Phycocyanin from Arthrospira platensis as Potential Anti-Cancer Drug: Review of In Vitro and In Vivo Studies

 ,
,
Abstract
:1. Introduction
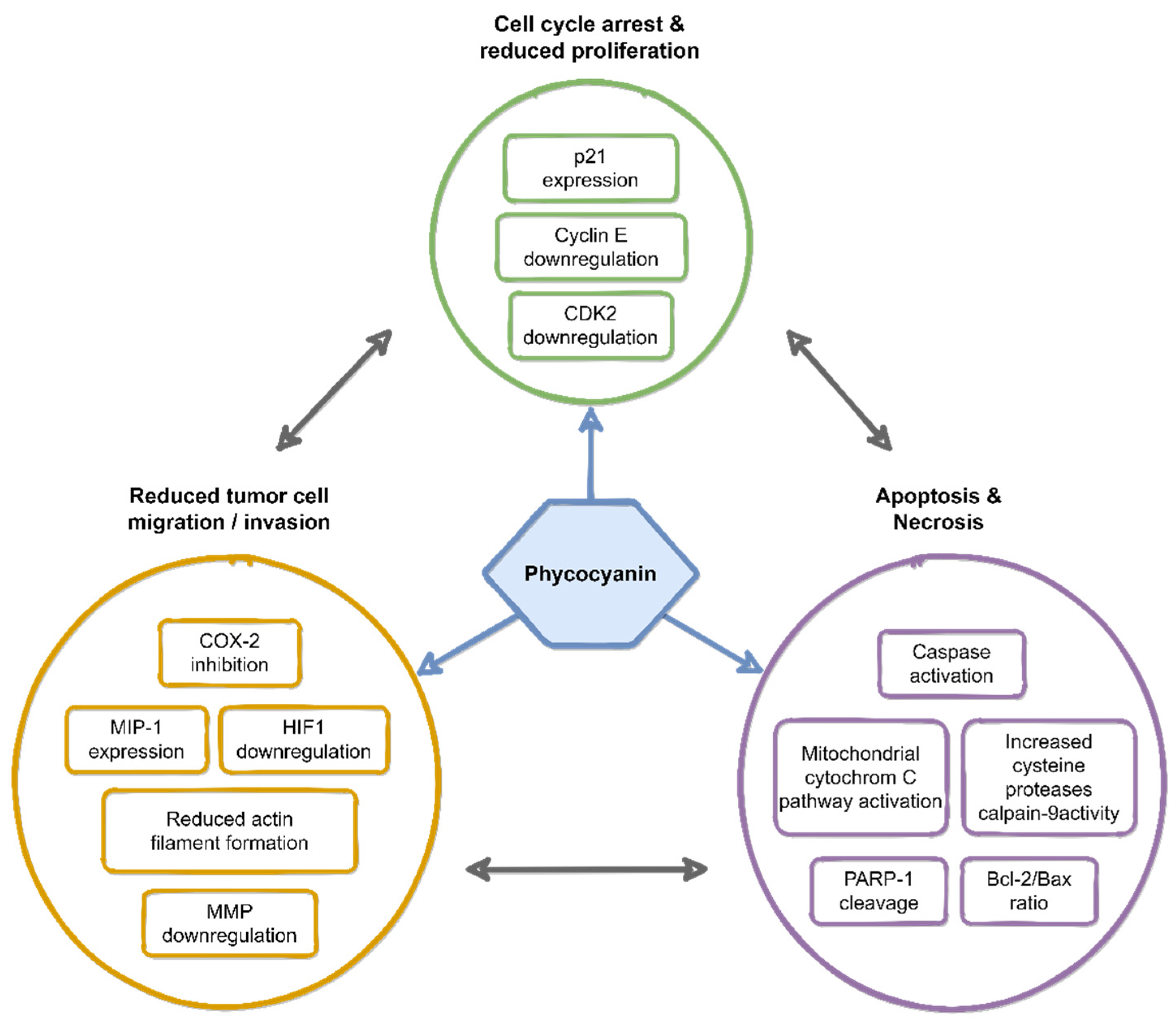
2. Anti-Cancer Effects of Phycocyanin
3. Molecular Mechanisms of Phycocyanin-Induced Cell Death in Tumor Cells
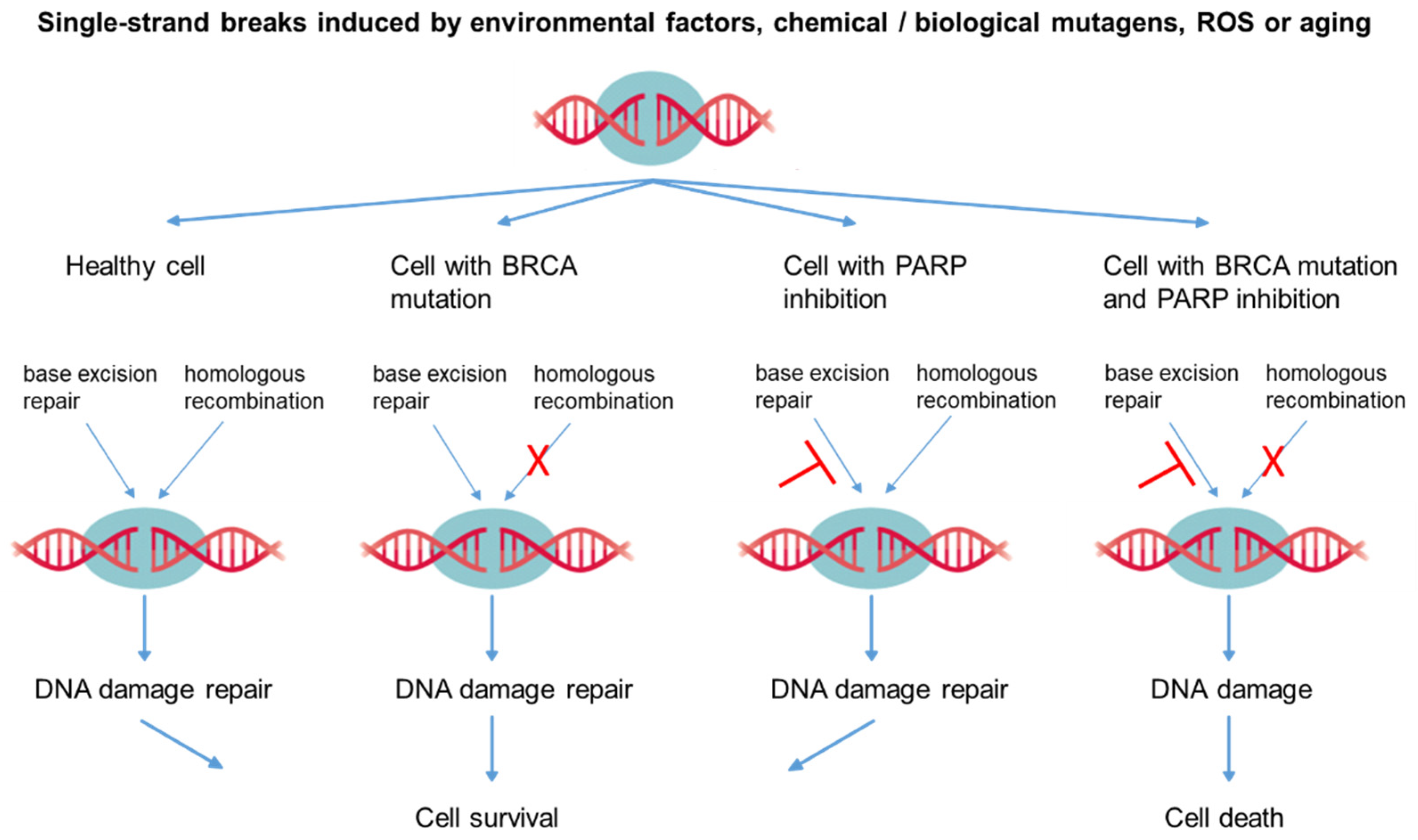
4. Strategies for Potential Phycocyanin-Based Anti-Cancer Therapies

{kind=link}
{kind=link}
{kind=link}
| Tumor Type | Tumor Induction | Animal Model | Combination Drug/Technique | Phycocyanin Concentration | Treatment Duration | Ref. |
|---|---|---|---|---|---|---|
| Lung | Injection of A549 cells (right flank) | Rat (nude) | Betaine | 370.0 mg/kg of body weight per day, food supplement | 28 days | [51] |
| Lung | Injection of A549 cells (armpit area) | Mouse (nude) | All-trans retinoic acid | 0.2 mL (320 mg/mL) per day, injected in tumor area | 10 days | [52,97] |
| Colon | Injection of 1,2-dimethyl-hydrazine dihydro-chloride (subcutaneous) | Rat, Sprague- Dawley | Piroxicam | Up to 200 mg/kg body weight per day, food suppl. | 42 days | [77,98,99] |
| Ehrlich ascites carcinoma (EAC) | Injection of EAC cells (peritoneum) | Mouse, Swiss albino | Cisplatin | 0.5 g/kg body weight of AP, food suppl. | 14 days pre and 14 days post inocul. | [100] |
| Esophag. Squamous cell carc. | ESCC EC9706 | Mouse | Injected in tumor area | [101] | ||
| Cervix | Injection of SiHa/HeLa cells (axillary fossa/ armpit area) | Mouse, BALB/c | Nanoparticles functionalized: CD95sp and PC | Injected in tumor area/tail vein once every 2 days | 20 days | [57,102,103] |
| Liver | Injection of H22 cells (armpit area) | Mouse, BALB/c | Photodynamic therapy (PDT) | 0.02 mL (10 mg/mL) per day injected in tumor area | 10 days | [96] |
| Breast | Injection of MCF-7 cells (spleen area) | Mouse, BALB/c | PDT | 2 mL (320 mg/mL) per day | 13 days | [48] |
| Breast | Injection of MCF-7 cells (right abdomen) | Mouse, BALB/c | PDT, Nanoparticle coated with hematoporphyrin mono-methyl ether and PC | 100 µL nanoparticle solution (3 µg/g) injected in tumor, per day, once every 2 days | 14 days | [95] |
| Breast | Injection of 4T1 cells (subcutaneous) | Mouse, BALB/c | PDT, Nanoparticles functionalized with PC | 100 µL nanoparticle solution (cor. 150 µg/mL PC) | 14 days | [94] |
5. Effect of Phycocyanin on Tumor Cells in Comparison to Non-Malignant Cells
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. Impact of Natural Products on Developing New Anti-Cancer Agents. Chem. Rev. 2009, 109, 3012–3043. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Oh, K.K.; Azad, O.K.; Shin, M.H.; Wang, M.-H.; Cho, D.H. Kenaf (Hibiscus cannabinus L.) Leaves and Seed as a Potential Source of the Bioactive Compounds: Effects of Various Extraction Solvents on Biological Properties. Life 2020, 10, 223. [Google Scholar] [CrossRef] [PubMed]
- Hua, P.; Yu, Z.; Xiong, Y.; Liu, B.; Zhao, L. Regulatory Efficacy of Spirulina platensis Protease Hydrolyzate on Lipid Metabolism and Gut Microbiota in High-Fat Diet-Fed Rats. Int. J. Mol. Sci. 2018, 19, 4023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorczyca, W.; Gong, J.; Ardelt, B.; Traganos, F.; Darzynkiewicz, Z. The cell cycle related differences in susceptibility of HL-60 cells to apoptosis induced by various antitumor agents. Cancer Res. 1993, 53, 3186–3192. [Google Scholar] [PubMed]
- Sapp, J. The Prokaryote-Eukaryote Dichotomy: Meanings and Mythology. Microbiol. Mol. Biol. Rev. 2005, 69, 292–305. [Google Scholar] [CrossRef] [Green Version]
- El-Sheekh, M.M.; Hamad, S.M.; Gomaa, M. Protective effects of Spirulina on the liver function and hyperlipidemia of rats and human. Braz. Arch. Biol. Technol. 2014, 57, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Serban, M.-C.; Sahebkar, A.; Dragan, S.; Stoichescu-Hogea, G.; Ursoniu, S.; Andrica, F.-M.; Banach, M. A systematic review and meta-analysis of the impact of Spirulina supplementation on plasma lipid concentrations. Clin. Nutr. 2016, 35, 842–851. [Google Scholar] [CrossRef]
- Chamorro, G.; Salazar, M.; Favila, L.; Bourges, H. Pharmacology and toxicology of Spirulina alga. Rev. Investig. Clin. 1996, 48, 389–399. [Google Scholar]
- Simpore, J.; Kabore, F.; Zongo, F.; Dansou, D.; Bere, A.; Pignatelli, S.; Biondi, D.M.; Ruberto, G.; Musumeci, S. Nutrition rehabilitation of undernourished children utilizing Spiruline and Misola. Nutr. J. 2006, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Salazar, M.; Martínez, E.; Madrigal, E.; Ruiz, L.; Chamorro, G. Subchronic toxicity study in mice fed Spirulina maxima. J. Ethnopharmacol. 1998, 62, 235–241. [Google Scholar] [CrossRef]
- Marles, R.J.; Barrett, M.L.; Barnes, J.; Chavez, M.L.; Gardiner, P.; Ko, R.; Mahady, G.B.; Dog, T.L.; Sarma, N.D.; Giancaspro, G.I.; et al. United States Pharmacopeia Safety Evaluation of Spirulina. Crit. Rev. Food Sci. Nutr. 2011, 51, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Mysliwa-Kurdziel, B. Phycobilins and Phycobiliproteins Used in Food Industry and Medicine. Mini-Rev. Med. Chem. 2017, 17, 1173–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Li, Y.; Pakpour, S.; Wang, S.; Pan, Z.; Liu, J.; Wei, Q.; She, J.; Cang, H.; Zhang, R. Dose Effects of Orally Administered Spirulina Suspension on Colonic Microbiota in Healthy Mice. Front. Cell. Infect. Microbiol. 2019, 9, 243. [Google Scholar] [CrossRef]
- Yu, T.; Wang, Y.; Chen, X.; Xiong, W.; Tang, Y.; Lin, L. Spirulina platensis alleviates chronic inflammation with modulation of gut microbiota and intestinal permeability in rats fed a high-fat diet. J. Cell. Mol. Med. 2020, 24, 8603–8613. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Li, W.; Zhu, L.; Zhai, S.; Qin, S.; Du, Z. Effects of phycocyanin in modulating the intestinal microbiota of mice. MicrobiologyOpen 2019, 8, e00825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Lu, L.; Liu, B.; Qin, S. Effects of phycocyanin on pulmonary and gut microbiota in a radiation-induced pulmonary fibrosis model. Biomed. Pharmacother. 2020, 132, 110826. [Google Scholar] [CrossRef]
- Chacón-Lee, T.; González-Mariño, G. Microalgae for “Healthy” Foods-Possibilities and Challenges. Compr. Rev. Food Sci. Food Saf. 2010, 9, 655–675. [Google Scholar] [CrossRef]
- Deng, R.; Chow, T.-J. Hypolipidemic, Antioxidant, and Antiinflammatory Activities of Microalgae Spirulina. Cardiovasc. Ther. 2010, 28, e33–e45. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Hermosillo, A.; Torres-Durán, P.V.; Juárez-Oropeza, M.A. Hepatoprotective effects of Spirulina maxima in patients with non-alcoholic fatty liver disease: A case series. J. Med Case Rep. 2010, 4, 103. [Google Scholar] [CrossRef] [Green Version]
- Ismail, M.; Hossain, M.; Tanu, A.R.; Shekhar, H.U. Effect of Spirulina Intervention on Oxidative Stress, Antioxidant Status, and Lipid Profile in Chronic Obstructive Pulmonary Disease Patients. BioMed Res. Int. 2015, 2015, 486120. [Google Scholar] [CrossRef] [Green Version]
- Kalafati, M.; Jamurtas, A.Z.; Nikolaidis, M.G.; Paschalis, V.; Theodorou, A.A.; Sakellariou, G.K.; Koutedakis, Y.; Kouretas, D. Ergogenic and Antioxidant Effects of Spirulina Supplementation in Humans. Med. Sci. Sports Exerc. 2010, 42, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Park, J.-E.; Choi, Y.-J.; Huh, K.-B.; Kim, W.Y. A randomized study to establish the effects of spirulina in type 2 diabetes mellitus patients. Nutr. Res. Pr. 2008, 2, 295–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazokopakis, E.E.; Starakis, I.K.; Papadomanolaki, M.G.; Mavroeidi, N.G.; Ganotakis, E.S. The hypolipidaemic effects of Spirulina (Arthrospira platensis) supplementation in a Cretan population: A prospective study. J. Sci. Food Agric. 2014, 94, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, Y.J.; Ryu, H.K.; Kim, M.-H.; Chung, H.W.; Kim, W.Y. A Randomized Double-Blind, Placebo-Controlled Study to Establish the Effects of Spirulina in Elderly Koreans. Ann. Nutr. Metab. 2008, 52, 322–328. [Google Scholar] [CrossRef]
- Choi, W.Y.; Lee, H.Y. Effect of Ultrasonic Extraction on Production and Structural Changes of C-Phycocyanin from Marine Spirulina maxima. Int. J. Mol. Sci. 2018, 19, 220. [Google Scholar] [CrossRef] [Green Version]
- Challouf, R.; Trabelsi, L.; Ben Dhieb, R.; El Abed, O.; Yahia, A.; Ghozzi, K.; Ben Ammar, J.; Omran, H.; Ben Ouada, H. Evaluation of cytotoxicity and biological activities in extracellular polysaccharides released by cyanobacterium Arthrospira platensis. Braz. Arch. Biol. Technol. 2011, 54, 831–838. [Google Scholar] [CrossRef]
- Vaňková, K.; Marková, I.; Jašprová, J.; Dvořák, A.; Subhanová, I.; Zelenka, J.; Novosádová, I.; Rasl, J.; Vomastek, T.; Sobotka, R.; et al. Chlorophyll-Mediated Changes in the Redox Status of Pancreatic Cancer Cells Are Associated with Its Anticancer Effects. Oxidative Med. Cell. Longev. 2018, 2018, 4069167. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Chen, Y.; Qu, C.; Pan, Z.; Qin, Y.; Zhang, X.; Liu, W.; Li, D.; Zheng, Q. Vitamin C induces human melanoma A375 cell apoptosis via Bax- and Bcl-2-mediated mitochondrial pathways. Oncol. Lett. 2019, 18, 3880–3886. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.N.; Sonani, R.R.; Jakharia, K.; Bhastana, B.; Patel, H.M.; Chaubey, M.G.; Singh, N.K.; Madamwar, D. Antioxidant activity and associated structural attributes of Halomicronema phycoerythrin. Int. J. Biol. Macromol. 2018, 111, 359–369. [Google Scholar] [CrossRef]
- Lee, J.-H.; Tachibana, H.; Morinaga, Y.; Fujimura, Y.; Yamada, K. Modulation of proliferation and differentiation of C2C12 skeletal muscle cells by fatty acids. Life Sci. 2009, 84, 415–420. [Google Scholar] [CrossRef]
- Jiang, Q.; Elson-Schwab, I.; Courtemanche, C.; Ames, B.N. γ-Tocopherol and its major metabolite, in contrast to α-tocopherol, inhibit cyclooxygenase activity in macrophages and epithelial cells. Proc. Natl. Acad. Sci. USA 2000, 97, 11494–11499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, S.; Yan, Y.; Li, S.; Zhao, L.; Zhang, C.; Liu, L.; Wang, C. The In Vitro Anti-Tumor Activity of Phycocyanin against Non-Small Cell Lung Cancer Cells. Mar. Drugs 2018, 16, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adir, N.; Lerner, N. The Crystal Structure of a Novel Unmethylated Form of C-phycocyanin, a Possible Connector between Cores and Rods in Phycobilisomes. J. Biol. Chem. 2003, 278, 25926–25932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arteni, A.-A.; Ajlani, G.; Boekema, E.J. Structural organisation of phycobilisomes from Synechocystis sp. strain PCC6803 and their interaction with the membrane. Biochim. Biophys. Acta (BBA) Gen. Subj. 2009, 1787, 272–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Huang, Y.; Zhang, R.; Cai, T.; Cai, Y. Medical Application ofSpirulina platensisDerived C-Phycocyanin. Evid. Based Complement. Altern. Med. 2016, 2016, 7803846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, R.; Lu, R.; Zhang, Y.; Zhu, M.; Zhu, W.; Yang, R.; Zhang, E.; Ying, J.; Xu, T.; Yi, H.; et al. Spirulina phycocyanin induces differential protein expression and apoptosis in SKOV-3 cells. Int. J. Biol. Macromol. 2015, 81, 951–959. [Google Scholar] [CrossRef]
- Hao, S.; Li, S.; Wang, J.; Zhao, L.; Yan, Y.; Wu, T.; Zhang, J.; Wang, C. C-Phycocyanin Suppresses the In Vitro Proliferation and Migration of Non-Small-Cell Lung Cancer Cells through Reduction of RIPK1/NF-κB Activity. Mar. Drugs 2019, 17, 362. [Google Scholar] [CrossRef] [Green Version]
- Hao, S.; Li, S.; Wang, J.; Zhao, L.; Yan, Y.; Cao, Q.; Wu, T.; Liu, L.; Wang, C. Transcriptome Analysis of Phycocyanin-Mediated Inhibitory Functions on Non-Small Cell Lung Cancer A549 Cell Growth. Mar. Drugs 2018, 16, 511. [Google Scholar] [CrossRef] [Green Version]
- Catassi, A.; Cesario, A.; Arzani, D.; Menichini, P.; Alama, A.; Bruzzo, C.; Imperatori, A.; Rotolo, N.; Granone, P.; Russo, P. Characterization of apoptosis induced by marine natural products in non small cell lung cancer A549 cells. Cell. Mol. Life Sci. 2006, 63, 2377–2386. [Google Scholar] [CrossRef]
- Fernández-Rojas, B.; Hernández-Juárez, J.; Pedraza-Chaverri, J. Nutraceutical properties of phycocyanin. J. Funct. Foods 2014, 11, 375–392. [Google Scholar] [CrossRef]
- Wu, X.-J.; Yang, H.; Sheng, Y.; Zhu, Y.; Li, P.-P. Fluorescence Properties of a Novel Cyanobacteriochrome GAF Domain from Spirulina that Exhibits Moderate Dark Reversion. Int. J. Mol. Sci. 2018, 19, 2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subhashini, J.; Mahipal, S.V.; Reddy, M.C.; Reddy, M.M.; Rachamallu, A.; Reddanna, P. Molecular mechanisms in C-Phycocyanin induced apoptosis in human chronic myeloid leukemia cell line-K562. Biochem. Pharmacol. 2004, 68, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, N.T. Production of phycocyanin—A pigment with applications in biology, biotechnology, foods and medicine. Appl. Microbiol. Biotechnol. 2008, 80, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jung, F.; Krüger-Genge, A.; Waldeck, P.; Küpper, J.-H. Spirulina platensis, a super food? J. Cell. Biotechnol. 2019, 5, 43–54. [Google Scholar] [CrossRef]
- Madhyastha, H.; Sivashankari, S.; Vatsala, T. C-phycocyanin from Spirulina fussiformis exposed to blue light demonstrates higher efficacy of in vitro antioxidant activity. Biochem. Eng. J. 2009, 43, 221–224. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, W.; Lu, J.-J.; Huang, M.; Li, Y.; Chen, M.; Wu, G.; Gong, J.; Zhong, Z.; Xu, Z.; Dang, Y.; et al. Anti-cancer natural products isolated from chinese medicinal herbs. Chin. Med. 2011, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Chu, X.; Gao, M.; Li, W. Apoptotic mechanism of MCF-7 breast cells in vivo and in vitro induced by photodynamic therapy with C-phycocyanin. Acta Biochim. Biophys. Sin. 2009, 42, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Wong, Y.-S. In Vitro Antioxidant and Antiproliferative Activities of Selenium-Containing Phycocyanin from Selenium-Enriched Spirulina platensis. J. Agric. Food Chem. 2008, 56, 4352–4358. [Google Scholar] [CrossRef]
- Arunasree, K.M.; Roy, K.R.; Reddy, N.P.; Dheeraj, B.; Reddy, G.V.; Reddanna, P. Alteration of mitochondrial membrane potential by Spirulina platensis C-phycocyanin induces apoptosis in the doxorubicinresistant human hepatocellular-carcinoma cell line HepG2. Biotechnol. Appl. Biochem. 2007, 47, 159–167. [Google Scholar] [CrossRef]
- Bingula, R.; Dupuis, C.; Pichon, C.; Berthon, J.-Y.; Filaire, M.; Pigeon, L.; Filaire, E. Study of the Effects of Betaine and/or C-Phycocyanin on the Growth of Lung Cancer A549 Cells In Vitroand In Vivo. J. Oncol. 2016, 2016, 8162952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Gao, M.; Chu, X.-M.; Teng, L.; Lv, C.-Y.; Yang, P.; Yin, Q.-F. The synergistic antitumor effects of all-trans retinoic acid and C-phycocyanin on the lung cancer A549 cells in vitro and in vivo. Eur. J. Pharmacol. 2015, 749, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, Y.; Gao, X.; Carter, C.L.; Liu, Z.-R. The recombinant β subunit of C-phycocyanin inhibits cell proliferation and induces apoptosis. Cancer Lett. 2007, 247, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Gardeva, E.; Toshkova, R.; Yossifova, L.; Minkova, K.; Ivanova, N.; Gigova, L. Antitumor activity of C-phycocyanin from Arthronema africanum (Cyanophyceae). Braz. Arch. Biol. Technol. 2014, 57, 675–684. [Google Scholar] [CrossRef] [Green Version]
- Liao, G.; Gao, B.; Gao, Y.; Yang, X.; Cheng, X.; Ou, Y. Phycocyanin Inhibits Tumorigenic Potential of Pancreatic Cancer Cells: Role of Apoptosis and Autophagy. Sci. Rep. 2016, 6, srep34564. [Google Scholar] [CrossRef]
- Remirez, D.; Ledón, N.; González, R. Role of histamine in the inhibitory effects of phycocyanin in experimental models of allergic inflammatory response. Mediat. Inflamm. 2002, 11, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Wang, Y.; Zhu, F.; Liu, G.; Liu, H.; Ji, H.; Zheng, S.; Li, B. Molecular Mechanism of Anti-Cancer Activity of the Nano-Drug C-PC/CMC-CD59sp NPs in Cervical Cancer. J. Cancer 2019, 10, 92–104. [Google Scholar] [CrossRef]
- Gao, M.-H.; Li, B.; Zhang, X.-C.; Chu, X.-M. Molecular immune mechanism of C-phycocyanin from Spirulina platensis induces apoptosis in HeLa cells in vitro. Biotechnol. Appl. Biochem. 2006, 43, 155–164. [Google Scholar] [CrossRef]
- Abu Zaid, A.A.; Hammad, D.M.; Sharaf, E.M. Antioxidant and Anticancer Activity of Spirulina platensis Water Extracts. Int. J. Pharmacol. 2015, 11, 846–851. [Google Scholar] [CrossRef]
- Koníčková, R.; Vaňková, K.; Vaníková, J.; Váňová, K.; Muchová, L.; Subhanová, I.; Zadinová, M.; Zelenka, J.; Dvořák, A.; Kolář, M.; et al. Anti-Cancer Effects of Blue-Green Alga Spirulina Platensis, a Natural Source of Bilirubin-like Tetrapyrrolic Compounds. Ann. Hepatol. 2014, 13, 273–283. [Google Scholar] [CrossRef]
- Basha, O.M.; A Hafez, R.; El-Ayouty, Y.M.; Mahrous, K.F.; Bareedy, M.H.; Salama, A.M. C-Phycocyanin inhibits cell proliferation and may induce apoptosis in human HepG2 cells. Egypt. J. Immunol. 2008, 15, 161–167. [Google Scholar] [PubMed]
- Czerwonka, A.; Kaławaj, K.; Sławińska-Brych, A.; Lemieszek, M.K.; Bartnik, M.; Wojtanowski, K.K.; Zdzisińska, B.; Rzeski, W. Anticancer effect of the water extract of a commercial Spirulina (Arthrospira platensis) product on the human lung cancer A549 cell line. Biomed. Pharmacother. 2018, 106, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Thangam, R.; Suresh, V.; Princy, W.A.; Rajkumar, M.; Senthilkumar, N.; Gunasekaran, P.; Rengasamy, R.; Anbazhagan, C.; Krishnasamy, K.; Kannan, S. C-Phycocyanin from Oscillatoria tenuis exhibited an antioxidant and in vitro antiproliferative activity through induction of apoptosis and G0/G1 cell cycle arrest. Food Chem. 2013, 140, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Tiwari, R.; Srivastava, V.; Singh, T.B.; Asthana, R.K. Fresh Water Cyanobacteria Geitlerinema sp. CCC728 and Arthrospira sp. CCC729 as an Anticancer Drug Resource. PLoS ONE 2015, 10, e0136838. [Google Scholar] [CrossRef] [Green Version]
- Ravi, M.; Tentu, S.; Baskar, G.; Prasad, S.R.; Raghavan, S.; Jayaprakash, P.; Jeyakanthan, J.; Rayala, S.K.; Venkatraman, G. Molecular mechanism of anti-cancer activity of phycocyanin in triple-negative breast cancer cells. BMC Cancer 2015, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Safaei, M.; Maleki, H.; Soleimanpour, H.; Norouzy, A.; Zahiri, H.S.; Vali, H.; Noghabi, K.A. Development of a novel method for the purification of C-phycocyanin pigment from a local cyanobacterial strain Limnothrix sp. NS01 and evaluation of its anticancer properties. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Pardhasaradhi, B.V.V.; Ali, A.M.; Kumari, A.L.; Reddanna, P.; Khar, A. Phycocyanin-mediated apoptosis in AK-5 tumor cells involves down-regulation of Bcl-2 and generation of ROS. Mol. Cancer Ther. 2003, 2, 1165–1170. [Google Scholar]
- Wang, C.-Y.; Wang, X.; Wang, Y.; Zhou, T.; Bai, Y.; Li, Y.-C.; Huang, B. Photosensitization of phycocyanin extracted from Microcystis in human hepatocellular carcinoma cells: Implication of mitochondria-dependent apoptosis. J. Photochem. Photobiol. B Biol. 2012, 117, 70–79. [Google Scholar] [CrossRef]
- Beneke, R.; Geisen, C.; Zevnik, B.; Bauch, T.; Müller, W.-U.; Küpper, J.-H.; Möröy, T. DNA Excision Repair and DNA Damage-Induced Apoptosis Are Linked to Poly(ADP-Ribosyl)ation but Have Different Requirements for p53. Mol. Cell. Biol. 2000, 20, 6695–6703. [Google Scholar] [CrossRef] [Green Version]
- Oftedal, L.; Selheim, F.; Wahlsten, M.; Sivonen, K.; Døskeland, S.O.; Herfindal, L. Marine Benthic Cyanobacteria Contain Apoptosis-Inducing Activity Synergizing with Daunorubicin to Kill Leukemia Cells, but not Cardiomyocytes. Mar. Drugs 2010, 8, 2659–2672. [Google Scholar] [CrossRef]
- Davies, G.; Martin, L.-A.; Sacks, N.; Dowsett, M. Cyclooxygenase-2 (COX-2), aromatase and breast cancer:a possible role for COX-2 inhibitors in breast cancer chemoprevention. Ann. Oncol. 2002, 13, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Telliez, A.; Furman, C.; Pommery, N.; Hénichart, J.-P. Mechanisms Leading to COX-2 Expression and COX-2 Induced Tumorigenesis: Topical Therapeutic Strategies Targeting COX-2 Expression and Activity. Anti-Cancer Agents Med. Chem. 2006, 6, 187–208. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-C.; Wu, C.-H.; Tu, Y.-K.; Huang, S.-Y.; Chou, P.-C. Exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin increases the activation of aryl hydrocarbon receptor and is associated with the aggressiveness of osteosarcoma MG-63 osteoblast-like cells. Oncol. Lett. 2018, 16, 3849–3857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Rahden, B.H.; Stein, H.J.; Pühringer, F.; Koch, I.; Langer, R.; Piontek, G.; