [Google Scholar] [CrossRef]
- Rodriguez-Nogueira, O.; Leiros-Rodriguez, R.; Benitez-Andrades, J.A.; Alvarez-Alvarez, M.J.; Marques-Sanchez, P.; Pinto-Carral, A. Musculoskeletal pain and teleworking in times of the COVID-19: Analysis of the impact on the workers at two Spanish universities. Int. J. Environ. Res. Public Health 2020, 18, 31. [Google Scholar] [CrossRef]
- Rygh, L.J.; Svendsen, F.; Fiska, A.; Haugan, F.; Hole, K.; Tjølsen, A. Long-term potentiation in spinal nociceptive systems—how acute pain may become chronic. Psychoneuroendocrinology 2005, 30, 959–964. [Google Scholar] [CrossRef]
- Sandkühler, J. Learning and memory in pain pathways. Pain 2000, 88, 113–118. [Google Scholar] [CrossRef]
- Henrich, F.; Magerl, W.; Klein, T.; Greffrath, W.; Treede, R.D. Capsaicin-sensitive C- and A-fibre nociceptors control long-term potentiation-like pain amplification in humans. Brain J. Neurol. 2015, 138, 2505–2520. [Google Scholar] [CrossRef] [Green Version]
- Schilder, A.; Magerl, W.; Hoheisel, U.; Klein, T.; Treede, R.D. Electrical high-frequency stimulation of the human thoracolumbar fascia evokes long-term potentiation-like pain amplification. Pain 2016, 157, 2309–2317. [Google Scholar] [CrossRef]
- Woolf, C.J. Central sensitization: Implications for the diagnosis and treatment of pain. Pain 2011, 152, S2–S15. [Google Scholar] [CrossRef]
- Banic, B.; Petersen-Felix, S.; Andersen, O.K.; Radanov, B.P.; Villiger, P.M.; Arendt-Nielsen, L.; Curatolo, M. Evidence for spinal cord hypersensitivity in chronic pain after whiplash injury and in fibromyalgia. Pain 2004, 107, 7–15. [Google Scholar] [CrossRef]
- Price, D.D.; Staud, R.; Robinson, M.E.; Mauderli, A.P.; Cannon, R.; Vierck, C.J. Enhanced temporal summation of second pain and its central modulation in fibromyalgia patients. Pain 2002, 99, 49–59. [Google Scholar] [CrossRef]
- Corey, S.M.; Vizzard, M.A.; Badger, G.J.; Langevin, H.M. Sensory innervation of the nonspecialized connective tissues in the low back of the rat. Cells Tissues Organ. 2011, 194, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Tesarz, J.; Hoheisel, U.; Wiedenhofer, B.; Mense, S. Sensory innervation of the thoracolumbar fascia in rats and humans. Neuroscience 2011, 194, 302–308. [Google Scholar] [CrossRef]
- Fede, C.; Porzionato, A.; Petrelli, L.; Fan, C.; Pirri, C.; Biz, C.; De Caro, R.; Stecco, C. Fascia and soft tissues innervation in the human hip and their possible role in post-surgical pain. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2020, 38, 1646–1654. [Google Scholar] [CrossRef]
- Casato, G.; Stecco, C.; Busin, R. Role of fasciae in nonspecific low back pain. Eur. J. Transl. Myol. 2019, 29, 8330. [Google Scholar] [CrossRef]
- Rolke, R.; Magerl, W.; Campbell, K.A.; Schalber, C.; Caspari, S.; Birklein, F.; Treede, R.D. Quantitative sensory testing: A comprehensive protocol for clinical trials. Eur. J. Pain 2006, 10, 77–88. [Google Scholar] [CrossRef]
- Klein, T.; Magerl, W.; Hopf, H.C.; Sandkuhler, J.; Treede, R.D. Perceptual correlates of nociceptive long-term potentiation and long-term depression in humans. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 964–971. [Google Scholar] [CrossRef] [Green Version]
- Klein, T.; Magerl, W.; Treede, R.D. Perceptual correlate of nociceptive long-term potentiation (LTP) in humans shares the time course of early-LTP. J. Neurophysiol. 2006, 96, 3551–3555. [Google Scholar] [CrossRef] [Green Version]
