
Hexane Extract of Chloranthus japonicus Increases Adipocyte Differentiation by Acting on Wnt/β-Catenin Signaling Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
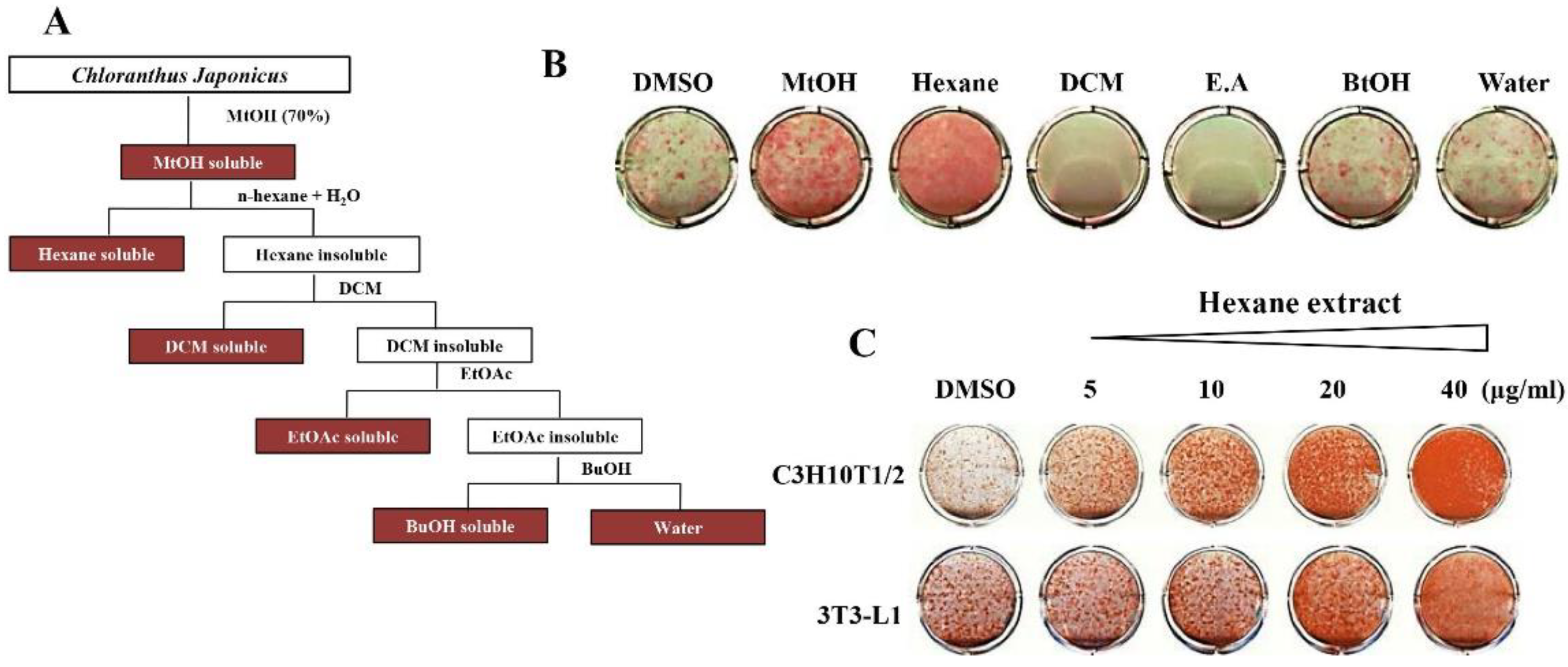
2.1. Chloranthus Japonicus Extraction
2.2. Cell Culture and Adipocyte Differentiation
2.3. mRNA Expression Analysis
2.4. Wnt Activation and Inhibition Studies
2.5. Statistical Analysis
3. Results
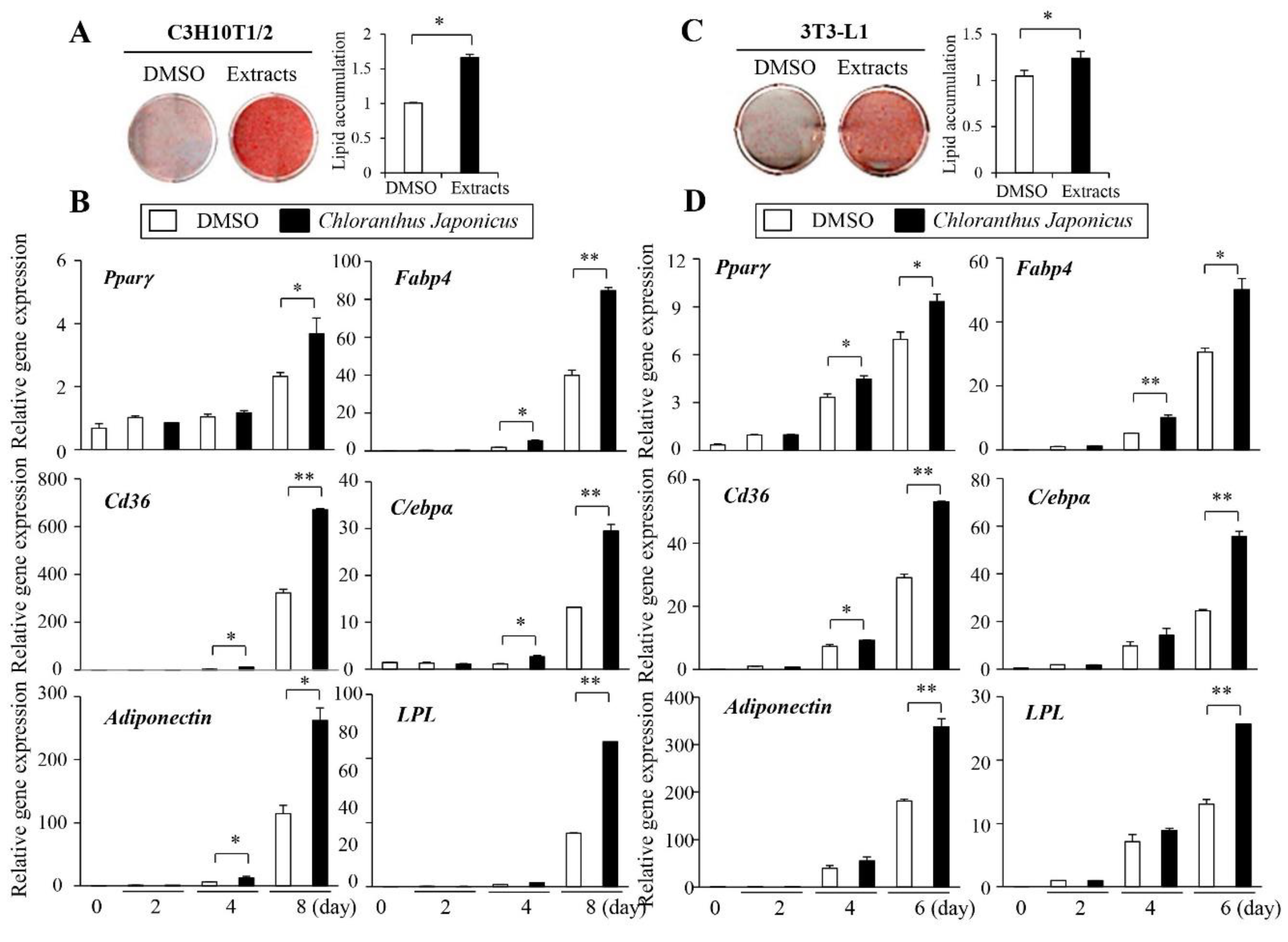
3.1. Chloranthus Japonicus Methanol Extract Promotes Adipogenesis in C3H10T1/2 and 3T3-L1 Cells
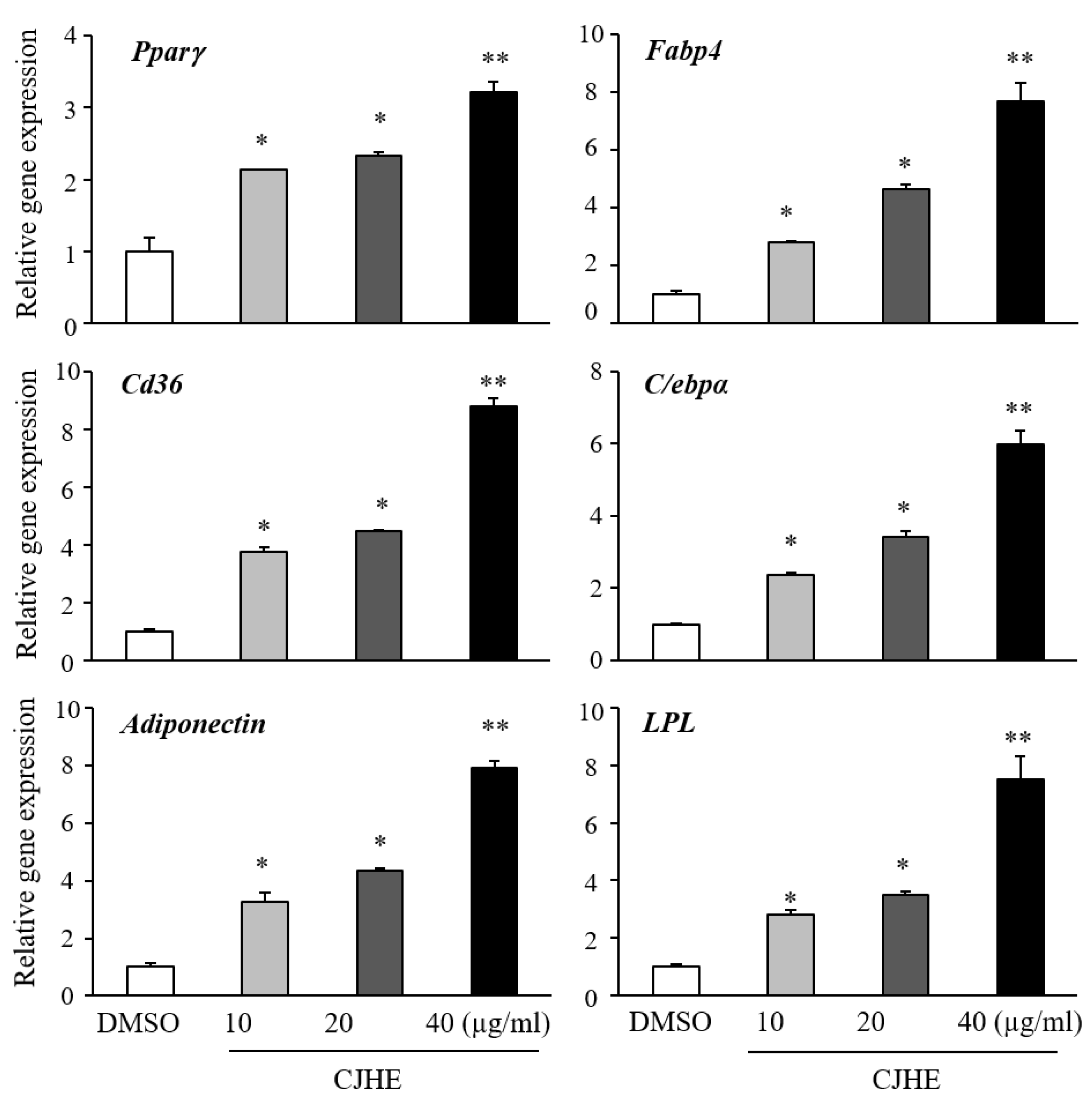
3.2. Chloranthus Japonicus Hexane Fraction Is the Most Potent Stimulator of Adipogenesis in C3H10T1/2 and 3T3-L1 Cells
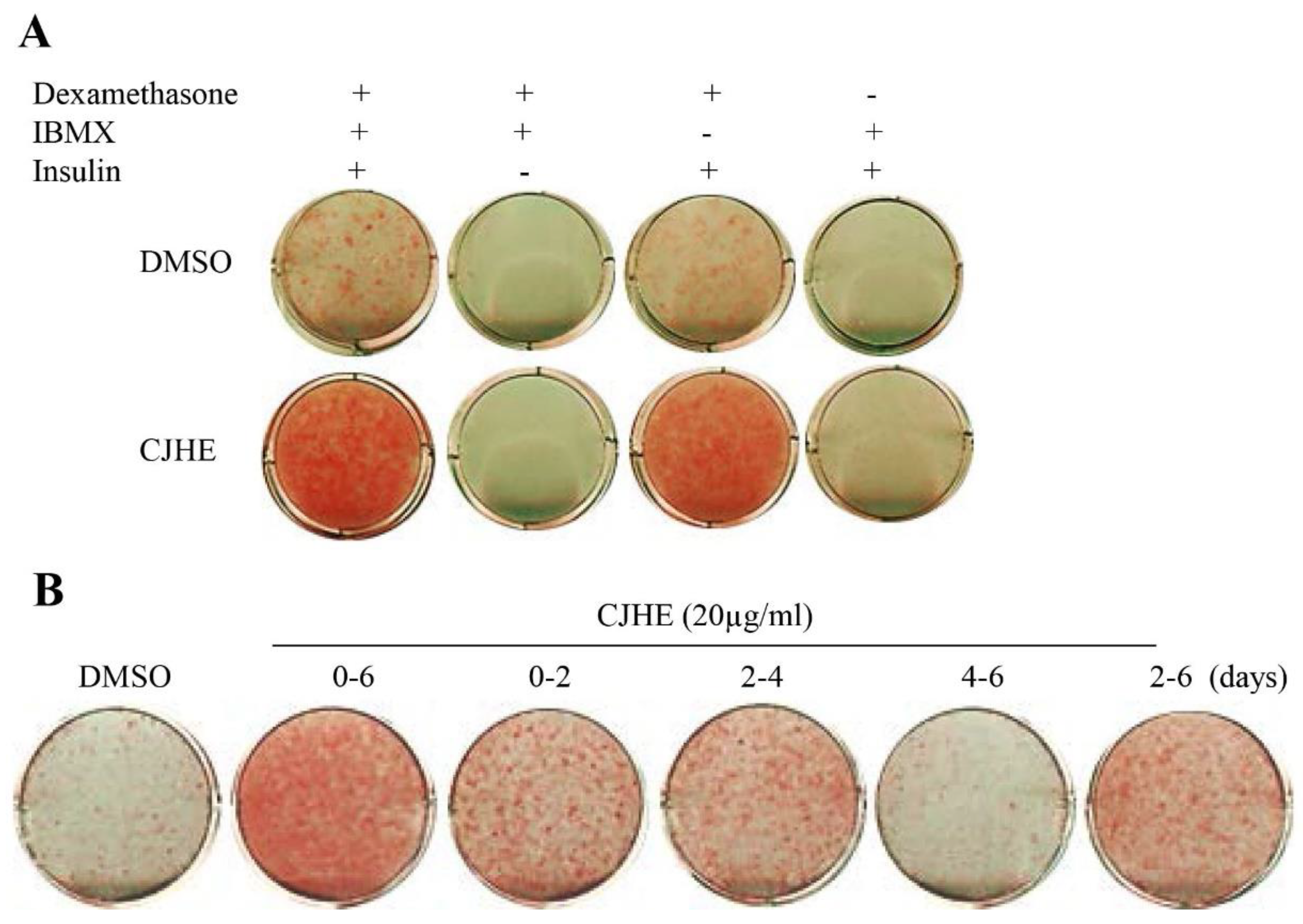
3.3. The Pro-Adipogenic Effect of CHJE Is Dependent upon Dexamethasone and Insulin
3.4. The Pro-Adipogenic Effect of CHJE Is Critical during the Early Adiogenic Stages
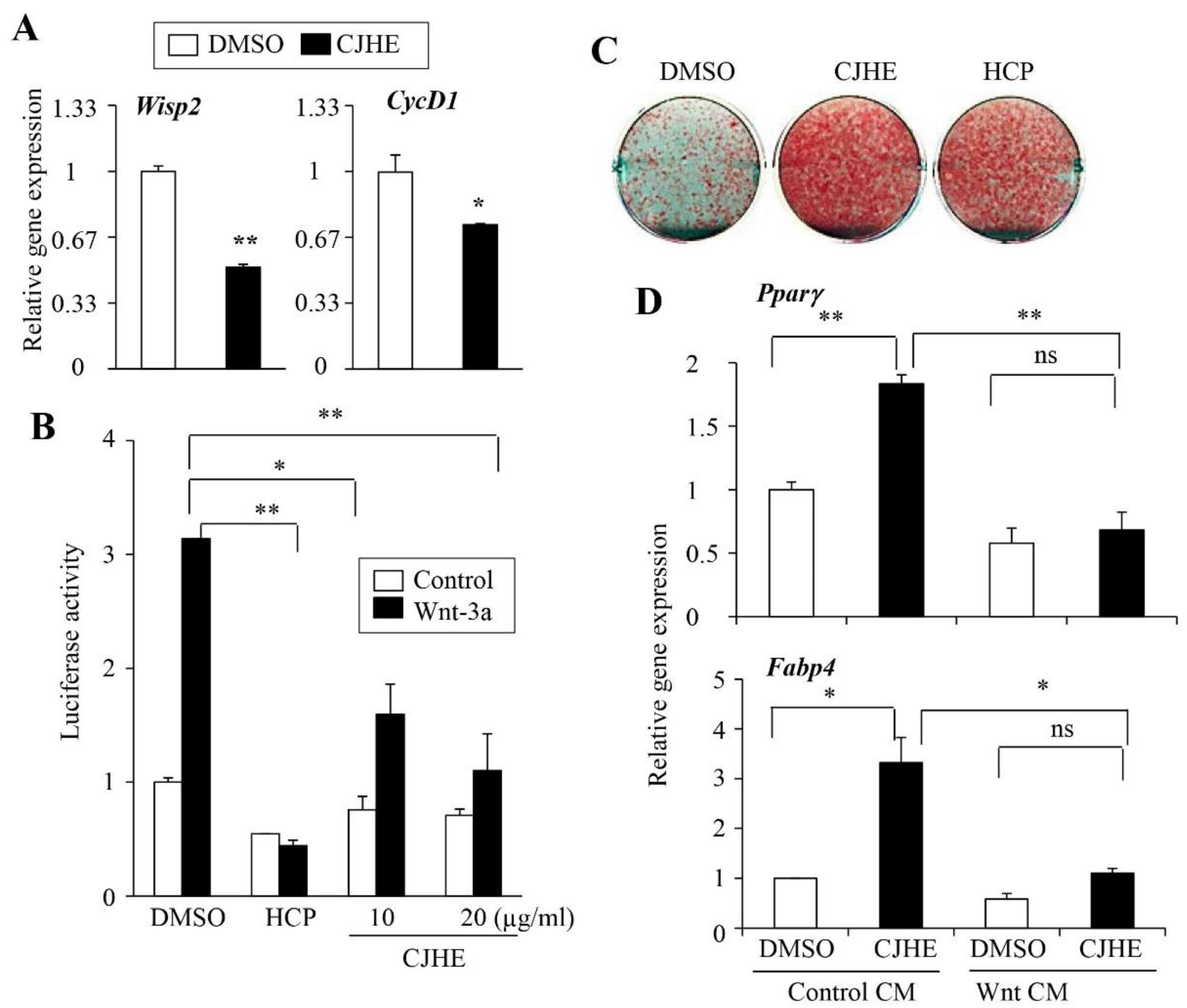
3.5. CJHE Inhibits the Wnt Pathway
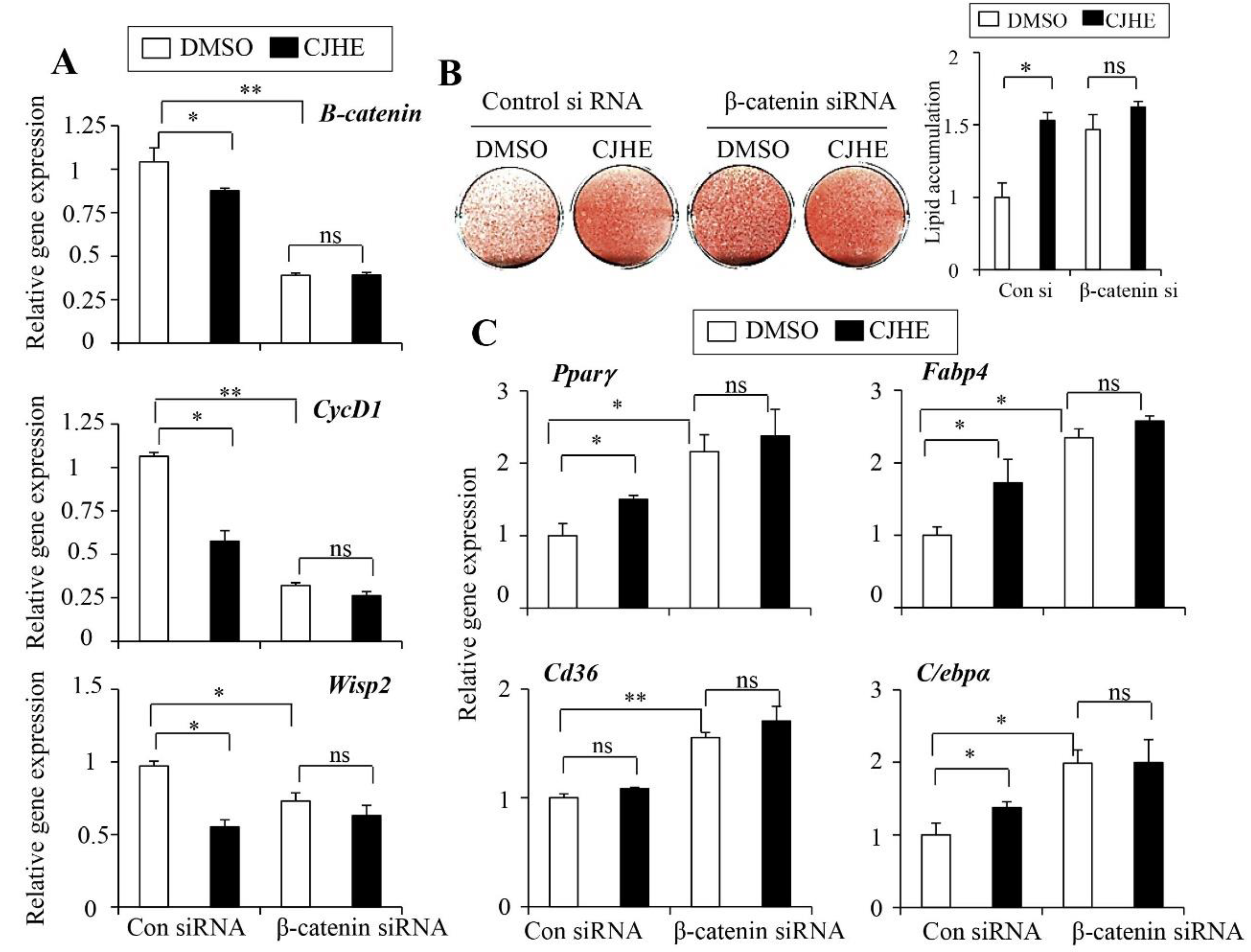
3.6. Wnt Signaling Is Essemtial for the Effects of CJHE
3.7. Differential Effects of the Extracts Generated from Different Plant Parts on Adipogenesis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tontonoz, P.; Spiegelman, B.M. Fat and beyond: The diverse biology of PPARgamma. Annu. Rev. Biochem. 2008, 77, 289–312. [Google Scholar]
- Tontonoz, P.; Hu, E.; Spiegelman, B.M. Stimulation of adipogenesis in fibroblasts by PPAR gamma 2, a lipid-activated transcription factor. Cell 1994, 79, 1147–1156. [Google Scholar]
