
Characterisation of Early Microbial Colonisers within the Spiral Colon of Pre- and Post-Natal Piglets

 ,
,  ,
,
Abstract
:1. Background
2. Materials and Methods
2.1. Animals and Experimental Procedures
2.2. Extraction of DNA and 16S rRNA Amplicon Sequencing
2.3. Statistical Analysis
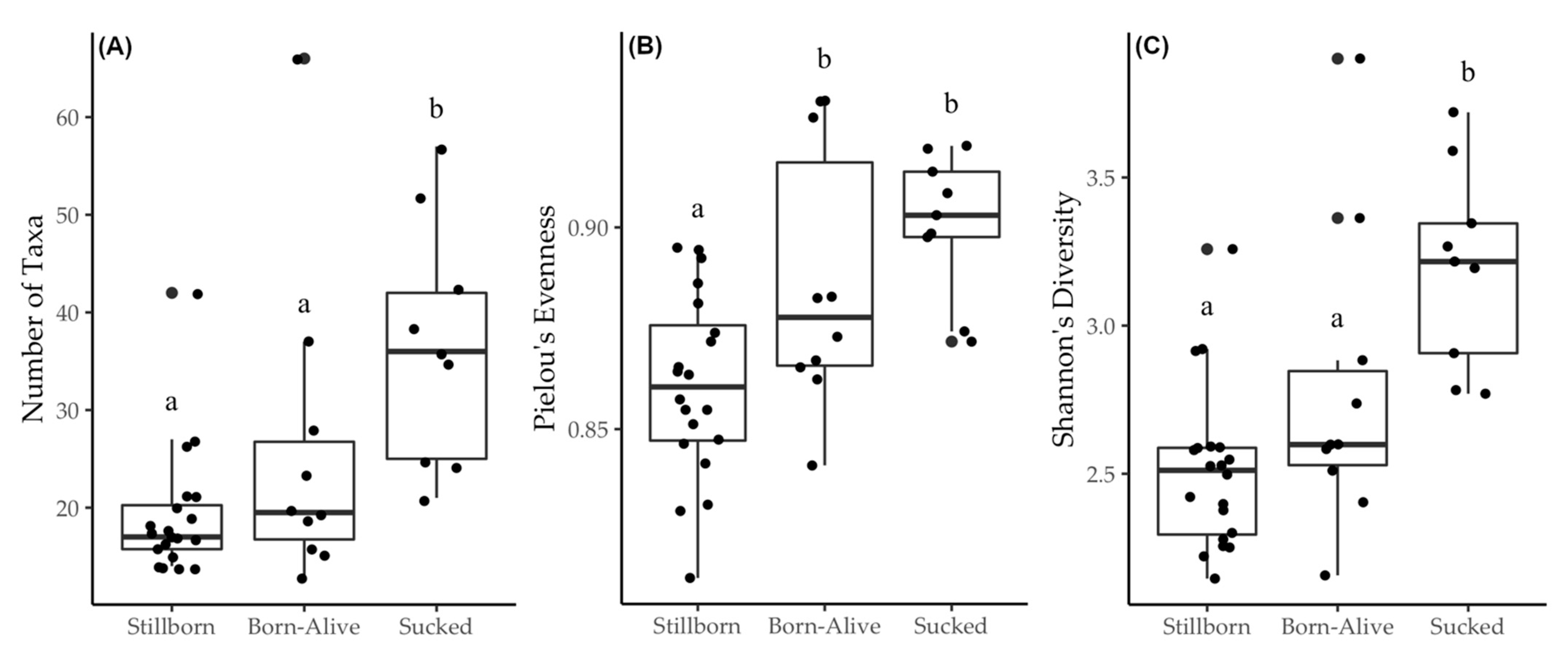
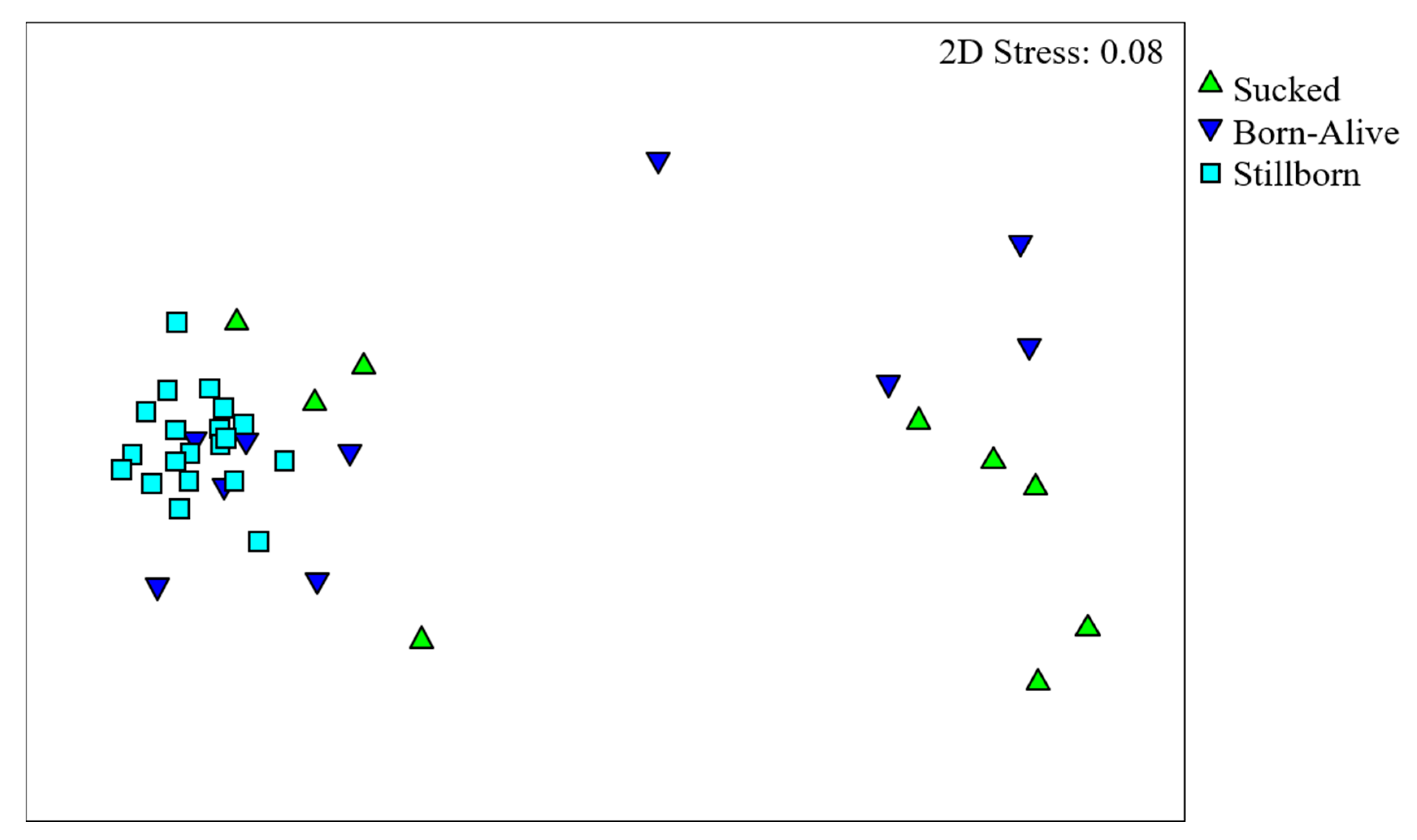
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nowland, T.L.; Plush, K.J.; Barton, M.; Kirkwood, R.N. Development and function of the intestinal microbiome and potential implications for pig production. Animals 2019, 9, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rautava, S.; Luoto, R.; Salminen, S.; Isolauri, E. Microbial contact during pregnancy, intestinal colonization and human disease. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Rautava, S.; Aakko, J.; Isolauri, E.; Salminen, S. Human gut colonisation may be initiated in utero by distinct microbial communities in the placenta and amniotic fluid. Sci. Rep. 2016, 6, 23129. [Google Scholar] [CrossRef] [Green Version]
- Younge, N.; McCann, J.R.; Ballard, J.; Plunkett, C.; Akhtar, S.; Araujo-Perez, F.; Murtha, A.; Brandon, D.; Seed, P.C. Fetal exposure to the maternal microbiota in humans and mice. JCI Insight 2019, 4, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, D.; Prince, A.; Ma, J.; Baquero, K.; Blundell, P.; Takahashi, D.; Dean, T.; Grove, K.; Aagaard, K. 114: Evidence of fetal microbiota and its maternal origins in a non-human primate model. Am. J. Obstet. Gynecol. 2017, 216, S80. [Google Scholar] [CrossRef]
