
Toxicity and Anti-Proliferative Properties of Anisomeles indica Ethanol Extract on Cervical Cancer HeLa Cells and Zebrafish Embryos

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
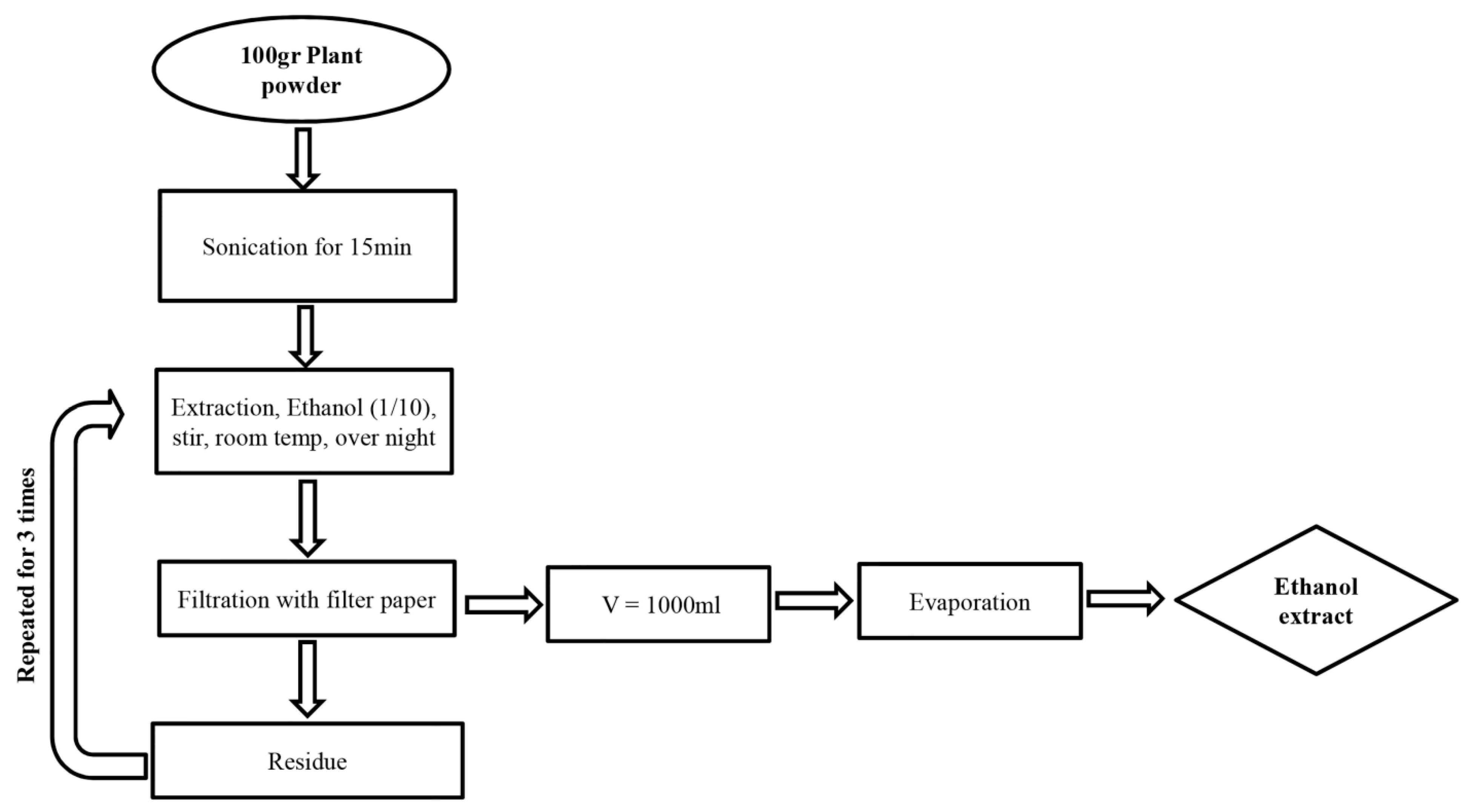
2.2. Preparation of Plant Extracts
2.3. Cell Culture
2.4. Embryo Zebrafish Toxicity Test
2.5. Cellular Toxicity MTT Assay
2.6. Annexin V Apoptosis Assay
2.7. Cell Cycle Analysis
2.8. Real-Time PCR Analysis