- Lang, S.; Klein, T.; Magerl, W.; Treede, R.D. Modality-specific sensory changes in humans after the induction of long-term potentiation (LTP) in cutaneous nociceptive pathways. Pain 2007, 128, 254–263. [Google Scholar] [CrossRef]
- Klein, T.; Stahn, S.; Magerl, W.; Treede, R.D. The role of heterosynaptic facilitation in long-term potentiation (LTP) of human pain sensation. Pain 2008, 139, 507–519. [Google Scholar] [CrossRef]
- Pfau, D.B.; Klein, T.; Putzer, D.; Pogatzki-Zahn, E.M.; Treede, R.D.; Magerl, W. Analysis of hyperalgesia time courses in humans after painful electrical high-frequency stimulation identifies a possible transition from early to late LTP-like pain plasticity. Pain 2011, 152, 1532–1539. [Google Scholar] [CrossRef]
- Van den Broeke, E.N.; van Heck, C.H.; van Rijn, C.M.; Wilder-Smith, O.H. Neural correlates of heterotopic facilitation induced after high frequency electrical stimulation of nociceptive pathways. Molec. Pain 2011, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Van den Broeke, E.N.; Mouraux, A. High-frequency electrical stimulation of the human skin induces heterotopical mechanical hyperalgesia, heat hyperalgesia, and enhanced responses to nonnociceptive vibrotactile input. J. Neurophysiol. 2014, 111, 1564–1573. [Google Scholar] [CrossRef] [Green Version]
- Matre, D.; Olsen, M.B.; Jacobsen, L.M.; Klein, T.; Gjerstad, J. Induction of the perceptual correlate of human long-term potentiation (LTP) is associated with the 5-HTT genotype. Brain Res. 2013, 1491, 54–59. [Google Scholar] [CrossRef]
- Schilder, A.; Magerl, W.; Klein, T.; Treede, R.D. Assessment of pain quality reveals distinct differences between nociceptive innervation of low back fascia and muscle in humans. Pain Rep. 2018, 3, e662. [Google Scholar] [CrossRef]
- Ji, R.R.; Kohno, T.; Moore, K.A.; Woolf, C.J. Central sensitization and LTP: Do pain and memory share similar mechanisms? Trends Neurosci. 2003, 26, 696–705. [Google Scholar] [CrossRef]
- Cooke, S.F.; Bliss, T.V.P. Plasticity in the human central nervous system. Brain J. Neurol. 2006, 129, 1659–1673. [Google Scholar] [CrossRef] [Green Version]
- Ruscheweyh, R.; Wilder-Smith, O.; Drdla, R.; Liu, X.G.; Sandkuhler, J. Long-term potentiation in spinal nociceptive pathways as a novel target for pain therapy. Molec. Pain 2011, 7, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, T.J.; Inyang, K.E. Commonalities between pain and memory mechanisms and their meaning for understanding chronic pain. Prog. Mol. Biol. Transl. Sci. 2015, 131, 409–434. [Google Scholar] [CrossRef] [Green Version]
- Wall, P.D.; Woolf, C.J. Muscle but not cutaneous C-afferent input produces prolonged increases in the excitability of the flexion reflex in the rat. J. Physiol. 1984, 356, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Lambertz, D.; Hoheisel, U.; Mense, S. Distribution of synaptic field potentials induced by TTX-resistant skin and muscle afferents in rat spinal segments L4 and L5. Neurosci. Lett. 2006, 409, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Kellgren, J.H. Referred Pains from Muscle. Br. Med. J. 1938, 1, 325–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graven-Nielsen, T.; McArdle, A.; Phoenix, J.; Arendt-Nielsen, L.; Jensen, T.S.; Jackson, M.J.; Edwards, R.H. In vivo model of muscle pain: Quantification of intramuscular chemical, electrical, and pressure changes associated with saline-induced muscle pain in humans. Pain 1997, 69, 137–143. [Google Scholar] [CrossRef]