- Hu, E.; Tontonoz, P.; Spiegelman, B.M. Transdifferentiation of myoblasts by the adipogenic transcription factors PPAR gamma and C/EBP alpha. Proc. Natl. Acad. Sci. USA 1995, 92, 9856–9860. [Google Scholar]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar]
- Hong, J.W.; Park, K.W. Further understanding of fat biology: Lessons from a fat fly. Exp. Mol. Med. 2010, 42, 12–20. [Google Scholar]
- Waki, H.; Tontonoz, P. Endocrine functions of adipose tissue. Annu. Rev. Pathol. 2007, 2, 31–56. [Google Scholar]
- Lazar, M.A. PPAR gamma, 10 years later. Biochimie 2005, 87, 9–13. [Google Scholar]
- Lehmann, J.M.; Moore, L.B.; Smith-Oliver, T.A.; Wilkison, W.O.; Willson, T.M.; Kliewer, S.A. An antidiabetic thiazolidinedione is a high affinity ligand for peroxisome proliferator-activated receptor gamma (PPAR gamma). J. Biol. Chem. 1995, 270, 12953–12956. [Google Scholar]
- Nissen, S.E.; Wolski, K. Effect of rosiglitazone on the risk of myocardial infarction and death from cardiovascular causes. N. Engl. J. Med. 2007, 356, 2457–2471. [Google Scholar]
- Graham, D.J.; Ouellet-Hellstrom, R.; MaCurdy, T.E.; Ali, F.; Sholley, C.; Worrall, C.; Kelman, J.A. Risk of acute myocardial infarction, stroke, heart failure, and death in elderly Medicare patients treated with rosiglitazone or pioglitazone. JAMA 2010, 304, 411–418. [Google Scholar]