2.9. Colony Formation Assay
2.10. Statistical Analysis
3. Results
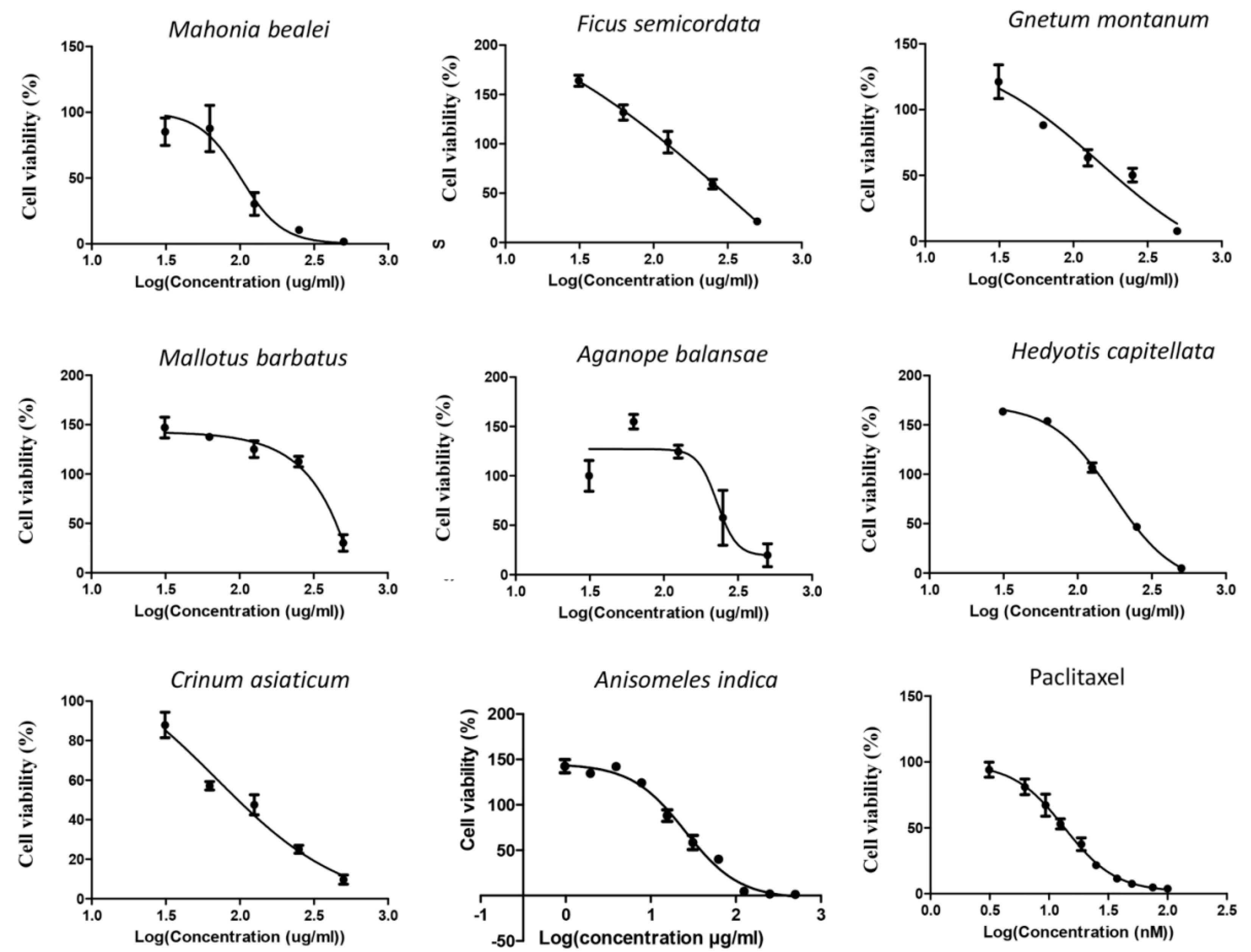
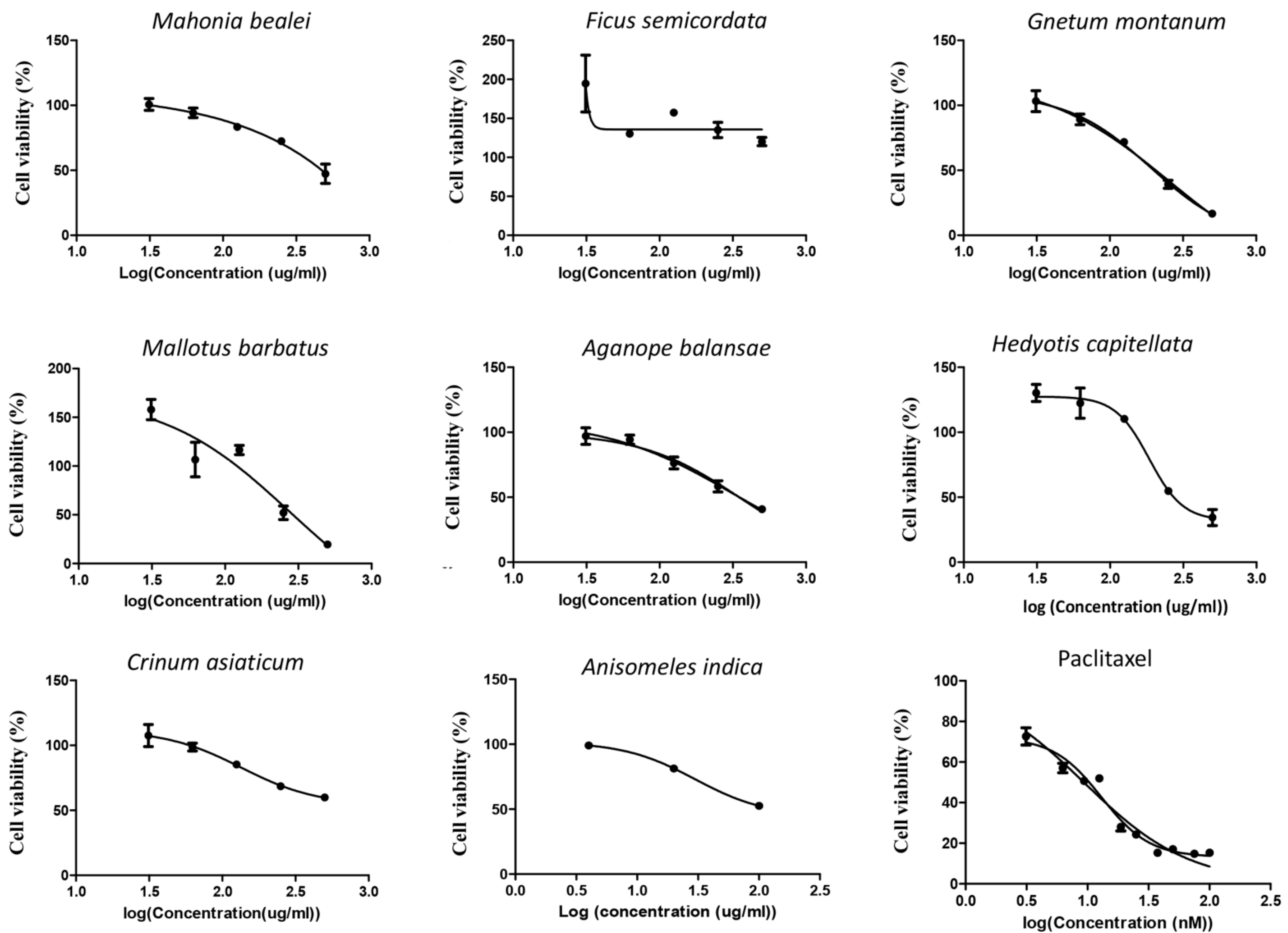
3.1. Anisomeles Indica Extract Displayed the Highest Cytotoxicity on Cervical Cancer HeLa Cells
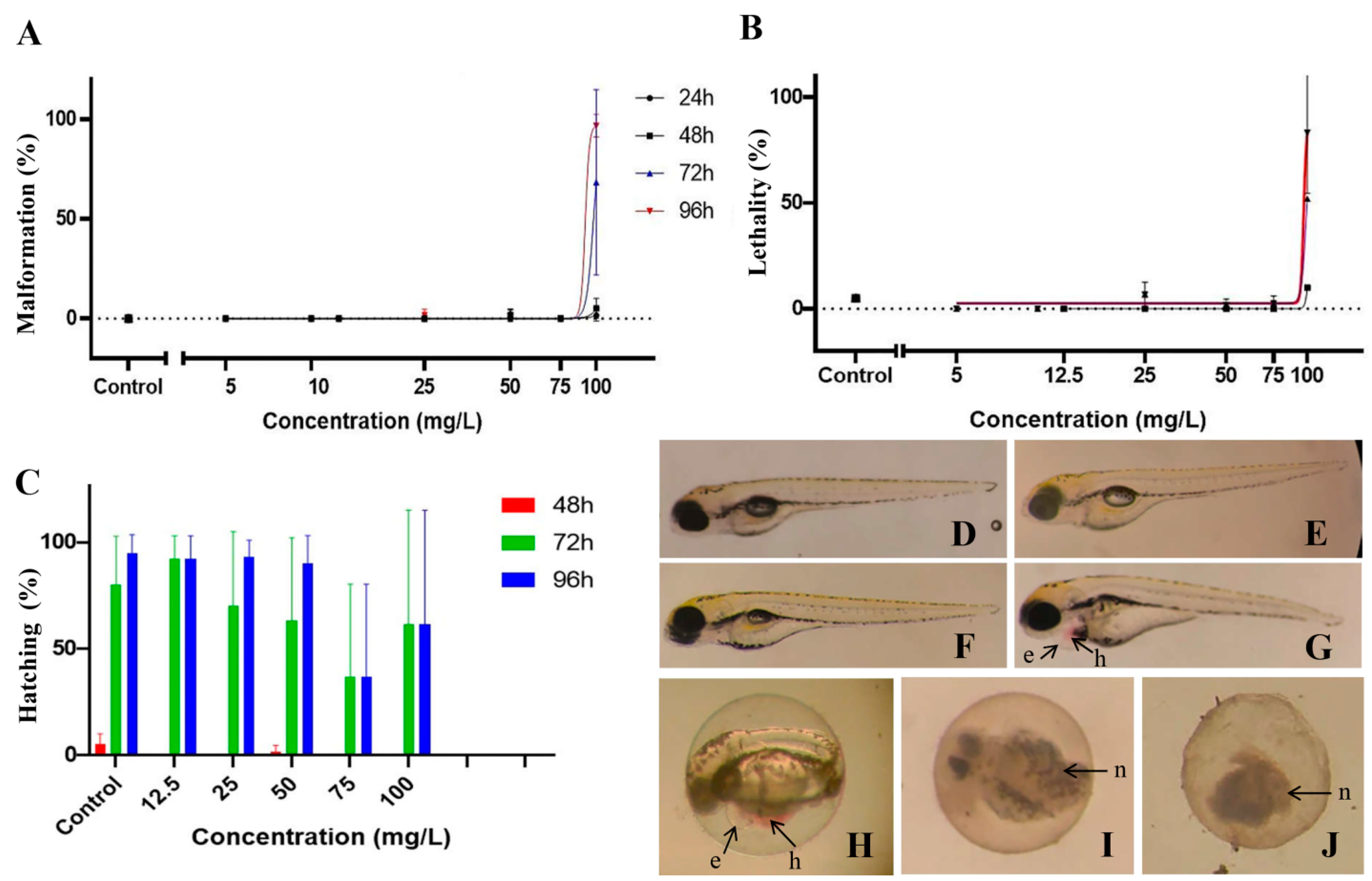
3.2. Toxicity of Anisomeles Indica Ethanol Extract on Zebrafish Embryos
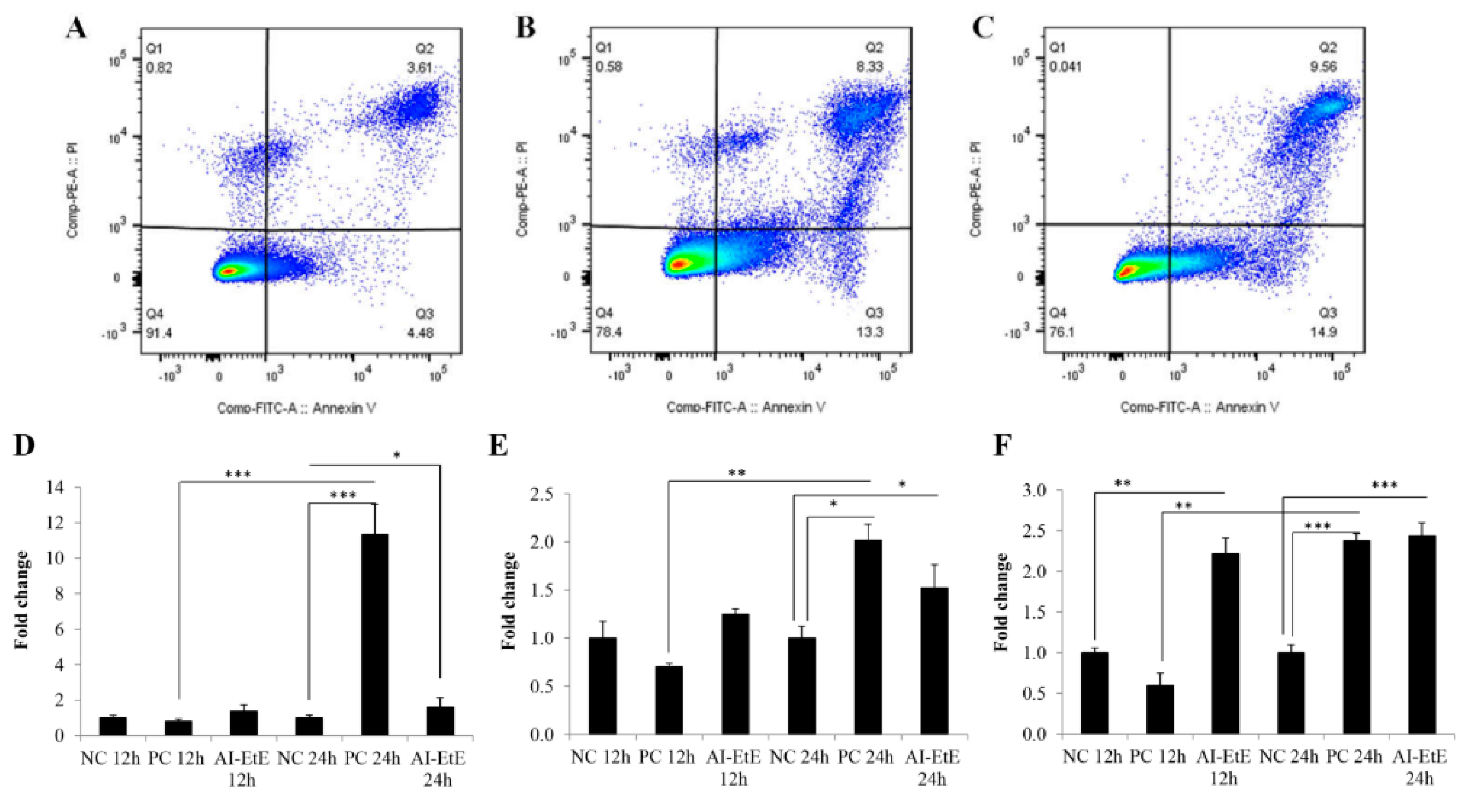
3.3. The Anisomeles Indica Ethanol Extract Induced HeLa Cell Apoptosis