- Schilder, A.; Hoheisel, U.; Magerl, W.; Benrath, J.; Klein, T.; Treede, R.D. Sensory findings after stimulation of the thoracolumbar fascia with hypertonic saline suggest its contribution to low back pain. Pain 2014, 155, 222–231. [Google Scholar] [CrossRef]
- Svensson, P.; Graven-Nielsen, T.; Arendt-Nielsen, L. Mechanical hyperesthesia of human facial skin induced by tonic painful stimulation of jaw muscles. Pain 1998, 74, 93–100. [Google Scholar] [CrossRef]
- Magerl, W.; Klein, T. Experimental human models of neuropathic pain. In Handbook of Clinical Neurology; Aminoff, M.J., Boller, F., Schwab, D.F., Cervero, F., Jensen, T.S., Eds.; Elsevier: Edinburgh, UK, 2006; Volume 81, pp. 503–516. [Google Scholar]
- Pirri, C.; Fede, C.; Petrelli, L.; Guidolin, D.; Fan, C.; De Caro, R.; Stecco, C. An anatomical comparison of the fasciae of the thigh: A macroscopic, microscopic and ultrasound imaging study. J. Anat. 2021, 238, 999–1009. [Google Scholar] [CrossRef]
- Davidge, K.M.; van Furth, W.R.; Agur, A.; Cusimano, M. Naming the soft tissue layers of the temporoparietal region: Unifying anatomic terminology across surgical disciplines. Neurosurgery 2010, 62, 753–766. [Google Scholar] [CrossRef]
- Rolke, R.; Baron, R.; Maier, C.; Tolle, T.R.; Treede, R.D.; Beyer, A.; Binder, A.; Birbaumer, N.; Birklein, F.; Botefur, I.C.; et al. Quantitative sensory testing in the German research network on neuropathic pain (DFNS): Standardized protocol and reference values. Pain 2006, 123, 231–243. [Google Scholar] [CrossRef]
- Magerl, W.; Krumova, E.K.; Baron, R.; Tolle, T.; Treede, R.D.; Maier, C. Reference data for quantitative sensory testing (QST): Refined stratification for age and a novel method for statistical comparison of group data. Pain 2010, 151, 598–605. [Google Scholar] [CrossRef]
- Ziegler, E.A.; Magerl, W.; Meyer, R.A.; Treede, R.D. Secondary hyperalgesia to punctate mechanical stimuli: Central sensitization to A-fibre nociceptor input. Brain J. Neurol. 1999, 122, 2245–2257. [Google Scholar] [CrossRef] [Green Version]
- Magerl, W.; Wilk, S.H.; Treede, R.D. Secondary hyperalgesia and perceptual wind-up following intradermal injection of capsaicin in humans. Pain 1998, 74, 257–268. [Google Scholar] [CrossRef]
- Capra, N.F.; Ro, J.Y. Human and animal experimental models of acute and chronic muscle pain: Intramuscular algesic injection. Pain 2004, 110, 3–7. [Google Scholar] [CrossRef]
- Reinert, A.; Treede, R.D.; Bromm, B. The pain inhibiting pain effect: An electrophysiological study in humans. Brain Res. 2000, 862, 103–110. [Google Scholar] [CrossRef]
- Yu, X.M.; Mense, S. Response properties and descending control of rat dorsal horn neurons with deep receptive fields. Neuroscience 1990, 39, 823–831. [Google Scholar]
- Cervero, F.; Schaible, H.G.; Schmidt, R.F. Tonic descending inhibition of spinal cord neurones driven by joint afferents in normal cats and in cats with an inflamed knee joint. Exp. Brain Res. 1991, 83, 675–678. [Google Scholar] [CrossRef]
- Vanegas, H.; Schaible, H.G. Descending control of persistent pain: Inhibitory or facilitatory? Brain Res. Rev. 2004, 46, 295–309. [Google Scholar] [CrossRef]