- Perez-Munoz, M.E.; Arrieta, M.C.; Ramer-Tait, A.E.; Walter, J. A critical assessment of the “sterile womb” and “in utero colonization” hypotheses: Implications for research on the pioneer infant microbiome. Microbiome 2017, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- De Goffau, M.C.; Lager, S.; Sovio, U.; Gaccioli, F.; Cook, E.; Peacock, S.J.; Parkhill, J.; Charnock-Jones, D.S.; Smith, G.C.S. Human placenta has no microbiome but can contain potential pathogens. Nature 2019, 572, 329–334. [Google Scholar] [CrossRef]
- Perez, P.F.; Dore, J.; Leclerc, M.; Levenez, F.; Benyacoub, J.; Serrant, P.; Segura-Roggero, I.; Schiffrin, E.J.; Donnet-Hughes, A. Bacterial imprinting of the neonatal immune system: Lessons from maternal cells? Pediatrics 2007, 119, E724–E732. [Google Scholar] [CrossRef]
- Gomez-Arango, L.F.; Barrett, H.L.; McIntyre, H.D.; Callaway, L.K.; Morrison, M.; Nitert, M.D. Contributions of the maternal oral and gut microbiome to placental microbial colonization in overweight and obese pregnant women. Sci. Rep. 2017, 7, 2860. [Google Scholar] [CrossRef]
- Ardissone, A.N.; de la Cruz, D.M.; Davis-Richardson, A.G.; Rechcigl, K.T.; Li, N.; Drew, J.C.; Murgas-Torrazza, R.; Sharma, R.; Hudak, M.L.; Triplett, E.W.; et al. Meconium microbiome analysis identifies bacteria correlated with premature birth. PLoS ONE 2014, 9, e90784. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, E.; Marin, M.L.; Martin, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernandez, L.; Rodriguez, J.M. Is meconium from healthy newborns actually sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef]
- Inman, C.F.; Haverson, K.; Konstantinov, S.R.; Jones, P.H.; Harris, C.; Smidt, H.; Miller, B.; Bailey, M.; Stokes, C. Rearing environment affects development of the immune system in neonates. Clin. Exp. Immunol. 2010, 160, 431–439. [Google Scholar] [CrossRef]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [Green Version]