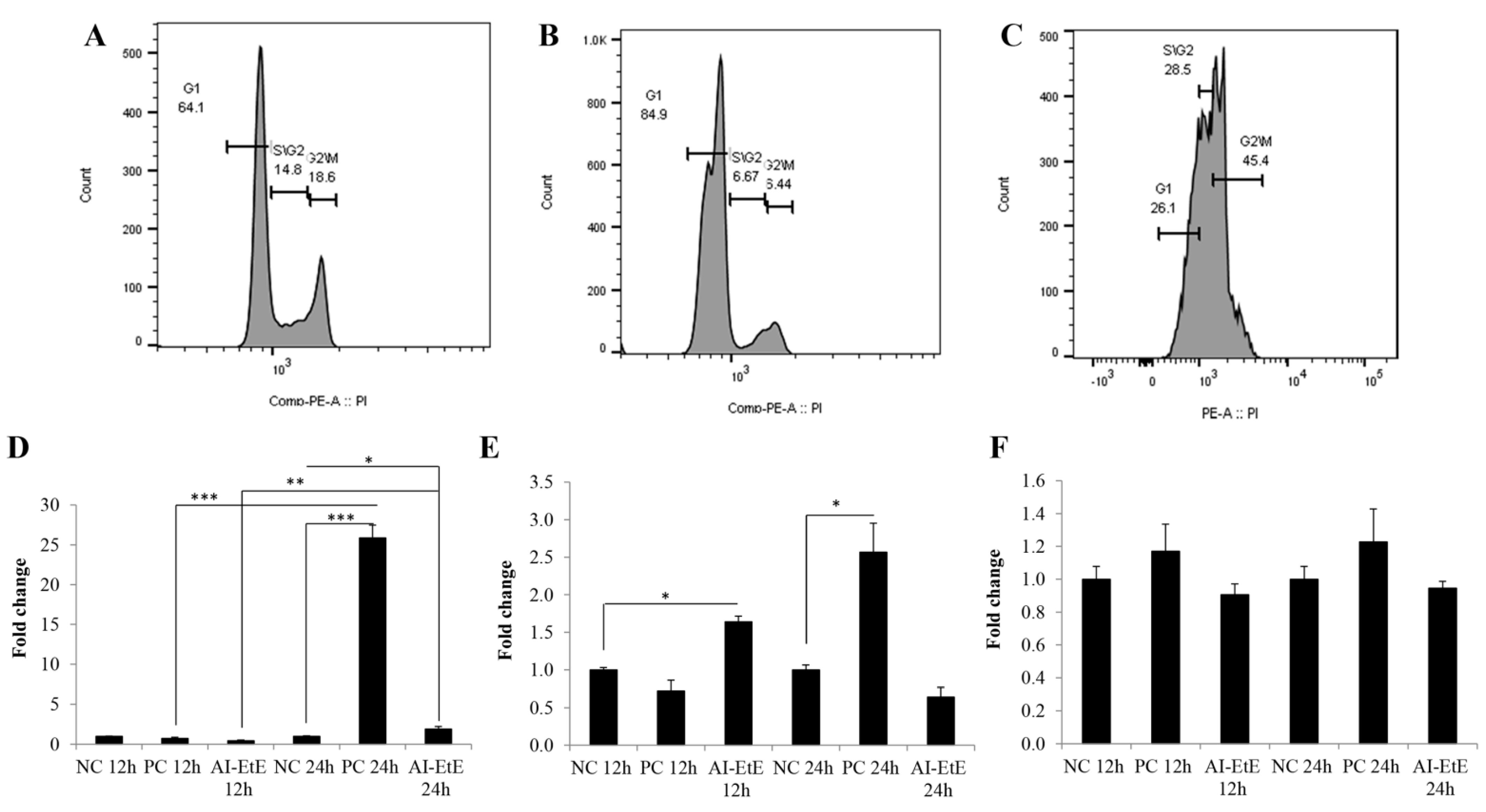
3.4. The Anisomeles Indica Ethanol Extract Induced HeLa Cell Cycle Arrest at the G1 Phase
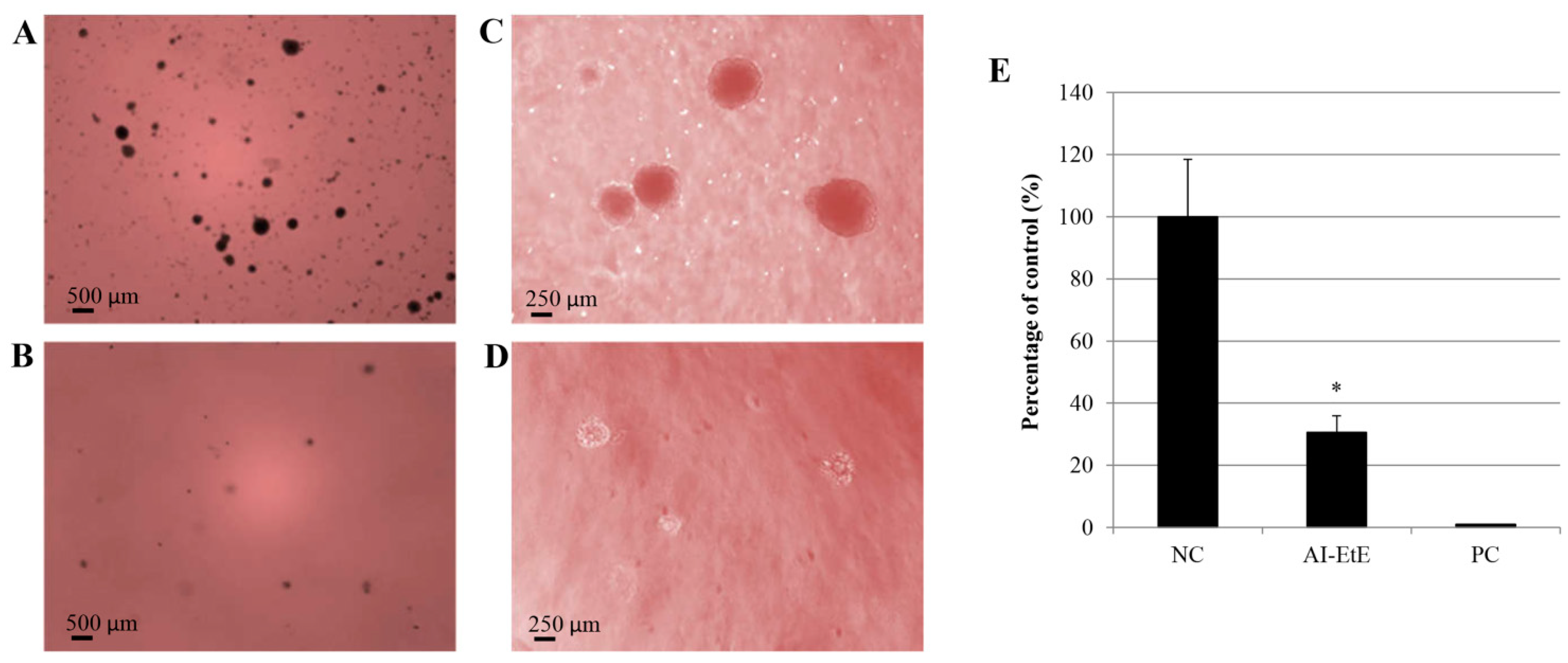
3.5. AI-EtE Decreased Anchorage-Independent Growth Ability of HeLa Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [Green Version]
- Kohler, D.R.; Goldspiel, B.R. Paclitaxel (taxol). Pharmacotherapy 1994, 14, 3–34. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Gill, N.S.; Chauhan, G.; Rana, A.C. An Overview about Versatile Molecule Capsaicin. Int. J. Pharm. Sci. Drug Res. 2001, 3, 280–286. [Google Scholar]
- Leonard, S.S.; Xia, C.; Jiang, B.H.; Stinefelt, B.; Klandorf, H.; Harris, G.K.; Shi, X. Resveratrol scavenges reactive oxygen species and effects radical-induced cellular responses. Biochem. Biophys. Res. Commun. 2003, 309, 1017–1026. [Google Scholar] [CrossRef]
- WHO. WHO Traditional Medicine Strategy: 2014–2023; WHO: Geneva, Switzerland, 2013; ISBN 978 92 4 150609 0. [Google Scholar]
- Shen, B. A New Golden Age of Natural Products Drug Discovery. Cell 2015, 163, 1297–1300. [Google Scholar] [CrossRef] [Green Version]
- WHO. Globocan 2012: Estimated Cancer Incidence, Mortality and Prevalence Worldwide in 2012. Available online: http://globocan.iarc.fr/Pages/online.aspx (accessed on 27 February 2021).