- Choi, J.H.; Banks, A.S.; Estall, J.L.; Kajimura, S.; Bostrom, P.; Laznik, D.; Ruas, J.L.; Chalmers, M.J.; Kamenecka, T.M.; Bluher, M.; et al. Anti-diabetic drugs inhibit obesity-linked phosphorylation of PPARgamma by Cdk5. Nature 2010, 466, 451–456. [Google Scholar]
- Choi, J.H.; Banks, A.S.; Kamenecka, T.M.; Busby, S.A.; Chalmers, M.J.; Kumar, N.; Kuruvilla, D.S.; Shin, Y.; He, Y.; Bruning, J.B.; et al. Antidiabetic actions of a non-agonist PPARgamma ligand blocking Cdk5-mediated phosphorylation. Nature 2011, 477, 477–481. [Google Scholar]
- Hall, J.A.; Ramachandran, D.; Roh, H.C.; DiSpirito, J.R.; Belchior, T.; Zushin, P.H.; Palmer, C.; Hong, S.; Mina, A.I.; Liu, B.; et al. Obesity-Linked PPARgamma S273 Phosphorylation Promotes Insulin Resistance through Growth Differentiation Factor 3. Cell Metab. 2020, 32, 665–675.e666. [Google Scholar]
- Waki, H.; Park, K.W.; Mitro, N.; Pei, L.; Damoiseaux, R.; Wilpitz, D.C.; Reue, K.; Saez, E.; Tontonoz, P. The small molecule harmine is an antidiabetic cell-type-specific regulator of PPARgamma expression. Cell Metab. 2007, 5, 357–370. [Google Scholar]
- Mann, J. Natural products in cancer chemotherapy: Past, present and future. Nat. Rev. Cancer 2002, 2, 143–148. [Google Scholar]
- Xu, Y.J. Phytochemical and biological studies of Chloranthus medicinal plants. Chem. Biodivers. 2013, 10, 1754–1773. [Google Scholar]
- Wang, Q.H.; Kuang, H.X.; Yang, B.Y.; Xia, Y.G.; Wang, J.S.; Kong, L.Y. Sesquiterpenes from Chloranthus japonicus. J. Nat. Prod. 2011, 74, 16–20. [Google Scholar]