- Martín, V.; Maldonado-Barragán, A.; Moles, L.; Rodriguez-Baños, M.; Campo, R.D.; Fernández, L.; Rodríguez, J.M.; Jiménez, E. Sharing of bacterial strains between breast milk and infant feces. J. Hum. Lact. 2012, 28, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xu, J.; Ren, E.; Su, Y.; Zhu, W. Co-occurrence of early gut colonization in neonatal piglets with microbiota in the maternal and surrounding delivery environments. Anaerobe 2018, 49, 30–40. [Google Scholar] [CrossRef]
- Baker, A.A.; Davis, E.; Spencer, J.D.; Moser, R.; Rehberger, T. The effect of a bacillus-based direct-fed microbial supplemented to sows on the gastrointestinal microbiota of their neonatal piglets. J. Anim. Sci. 2013, 91, 3390–3399. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Feng, C.; Tao, S.; Li, N.; Zuo, B.; Han, D.; Wang, J. Maternal imprinting of the neonatal microbiota colonization in intrauterine growth restricted piglets: A review. J. Anim. Sci. Biotechnol. 2019, 10, 88. [Google Scholar] [CrossRef]
- Langendijk, P.; Plush, K. Parturition and its relationship with stillbirths and asphyxiated piglets. Animals 2019, 9, 885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stirling, G.R.; Griffin, D.; Ophel-Keller, K.; McKay, A.; Hartley, A.; Curran, J.; Stirling, A.M.; Monsour, C.; Winch, J.; Hardie, B. Combining an initial risk assessment process with DNA assays to improve prediction of soilborne diseases caused by root-knot nematode (meloidogyne spp.) and fusarium oxysporum f. Sp. Lycopersici in the queensland tomato industry. Australas. Plant Path. 2004, 33, 285–293. [Google Scholar] [CrossRef]