- Vollert, J.; Magerl, W.; Baron, R.; Binder, A.; Enax-Krumova, E.K.; Geisslinger, G.; Gierthmuhlen, J.; Henrich, F.; Hullemann, P.; Klein, T.; et al. Pathophysiological mechanisms of neuropathic pain: Comparison of sensory phenotypes in patients and human surrogate pain models. Pain 2018, 159, 1090–1102. [Google Scholar] [CrossRef]
- LaMotte, R.H.; Shain, C.N.; Simone, D.A.; Tsai, E.F.P. Neurogenic hyperalgesia: Psychophysical studies of underlying mechanisms. J. Neurophysiol. 1991, 66, 190–211. [Google Scholar] [CrossRef] [PubMed]
- Simone, D.A.; Sorkin, L.S.; Oh, U.; Chung, J.M.; Owens, C.; LaMotte, R.H.; Willis, W.D. Neurogenic hyperalgesia: Central neural correlates in responses of spinothalamic tract neurons. J. Neurophysiol. 1991, 66, 228–246. [Google Scholar] [CrossRef] [PubMed]
- Treede, R.D.; Meyer, R.A.; Raja, S.N.; Campbell, J.N. Peripheral and central mechanisms of cutaneous hyperalgesia. Prog. Neurobiol. 1992, 38, 397–421. [Google Scholar] [CrossRef]
- Head, H. On the disturbances of sensation with especial reference to the pain of visceral disease. Brain J. Neurol. 1893, 16, 1–133. [Google Scholar] [CrossRef]
- Woolf, C.J. Evidence for a central component of post-injury pain hypersensitivity. Nature 1983, 306, 686–688. [Google Scholar] [CrossRef]
- Zhang, J.; Hoheisel, U.; Klein, T.; Magerl, W.; Mense, S.; Treede, R.D. High-frequency modulation of rat spinal field potentials: Effects of slowly conducting muscle vs. skin afferents. J. Neurophysiol. 2016, 115, 692–700. [Google Scholar] [CrossRef] [Green Version]
- Gibson, W.; Arendt-Nielsen, L.; Taguchi, T.; Mizumura, K.; Graven-Nielsen, T. Increased pain from muscle fascia following eccentric exercise: Animal and human findings. Exp. Brain Res. 2009, 194, 299–308. [Google Scholar] [CrossRef]
- Graven-Nielsen, T.; Babenko, V.; Svensson, P.; Arendt-Nielsen, L. Experimentally induced muscle pain induces hypoalgesia in heterotopic deep tissues, but not in homotopic deep tissues. Brain Res. 1998, 787, 203–210. [Google Scholar] [CrossRef]
- Lo Vecchio, S.; Petersen, L.J.; Finocchietti, S.; Gazerani, P.; Arendt-Nielsen, L.; Graven-Nielsen, T. The effect of combined skin and deep tissue inflammatory pain models. Pain Med. 2015, 16, 2053–2064. [Google Scholar] [CrossRef] [Green Version]
- Lo Vecchio, S.; Petersen, L.J.; Finocchietti, S.; Gazerani, P.; Christensen, S.W.; Arendt-Nielsen, L.; Graven-Nielsen, T. Interaction between ultraviolet B-induced cutaneous hyperalgesia and nerve growth factor-induced muscle hyperalgesia. Eur. J. Pain 2016, 20, 1058–1069. [Google Scholar] [CrossRef] [Green Version]
- Hoheisel, U.; Taguchi, T.; Treede, R.D.; Mense, S. Nociceptive input from the rat thoracolumbar fascia to lumbar dorsal horn neurones. Eur. J. Pain 2011, 15, 810–815. [Google Scholar] [CrossRef]
- Barry, C.M.; Kestell, G.; Gillan, M.; Haberberger, R.V.; Gibbins, I.L. Sensory nerve fibers containing calcitonin gene-related peptide in gastrocnemius, latissimus dorsi and erector spinae muscles and thoracolumbar fascia in mice. Neuroscience 2015, 291, 106–117. [Google Scholar] [CrossRef]
- Graven-Nielsen, T.; Mense, S. The peripheral apparatus of muscle pain: Evidence from animal and human studies. Clin. J. Pain 2001, 17, 2–10. [Google Scholar] [CrossRef]