- World Health Organization. Human Papillomavirus (HPV) and Cervical Cancer. Available online: http://www.who.int/mediacentre/factsheets/fs380/en/ (accessed on 26 March 2018).
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef]
- Cavalcante, G.C.; Schaan, A.P.; Cabral, G.F.; Santana-da-Silva, M.N.; Pinto, P.; Vidal, A.F.; Ribeiro-Dos-Santos, Â. A Cell’s Fate: An Overview of the Molecular Biology and Genetics of Apoptosis. Int. J. Mol. Sci. 2019, 20, 4133. [Google Scholar] [CrossRef] [Green Version]
- Levine, A.J. p53, the cellular gatekeeper for growth and division. Cell 1997, 88, 323. [Google Scholar] [CrossRef] [Green Version]
- Vogelstein, B.; Lane, D.; Levine, A.J. Surfing the p53 network. Nature 2000, 408, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, M.L.; Taylor, W.R.; Chernov, M.V.; Chernova, O.B.; Stark, G.R. The p53 network. J. Biol. Chem. 1998, 273, 1. [Google Scholar] [CrossRef] [Green Version]
- Al-Khalaf, H.H.; Aboussekhra, A. p16(INK4A) positively regulates p21(WAF1) expression by suppressing AUF1-dependent mRNA decay. PLoS ONE 2013, 8, e70133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.H.; Kang, S.S.; Park, K.K.; Chang, H.W.; Magae, J.; Chang, Y.C. p53-independent induction of G1 arrest and p21WAF1/CIP1 expression by ascofuranone, an isoprenoid antibiotic, through downregulation of c-Myc. Mol. Cancer Ther. 2010, 9, 2102–2113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, C.J.; Lin, R.W.; Zhu, W.H.; Wen, T.K.; Hu, C.J.; Lee, Y.L.; Hung, T.I.; Wang, C. Transcription-independent and -dependent p53-mediated apoptosis in response to genotoxic and non-genotoxic stress. Cell Death. Discov. 2019, 5, 131. [Google Scholar] [CrossRef] [Green Version]
- Basappa, G.; Kumar, V.; Sarojini, B.K.; Poornima, D.V.; Gajula, H.; Sannabommaji, T.K.; Rajashekar, J. Chemical composition, biological properties of Anisomeles indica Kuntze essential oil. Ind. Crops Products 2015, 77, 89–96. [Google Scholar] [CrossRef]
- Junaid, S.; Dileep, N.; Rakesh, K.N.; Prashith, K.T.R.; Raghavendra, H.L. Contents of total phenolics and flavonoids, radical scavenging and anticaries activity of leaf and seed extract of Anisomeles indica Linn. Int. J. Drug. Dev. Res. 2013, 5, 286–292. [Google Scholar]
- Melkani, A.B.; Mohan, L.; Pant, C.C. Diterpene rich essential oil from Anisomeles indica (L.) o. kuntz. and its antimicrobial activity. World J. Pharma. Res. 2016, 5, 932–943. [Google Scholar]
- Yu, C.Y.; Jerry-Teng, C.L.; Hung, P.S.; Cheng, C.C.; Hsu, S.L.; Hwang, G.Y.; Tzeng, Y.M. Ovatodiolide isolated from Anisomeles indica induces cell cycle G2/M arrest and apoptosis via a ROS-dependent ATM/ATR signaling pathways. Eur. J. Pharmacol. 2018, 819, 16–29. [Google Scholar] [CrossRef]
- Lien, H.M.; Wang, C.Y.; Chang, H.Y.; Huang, C.L.; Peng, M.T.; Sing, Y.T.; Chen, C.C.; Lai, C.H. Bioevaluation of Anisomeles indica extracts and their inhibitory effects on Helicobacter pylori-mediated inflammation. J. Ethnopharmacol. 2013, 145, 397–401. [Google Scholar] [CrossRef]
- Hou, Y.Y.; Wu, M.L.; Hwang, Y.C.; Chang, F.R.; Wu, Y.C.; Wu, C.C. The natural diterpenoid ovatodiolide induces cell cycle arrest and apoptosis in human oral squamous cell carcinoma Ca9-22 cells. Life Sci. 2009, 85, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.F.; Rao, Y.K.; Tzeng, Y.M. Aqueous extract of Anisomeles indica and its purified compound exerts anti-metastatic activity through inhibition of NF-κB/AP-1-dependent MMP-9 activation in human breast cancer MCF-7 cells. Food Chem. Toxicol. 2012, 50, 2930–2936. [Google Scholar] [CrossRef] [PubMed]
- Lawson, N.D.; Weinstein, B.M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OECD Publishing. OECD Test No. 423: Acute Oral Toxicity—Acute Toxic Class Method; OECD Publishing: Paris, France, 2002. [Google Scholar]