- Zhang, M.; Wang, J.S.; Wang, P.R.; Oyama, M.; Luo, J.; Ito, T.; Iinuma, M.; Kong, L.Y. Sesquiterpenes from the aerial part of Chloranthus japonicus and their cytotoxicities. Fitoterapia 2012, 83, 1604–1609. [Google Scholar]
- Zhao, J.J.; Guo, Y.Q.; Yang, D.P.; Xue, X.; Liu, Q.; Zhu, L.P.; Yin, S.; Zhao, Z.M. Chlojaponilactone B from Chloranthus japonicus: Suppression of Inflammatory Responses via Inhibition of the NF-kappaB Signaling Pathway. J. Nat. Prod. 2016, 79, 2257–2263. [Google Scholar]
- Hu, R.; Yan, H.; Hao, X.; Liu, H.; Wu, J. Shizukaol D isolated from Chloranthus japonicas inhibits AMPK-dependent lipid content in hepatic cells by inducing mitochondrial dysfunction. PLoS ONE 2013, 8, e73527. [Google Scholar]
- Song, N.J.; Yoon, H.J.; Kim, K.H.; Jung, S.R.; Jang, W.S.; Seo, C.R.; Lee, Y.M.; Kweon, D.H.; Hong, J.W.; Lee, J.S.; et al. Butein is a novel anti-adipogenic compound. J. Lipid Res. 2013, 54, 1385–1396. [Google Scholar]
- Chang, S.H.; Yun, U.J.; Choi, J.H.; Kim, S.; Lee, A.R.; Lee, D.H.; Seo, M.J.; Panic, V.; Villanueva, C.J.; Song, N.J.; et al. Identification of Phf16 and Pnpla3 as new adipogenic factors regulated by phytochemicals. J. Cell. Biochem. 2019, 120, 3599–3610. [Google Scholar]
- Park, K.W.; Waki, H.; Villanueva, C.J.; Monticelli, L.A.; Hong, C.; Kang, S.; MacDougald, O.A.; Goldrath, A.W.; Tontonoz, P. Inhibitor of DNA binding 2 is a small molecule-inducible modulator of peroxisome proliferator-activated receptor-gamma expression and adipocyte differentiation. Mol. Endocrinol. 2008, 22, 2038–2048. [Google Scholar]