- Mense, S. Innervation of the thoracolumbar fascia. Eur. J. Transl. Myol. 2019, 29, 8297. [Google Scholar] [CrossRef]
- Mantyh, P.W.; Rogers, S.D.; Honoré, P.; Allen, B.J.; Ghilardi, J.R.; Li, J.; Daughters, R.S.; Lappi, D.A.; Wiley, R.G.; Simone, D.A. Inhibition of hyperalgesia by ablation of lamina I spinal neurons expressing the substance P receptor. Science 1997, 278, 275–279. [Google Scholar] [CrossRef]
- Khasabov, S.G.; Rogers, S.D.; Ghilardi, J.R.; Peters, C.M.; Mantyh, P.W.; Simone, D.A. Spinal neurons that possess the substance P receptor are required for the development of central sensitization. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 9086–9098. [Google Scholar] [CrossRef]
- Stecco, A.; Gesi, M.; Stecco, C.; Stern, R. Fascial components of the myofascial pain syndrome. Curr. Pain Headache Rep. 2013, 17, 352. [Google Scholar] [CrossRef]
- Moriwaki, K.; Yuge, O. Topographical features of cutaneous tactile hypoesthetic and hyperesthetic abnormalities in chronic pain. Pain 1999, 81, 1–6. [Google Scholar] [CrossRef]
- Magerl, W.; Treede, R.D. Secondary tactile hypoesthesia: A novel type of pain-induced somatosensory plasticity in human subjects. Neurosci. Lett. 2004, 361, 136–139. [Google Scholar] [CrossRef]
- Stohler, C.S.; Kowalski, C.J.; Lund, J.P. Muscle pain inhibits cutaneous touch perception. Pain 2001, 92, 327–333. [Google Scholar] [CrossRef]
- Moriwaki, K.; Shiroyama, K.; Yasuda, M.; Uesugi, F. Reversible tactile hypoesthesia associated with myofascial trigger points: A pilot study on prevalence and clinical implications. Pain Rep. 2019, 4, e772. [Google Scholar] [CrossRef]
- Ramaswamy, S.; Wodehouse, T. Conditioned pain modulation-A comprehensive review. Neurophysiol. Clin. 2021. [Google Scholar] [CrossRef]
- Stammler, T.; De Col, R.; Seifert, F.; Maihöfner, C. Functional imaging of sensory decline and gain induced by differential noxious stimulation. NeuroImage 2008, 42, 1151–1163. [Google Scholar] [CrossRef]
- Sandkühler, J. The organization and function of endogenous antinociceptive systems. Prog. Neurobiol. 1996, 50, 49–81. [Google Scholar] [CrossRef]
- Prescott, S.A. Interactions between depression and facilitation within neural networks: Updating the dual-process theory of plasticity. Learn. Mem. 1998, 5, 446–466. [Google Scholar]
- Benjamin, M. The fascia of the limbs and back--A review. J. Anat. 2009, 214, 1–18. [Google Scholar] [CrossRef]
- Willard, F.H.; Vleeming, A.; Schuenke, M.D.; Danneels, L.; Schleip, R. The thoracolumbar fascia: Anatomy, function and clinical considerations. J. Anat. 2012, 221, 507–536. [Google Scholar] [CrossRef] [Green Version]
- Wilke, J.; Schleip, R.; Yucesoy, C.A.; Banzer, W. Not merely a protective packing organ? A review of fascia and its force transmission capacity. J. Appl. Physiol. 2018, 124, 234–244. [Google Scholar] [CrossRef]
- Stecco, C.; Gagey, O.; Belloni, A.; Pozzuoli, A.; Porzionato, A.; Macchi, V.; Aldegheri, R.; De Caro, R.; Delmas, V. Anatomy of the deep fascia of the upper limb. Second part: Study of innervation. Morphologie 2007, 91, 38–43. [Google Scholar] [CrossRef]
- Renton, T. Chronic orofacial pain. Oral Dis. 2017, 23, 566–571. [Google Scholar] [CrossRef]