- Wijesundara, N.M.; Rupasinghe, H. Bactericidal and Anti-Biofilm Activity of Ethanol Extracts Derived from Selected Medicinal Plants against Streptococcus pyogenes. Molecules 2019, 24, 1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Schutte, B.; Nuydens, R.; Geerts, H.; Ramaekers, F. Annexin V binding assay as a tool to measure apoptosis in differentiated neuronal cells. J. Neurosci. Methods 1998, 86, 63–69. [Google Scholar] [CrossRef]
- Darzynkiewicz, Z.; Huang, X.; Zhao, H. Analysis of Cellular DNA Content by Flow Cytometry. Curr. Protoc. Cytom. 2017, 82, 7.5.1–7.5.20. [Google Scholar]
- Ohgami, N.; Yamanoshita, O.; Thang, N.D.; Yajima, I.; Nakano, C.; Wenting, W.; Ohnuma, S.; Kato, M. Carcinogenic risk of chromium, copper and arsenic in CCA-treated wood. Environ. Pollut. 2015, 206, 456–460. [Google Scholar] [CrossRef]
- Thang, N.D.; Yajima, I.; Kumasaka, M.Y.; Kato, M. Bidirectional functions of arsenic as a carcinogen and an anti-cancer agent in human squamous cell carcinoma. PLoS ONE 2014, 9, e96945. [Google Scholar] [CrossRef]
- Lu, K.T.; Wang, B.Y.; Chi, W.Y.; Chang-Chien, J.; Yang, J.J.; Lee, H.T.; Tzeng, Y.M.; Chang, W.W. Ovatodiolide Inhibits Breast Cancer Stem/Progenitor Cells through SMURF2-Mediated Downregulation of Hsp27. Toxins 2016, 8, 127. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.L.; Tsai, P.C.; Hsieh, C.Y.; Chang, L.S.; Lin, S.R. Antimetastatic effect and mechanism of ovatodiolide in MDA-MB-231 human breast cancer cells. Chem. Biol. Interact. 2011, 194, 148–158. [Google Scholar] [CrossRef]
- Koster, R.; Timmer-Bosscha, H.; Bischoff, R.; Gietema, J.A.; de Jong, S. Disruption of the MDM2-p53 interaction strongly potentiates p53-dependent apoptosis in cisplatin-resistant human testicular carcinoma cells via the Fas/FasL pathway. Cell. Death Dis. 2011, 2, e148. [Google Scholar] [CrossRef] [Green Version]
- Lanni, J.S.; Lowe, S.W.; Licitra, E.J.; Liu, J.O.; Jacks, T. p53-independent apoptosis induced by paclitaxel through an indirect mechanism. Proc. Natl. Acad. Sci. USA 1997, 94, 9679–9683. [Google Scholar] [CrossRef] [Green Version]
- Leong, W.F.; Chau, J.F.; Li, B. p53 Deficiency leads to compensatory up-regulation of p16INK4a. Mol. Cancer Res. 2009, 7, 354–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pines, J. Cyclins: Wheels within wheels. Cell Growth Differ. 1991, 2, 305. [Google Scholar] [PubMed]
- Sherr, C.J. G1 phase progression: Cycling on cue. Cell 1994, 79, 551. [Google Scholar] [CrossRef]
- Ohtsubo, M.; Theodoras, A.M.; Schumacher, J.; Roberts, J.M.; Pagano, M. Human cyclin E, a nuclear protein essential for the G1-to-S phase transition. Mol. Cell. Biol. 1995, 15, 2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, F.; Strausfeld, U.; Fernandez, A.; Lamb, N.J. Cyclin A is required for the onset of DNA replication in mammalian fibroblasts. Cell 1991, 67, 1169. [Google Scholar] [CrossRef]
- Arellano, M.; Moreno, S. Regulation of CDK/cyclin complexes during the cell cycle. Int. J. Biochem. Cell Biol. 1997, 29, 559. [Google Scholar] [CrossRef]
- Ko, L.J.; Prives, C. p53: Puzzle and paradigm. Genes Dev. 1996, 10, 1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Zhao, J.; Du, Y.; Park, H.R.; Sun, S.Y.; Bernal-Mizrachi, L.; Aitken, A.; Khuri, F.R.; Fu, H. Down-regulation of 14-3-3zeta suppresses anchorage-independent growth of lung cancer cells through anoikis activation. Proc. Natl. Acad. Sci. USA 2008, 105, 162–167. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, F.; Hamanaka, R.; Wheaton, W.W.; Weinberg, S.; Joseph, J. Mitochondrial metabolism and ROS generation are essential for Kras-mediated tumorigenicity. Proc. Natl. Acad. Sci. USA 2010, 107, 8788–8793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.L.; Lan, Y.H.; Hsieh, P.W.; Wu, C.C.; Chen, S.L.; Yen, C.T.; Chang, F.R.; Hung, W.C.; Wu, Y.C. Bioactive cembrane diterpenoids of anisomeles indica. J. Nat. Prod. 2008, 71, 1207–1212. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; He, H.; Xue, J.; Wang, J.; Jin, H.; Wu, J.; Hu, J.; Wang, R.; Kuchta, K. Mori Ramulus (Chin.Ph.)-the Dried Twigs of Morus alba L./Part 1: Discovery of Two Novel Coumarin Glycosides from the Anti-Hyperuricemic Ethanol Extract. Molecules 2018, 24, 629. [Google Scholar] [CrossRef] [Green Version]