- Kim, M.J.; Jang, W.S.; Lee, I.K.; Kim, J.K.; Seong, K.S.; Seo, C.R.; Song, N.J.; Bang, M.H.; Lee, Y.M.; Kim, H.R.; et al. Reciprocal regulation of adipocyte and osteoblast differentiation of mesenchymal stem cells by Eupatorium japonicum prevents bone loss and adiposity increase in osteoporotic rats. J. Med. Food 2014, 17, 772–781. [Google Scholar]
- Tang, L.; Zhu, H.; Yang, X.; Xie, F.; Peng, J.; Jiang, D.; Xie, J.; Qi, M.; Yu, L. Shizukaol D, a Dimeric Sesquiterpene Isolated from Chloranthus serratus, Represses the Growth of Human Liver Cancer Cells by Modulating Wnt Signalling Pathway. PLoS ONE 2016, 11, e0152012. [Google Scholar]
- Ross, S.E.; Hemati, N.; Longo, K.A.; Bennett, C.N.; Lucas, P.C.; Erickson, R.L.; MacDougald, O.A. Inhibition of adipogenesis by Wnt signaling. Science 2000, 289, 950–953. [Google Scholar]
- L’Episcopo, F.; Serapide, M.F.; Tirolo, C.; Testa, N.; Caniglia, S.; Morale, M.C.; Pluchino, S.; Marchetti, B. A Wnt1 regulated Frizzled-1/beta-Catenin signaling pathway as a candidate regulatory circuit controlling mesencephalic dopaminergic neuron-astrocyte crosstalk: Therapeutical relevance for neuron survival and neuroprotection. Mol. Neurodegener. 2011, 6, 49. [Google Scholar]
- Aires, A.; Fernandes, C.; Carvalho, R.; Bennett, R.N.; Saavedra, M.J.; Rosa, E.A. Seasonal effects on bioactive compounds and antioxidant capacity of six economically important brassica vegetables. Molecules 2011, 16, 6816–6832. [Google Scholar]