- Torok, V.A.; Ophel-Keller, K.; Loo, M.; Hughes, R.J. Application of methods for identifying broiler chicken gut bacterial species linked with increased energy metabolism. Appl. Environ. Microbiol. 2008, 74, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Torok, V.A.; Percy, N.J.; Moate, P.J.; Ophel-Keller, K. Influence of dietary docosahexaenoic acid supplementation on the overall rumen microbiota of dairy cows and linkages with production parameters. Can. J. Microbiol. 2014, 60, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. Pear: A fast and accurate illumina paired-end read merger. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kucztnski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than blast. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. Uchime improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Uparse: Highly accurate otu sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16s rrna gene database and workbench compatible with arb. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Kruskal, J.B. Multidimensional scaling by optimizing a goodness of fit to a nonmetric hypothesis. Psychometrics 1964, 29, 1–28. [Google Scholar] [CrossRef]
- Shepard, R.N. The analysis of proximities: Multidimensional scaling with an unknown distance function. I. Psychometrika 1962, 27, 125–140. [Google Scholar] [CrossRef]
- Patil, Y.; Gooneratne, R.; Ju, X.-H. Interactions between host and gut microbiota in domestic pigs: A review. Gut Microbes 2020, 11, 310–334. [Google Scholar] [CrossRef]
- Petri, D.; Hill, J.E.; Van Kessel, A.G. Microbial succession in the gastrointestinal tract (git) of the preweaned pig. Livest. Sci. 2010, 133, 107–109. [Google Scholar] [CrossRef]
- Sanglard, L.P.; Schmitz-Esser, S.; Gray, K.A.; Linhares, D.C.L.; Yeoman, C.J.; Dekkers, J.C.M.; Niederwerder, M.C.; Serão, N.V.L. Investigating the relationship between vaginal microbiota and host genetics and their impact on immune response and farrowing traits in commercial gilts. J. Anim. Breed. Genet. 2020, 137, 84–102. [Google Scholar] [CrossRef]
- Wang, H.; Hu, C.; Cheng, C.; Cui, J.; Ji, Y.; Hao, X.; Li, Q.; Ren, W.; Deng, B.; Yin, Y.; et al. Unraveling the association of fecal microbiota and oxidative stress with stillbirth rate of sows. Theriogenology 2019, 136, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Spring, S.; Premathilake, H.; Bradway, C.; Shili, C.; DeSilva, U.; Carter, S.; Pezeshki, A. Effect of very low-protein diets supplemented with branched-chain amino acids on energy balance, plasma metabolomics and fecal microbiome of pigs. Sci. Rep. 2020, 10, 15859. [Google Scholar] [CrossRef] [PubMed]
- Konstantinov, S.R.; Awati, A.A.; Williams, B.A.; Miller, B.G.; Jones, P.; Stokes, C.R.; Akkermans, A.D.; Smidt, H.; de Vos, W.M. Post-natal development of the porcine microbiota composition and activities. Environ. Microbiol. 2006, 8, 1191–1199. [Google Scholar] [CrossRef] [PubMed]
- Inoue, R.; Tsukahara, T.; Nakanishi, N.; Ushida, K. Development of the intestinal microbiota in the piglet. J. Gen. Appl. Microbiol. 2005, 51, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guevarra, R.B.; Lee, J.H.; Lee, S.H.; Seok, M.J.; Kim, D.W.; Kang, B.N.; Johnson, T.J.; Isaacson, R.E.; Kim, H.B. Piglet gut microbial shifts early in life: Causes and effects. J. Anim. Sci. Biotechnol. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mao, S.Y.; Yang, C.F.; Zhu, W.Y. Phylogenetic analysis of methanogens in the pig feces. Curr. Microbiol. 2011, 62, 1386–1389. [Google Scholar] [CrossRef]
- Su, Y.; Bian, G.; Zhu, Z.; Smidt, H.; Zhu, W. Early methanogenic colonisation in the faeces of meishan and yorkshire piglets as determined by pyrosequencing analysis. Archaea 2014, 2014, 547908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieu-Lesme, F.; Delbes, C.; Sollelis, L. Recovery of partial 16s rdna sequences suggests the presence of crenarchaeota in the human digestive ecosystem. Curr. Microbiol. 2005, 51, 317–321. [Google Scholar] [CrossRef]