- Gori, A.; Ferrini, F.; Marzano, M.C.; Tattini, M.; Centritto, M.; Baratto, M.C.; Pogni, R.; Brunetti, C. Characterisation and Antioxidant Activity of Crude Extract and Polyphenolic Rich Fractions from C. incanus Leaves. Int. J. Mol. Sci. 2016, 17, 1344. [Google Scholar] [CrossRef]
- Adnan, M.; Oh, K.K.; Azad, M.O.K.; Shin, M.H.; Wang, M.H.; Cho, D.H. Kenaf (Hibiscus cannabinus L.) Leaves and Seed as a Potential Source of the Bioactive Compounds: Effects of Various Extraction Solvents on Biological Properties. Life 2020, 10, 223. [Google Scholar] [CrossRef] [PubMed]
- Attimarad, M.; Ahmed, K.K.; Aldhubaib, B.E.; Harsha, S. High-performance thin layer chromatography: A powerful analytical technique in pharmaceutical drug discovery. Pharm. Methods 2011, 2, 71–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razafindrakoto, Z.R.; Donno, D.; Tombozara, N.; Andriamaniraka, H.; Andrianjara, C.; Ramanitrahasimbola, D.; Beccaro, G.L. Antioxidant, Anti-Inflammatory, and Antidiabetic Activities of Leaves and Stems of Uapaca bojeri Bail. (EUPHORBIACEAE), an Endemic Plant of Madagascar. Pharmaceuticals 2020, 13, 71. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vietnamese Name | Scientific Name | Code in Museum | Place for Collection | Part Used | |
|---|---|---|---|---|---|
| 1 | Phòng phong thảo | Anisomeles indica | HNU 024777 | Bát xát, Lào Cai | Dry stem leaves |
| 2 | Hoàng liên ôro lá dày | Mahonia bealei | HNU 024779 | Bát xát, Lào Cai | Dry stem |
| 3 | Đa lá lệch | Ficus semicordata | HNU 024780 | Vị Xuyên, Hà Giang | Dry leaves |
| 4 | Gắm núi | Gnetum montanum | HNU 024781 | Vị Xuyên, Hà Giang | Dry stem |
| 5 | Náng hoa trắng | Crinum asiaticum | HNU 024783 | Cẩm Phả, Quảng Ninh | Dry leaves |
| 6 | Bùm bụp | Mallotus barbatus | HNU 024784 | Lạc Sơn, Hòa Bình | Dry root |
| 7 | Mạn mân | Aganope balansae | HNU 024785 | Bắc Quang, Hà Giang | Dry stem |
| 8 | Dạ cẩm | Hedyotis capitellata | HNU 024786 | Bắc Quang, Hà Giang | Fresh leaves |
| No | Sample | HeLa | MCF-7 | ||
|---|---|---|---|---|---|
| IC 50 (µg/mL) | R2 | IC 50 (µg/mL) | R2 | ||
| 1 | Anisomeles indica | 38.8 | 0.92 | 133 | 0.99 |
| 2 | Mahonia bealei | 101 | 0.84 | 254 | 0.90 |
| 3 | Ficus semicordata | 358 | 0.96 | 188 | 0.86 |
| 4 | Gnetum montanum | 150 | 0.93 | 257 | 0.96 |
| 5 | Crinum asiaticum | 69.5 | 0.93 | 215 | 0,95 |
| 6 | Mallotus barbatus | 498 | 0.98 | 276 | 0.84 |
| 7 | Aganope balansae | 228 | 0.71 | 310 | 0.91 |
| 8 | Hedyotis capitellata | 173 | 0.99 | 184 | 0.94 |
| Paclitaxel | 13.5 (ng/mL) | 0.96 | 15 (ng/mL) | 0.98 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bich-Loan, N.T.; Kien, K.T.; Thanh, N.L.; Kim-Thanh, N.T.; Huy, N.Q.; The-Hai, P.; Muller, M.; Nachtergael, A.; Duez, P.; Thang, N.D. Toxicity and Anti-Proliferative Properties of Anisomeles indica Ethanol Extract on Cervical Cancer HeLa Cells and Zebrafish Embryos. Life 2021, 11, 257. https://doi.org/10.3390/life11030257
Bich-Loan NT, Kien KT, Thanh NL, Kim-Thanh NT, Huy NQ, The-Hai P, Muller M, Nachtergael A, Duez P, Thang ND. Toxicity and Anti-Proliferative Properties of Anisomeles indica Ethanol Extract on Cervical Cancer HeLa Cells and Zebrafish Embryos. Life. 2021; 11(3):257. https://doi.org/10.3390/life11030257
Chicago/Turabian StyleBich-Loan, Nguyen T., Kieu Trung Kien, Nguyen Lai Thanh, Nguyen T. Kim-Thanh, Nguyen Quang Huy, Pham The-Hai, Marc Muller, Amandine Nachtergael, Pierre Duez, and Nguyen Dinh Thang. 2021. "Toxicity and Anti-Proliferative Properties of Anisomeles indica Ethanol Extract on Cervical Cancer HeLa Cells and Zebrafish Embryos" Life 11, no. 3: 257. https://doi.org/10.3390/life11030257