- Chew, A.L.; Jessica, J.J.; Sasidharan, S. Antioxidant and antibacterial activity of different parts of Leucas aspera. Asian Pac. J. Trop. Biomed. 2012, 2, 176–180. [Google Scholar]
- Hu, R.; Yan, H.; Fei, X.; Liu, H.; Wu, J. Modulation of glucose metabolism by a natural compound from Chloranthus japonicus via activation of AMP-activated protein kinase. Sci. Rep. 2017, 7, 778. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar]
- Blagodatski, A.; Klimenko, A.; Jia, L.; Katanaev, V.L. Small Molecule Wnt Pathway Modulators from Natural Sources: History, State of the Art and Perspectives. Cells 2020, 9, 589. [Google Scholar]
- Hwang, J.H.; Cha, P.H.; Han, G.; Bach, T.T.; Min do, S.; Choi, K.Y. Euodia sutchuenensis Dode extract stimulates osteoblast differentiation via Wnt/beta-catenin pathway activation. Exp. Mol. Med. 2015, 47, e152. [Google Scholar]
- Ishibashi, M. Screening for natural products that affect Wnt signaling activity. J. Nat. Med. 2019, 73, 697–705. [Google Scholar]
- Baek, K.; Park, H.J.; Hwang, H.R.; Baek, J.H. Propranolol attenuates calorie restriction- and high calorie diet-induced bone marrow adiposity. BMB Rep. 2014, 47, 587–592. [Google Scholar]
- Vougogiannopoulou, K.; Skaltsounis, A.L. From Tyrian purple to kinase modulators: Naturally halogenated indirubins and synthetic analogues. Planta Med. 2012, 78, 1515–1528. [Google Scholar]