- Aviles-Rosa, E.O.; Rakhshandeh, A.; McGlone, J.J. Preliminary study: Depriving piglets of maternal feces for the first seven days post-partum changes piglet physiology and performance before and after weaning. Animals 2019, 9, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knecht, D.; Cholewińska, P.; Jankowska-Ma, K.A.; Czyź, K. Development of swine’s digestive tract microbiota and its relation to production indices—a review. Animals 2020, 10, 527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, J.E.; Brown, W.R. The immunoglobulins and immunoglobulin genes of swine. Vet. Immunol. Immunopathol. 1994, 43, 5–12. [Google Scholar] [CrossRef]
- Chen, W.; Mi, J.; Lv, N.; Gao, J.; Cheng, J.; Wu, R.; Ma, J.; Lan, T.; Liao, X. Lactation stage-dependency of the sow milk microbiota. Front. Microbiol. 2018, 9, 945. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
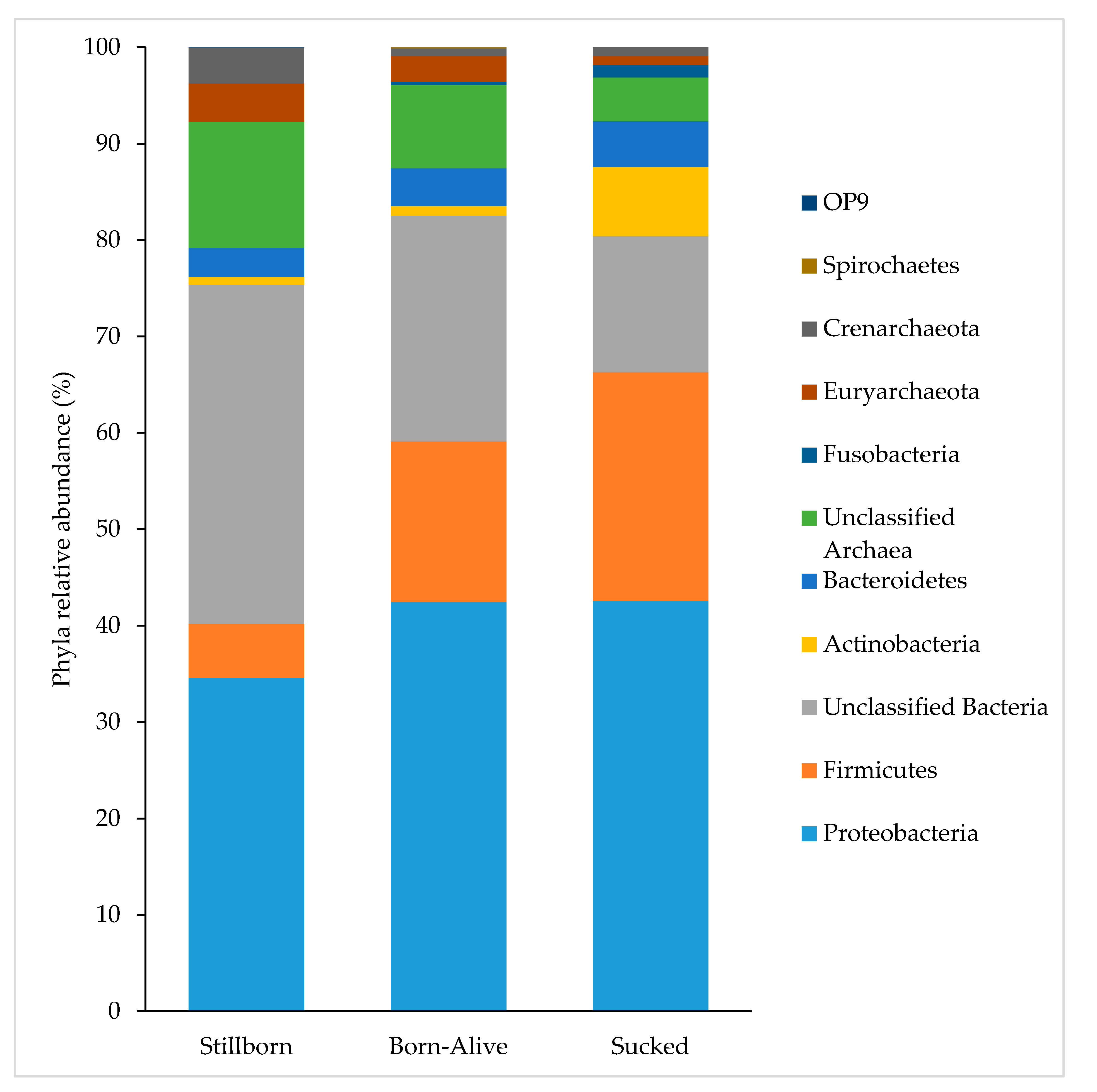{kind=link}
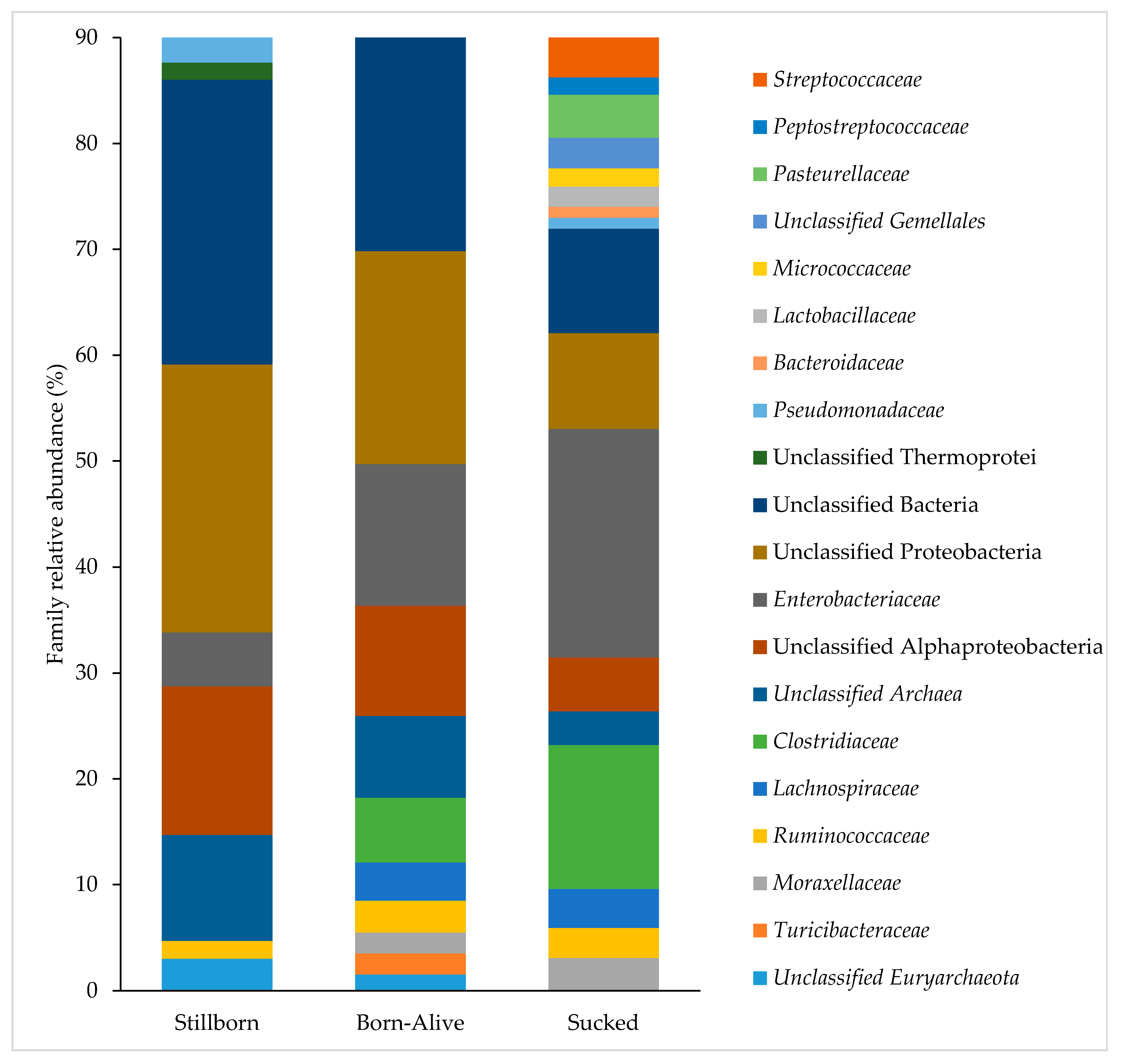{kind=link}
| Born-Alive | Stillborn | ||
|---|---|---|---|
| Genus | Average Abundance | Average Abundance | % |
| Unclassified Archaea | 0.67 | 1.03 | 2.15 |
| Unclassified Thermoprotei | 0.18 | 0.30 | 1.85 |
| Unclassified Crenarchaeota | 0.07 | 0.19 | 1.20 |
| Unclassified Euryarchaeota | 0.19 | 0.35 | 1.11 |
| Clostridium | 0.98 | 0.09 | 5.71 |
| Faecalibacterium | 0.10 | 0.11 | 0.79 |
| Unclassified Alphaproteobacteria | 1.00 | 1.44 | 2.91 |
| Unclassified Rickettsiales | 0.14 | 0.21 | 1.06 |
| Escherichia | 1.59 | 0.67 | 7.11 |
| Pseudomonas | 0.19 | 0.53 | 2.96 |
| Bacteroides | 0.14 | 0.15 | 0.82 |
| Sucked | Stillborn | ||
|---|---|---|---|
| Genus | Average Abundance | Average Abundance | % |
| Unclassified Archaea | 0.54 | 1.03 | 2.17 |
| Unclassified Crenarchaeota | 0.13 | 0.19 | 0.76 |
| Unclassified Thermoprotei | 0.10 | 0.30 | 1.16 |
| Unclassified Euryarchaeota | 0.16 | 0.35 | 0.92 |
| Unclassified Bacteria | 1.47 | 2.66 | 5.11 |
| Unclassified Gemellales | 0.45 | 0.03 | 1.85 |
| Lactobacillus | 0.39 | 0.04 | 1.53 |
| Streptococcus | 0.60 | 0.10 | 2.45 |
| Unclassified Clostridiaceae | 0.42 | 0.00 | 1.73 |
| Clostridium | 1.25 | 0.09 | 5.07 |
| SMB53 | 0.74 | 0.04 | 3.19 |
| Unclassified Peptostreptococcaceae | 0.31 | 0.00 | 1.29 |
| Unclassified Lachnospiraceae | 0.16 | 0.02 | 0.69 |
| Roseburia | 0.11 | 0.06 | 0.45 |
| Faecalibacterium | 0.16 | 0.11 | 0.58 |
| Unclassified Proteobacteria | 1.39 | 2.50 | 4.78 |
| Unclassified Alphaproteobacteria | 0.80 | 1.44 | 2.89 |
| Unclassified Rickettsiales | 0.07 | 0.21 | 0.82 |
| Escherichia | 2.28 | 0.67 | 7.20 |
| Actinobacillus | 0.83 | 0.00 | 3.37 |
| Pseudomonas | 0.30 | 0.53 | 2.01 |
| Unclassified S24-7 | 0.13 | 0.02 | 0.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowland, T.L.; Kirkwood, R.N.; Torok, V.A.; Plush, K.J.; Barton, M.D. Characterisation of Early Microbial Colonisers within the Spiral Colon of Pre- and Post-Natal Piglets. Life 2021, 11, 312. https://doi.org/10.3390/life11040312
Nowland TL, Kirkwood RN, Torok VA, Plush KJ, Barton MD. Characterisation of Early Microbial Colonisers within the Spiral Colon of Pre- and Post-Natal Piglets. Life. 2021; 11(4):312. https://doi.org/10.3390/life11040312
Chicago/Turabian StyleNowland, Tanya L., Roy N. Kirkwood, Valeria A. Torok, Kate J. Plush, and Mary D. Barton. 2021. "Characterisation of Early Microbial Colonisers within the Spiral Colon of Pre- and Post-Natal Piglets" Life 11, no. 4: 312. https://doi.org/10.3390/life11040312