- De Tommaso, M.; Fernandez-de-Las-Penas, C. Tension Type Headache. Curr. Rheumatol. Rev. 2016, 12, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Patrick, N.; Emanski, E.; Knaub, M.A. Acute and chronic low back pain. Med. Clin. N. Am. 2016, 100, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Wilke, J.; Schleip, R.; Klingler, W.; Stecco, C. The Lumbodorsal Fascia as a Potential Source of Low Back Pain: A Narrative Review. BioMed Res. Int. 2017, 2017, 5349620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langevin, H.M.; Sherman, K.J. Pathophysiological model for chronic low back pain integrating connective tissue and nervous system mechanisms. Med. Hypoth. 2007, 68, 74–80. [Google Scholar] [CrossRef]
- Deising, S.; Weinkauf, B.; Blunk, J.; Obreja, O.; Schmelz, M.; Rukwied, R. NGF-evoked sensitization of muscle fascia nociceptors in humans. Pain 2012, 153, 1673–1679. [Google Scholar] [CrossRef]
- Weinkauf, B.; Deising, S.; Obreja, O.; Hoheisel, U.; Mense, S.; Schmelz, M.; Rukwied, R. Comparison of nerve growth factor-induced sensitization pattern in lumbar and tibial muscle and fascia. Muscle Nerv. 2015, 52, 265–272. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | dfeffect, dferror | F-Value | p-Value | Level of Significance |
|---|---|---|---|---|
| (1) Before vs. after injection | 115 | 4.25 | 0.057 | (*) |
| (2) Muscle vs. fascia | 115 | 2,53 | 0.133 | |
| (3) Test site vs. control site | 115 | 2.01 | 0.176 | |
| (4) Injected volumes | 330 | 0.77 | 0.473 | |
| 1 × 2 interaction | 115 | 2.53 | 0.133 | |
| 1 × 3 interaction | 115 | 2.02 | 0.176 | |
| 1 × 4 interaction | 115 | 0.47 | 0.631 | |
| 2 × 3 interaction | 115 | 0.09 | 0.767 | |
| 2 × 4 interaction | 330 | 2.03 | 0.149 | |
| 3 × 4 interaction | 330 | 0.71 | 0.498 | |
| 1 × 2 × 3 interaction | 115 | 0.60 | 0.810 | |
| 1 × 2 × 4 interaction | 330 | 0.87 | 0.428 | |
| 1 × 3 × 4 interaction | 330 | 0.84 | 0.441 | |
| 2 × 3 × 4 interaction | 330 | 0.98 | 0.387 | |
| 1 × 2 × 3 × 4 interaction | 330 | 0.03 | 0.968 |
| Effect | dfeffect, dferror | F-Value | p-Value | Level of Significance |
|---|---|---|---|---|
| (1) Before vs. after injection | 115 | 4.14 | 0.060 | (*) |
| (2) Test site vs. control site | 115 | 0.64 | 0.436 | |
| (3) Test areas | 345 | 4.48 | 0.008 | ** |
| 1 × 2 interaction | 115 | 1.77 | 0.204 | |
| 1 × 3 interaction | 345 | 1.01 | 0.525 | |
| 2 × 3 interaction | 345 | 2.24 | 0.397 | |
| 1 × 2 × 3 interaction | 345 | 1.58 | 0.097 | (*) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magerl, W.; Thalacker, E.; Vogel, S.; Schleip, R.; Klein, T.; Treede, R.-D.; Schilder, A. Tenderness of the Skin after Chemical Stimulation of Underlying Temporal and Thoracolumbar Fasciae Reveals Somatosensory Crosstalk between Superficial and Deep Tissues. Life 2021, 11, 370. https://doi.org/10.3390/life11050370
Magerl W, Thalacker E, Vogel S, Schleip R, Klein T, Treede R-D, Schilder A. Tenderness of the Skin after Chemical Stimulation of Underlying Temporal and Thoracolumbar Fasciae Reveals Somatosensory Crosstalk between Superficial and Deep Tissues. Life. 2021; 11(5):370. https://doi.org/10.3390/life11050370
Chicago/Turabian StyleMagerl, Walter, Emanuela Thalacker, Simon Vogel, Robert Schleip, Thomas Klein, Rolf-Detlef Treede, and Andreas Schilder. 2021. "Tenderness of the Skin after Chemical Stimulation of Underlying Temporal and Thoracolumbar Fasciae Reveals Somatosensory Crosstalk between Superficial and Deep Tissues" Life 11, no. 5: 370. https://doi.org/10.3390/life11050370