- Meijer, L.; Skaltsounis, A.L.; Magiatis, P.; Polychronopoulos, P.; Knockaert, M.; Leost, M.; Ryan, X.P.; Vonica, C.A.; Brivanlou, A.; Dajani, R.; et al. GSK-3-selective inhibitors derived from Tyrian purple indirubins. Chem. Biol. 2003, 10, 1255–1266. [Google Scholar]
- Tapia-Rojas, C.; Schuller, A.; Lindsay, C.B.; Ureta, R.C.; Mejias-Reyes, C.; Hancke, J.; Melo, F.; Inestrosa, N.C. Andrographolide activates the canonical Wnt signalling pathway by a mechanism that implicates the non-ATP competitive inhibition of GSK-3beta: Autoregulation of GSK-3beta in vivo. Biochem. J. 2015, 466, 415–430. [Google Scholar]
- Li, Y.; Xu, J.; Chen, Y.; Mei, Z.; Xiao, Y. Screening of inhibitors of glycogen synthase kinase-3beta from traditional Chinese medicines using enzyme-immobilized magnetic beads combined with high-performance liquid chromatography. J. Chromatogr. A. 2015, 1425, 8–16. [Google Scholar]
- Ahn, J.; Lee, H.; Kim, S.; Ha, T. Curcumin-induced suppression of adipogenic differentiation is accompanied by activation of Wnt/beta-catenin signaling. Am. J. Physiol. Cell Physiol. 2010, 298, C1510–C1516. [Google Scholar]
- Rahman, N.; Jeon, M.; Kim, Y.S. Delphinidin, a major anthocyanin, inhibits 3T3-L1 pre-adipocyte differentiation through activation of Wnt/beta-catenin signaling. Biofactors 2016, 42, 49–59. [Google Scholar]
- Jang, Y.J.; Son, H.J.; Ahn, J.; Jung, C.H.; Ha, T. Coumestrol modulates Akt and Wnt/beta-catenin signaling during the attenuation of adipogenesis. Food Funct. 2016, 7, 4984–4991. [Google Scholar]
- Lee, H.; Bae, S.; Yoon, Y. The anti-adipogenic effects of (-)epigallocatechin gallate are dependent on the WNT/beta-catenin pathway. J. Nutr. Biochem. 2013, 24, 1232–1240. [Google Scholar]
- Galvis, A.; Marcano, A.; Stefancin, C.; Villaverde, N.; Priestap, H.A.; Tonn, C.E.; Lopez, L.A.; Barbieri, M.A. The effect of dehydroleucodine in adipocyte differentiation. Eur. J. Pharmacol. 2011, 671, 18–25. [Google Scholar]
- Bu, S.; Yuan, C.Y.; Xue, Q.; Chen, Y.; Cao, F. Bilobalide Suppresses Adipogenesis in 3T3-L1 Adipocytes via the AMPK Signaling Pathway. Molecules 2019, 24, 3503. [Google Scholar]
- Kwak, S.H.; Kim, Y.H. Zaluzanin C Inhibits Differentiation of 3T3-L1 Preadipocytes into Mature Adipocytes. J. Obes. Metab. Syndr. 2019, 28, 105–111. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yun, U.J.; Nho, C.W.; Park, K.W.; Yang, D.K. Hexane Extract of Chloranthus japonicus Increases Adipocyte Differentiation by Acting on Wnt/β-Catenin Signaling Pathway. Life 2021, 11, 241. https://doi.org/10.3390/life11030241
Yun UJ, Nho CW, Park KW, Yang DK. Hexane Extract of Chloranthus japonicus Increases Adipocyte Differentiation by Acting on Wnt/β-Catenin Signaling Pathway. Life. 2021; 11(3):241. https://doi.org/10.3390/life11030241
Chicago/Turabian StyleYun, Ui Jeong, Chu Won Nho, Kye Won Park, and Dong Kwon Yang. 2021. "Hexane Extract of Chloranthus japonicus Increases Adipocyte Differentiation by Acting on Wnt/β-Catenin Signaling Pathway" Life 11, no. 3: 241. https://doi.org/10.3390/life11030241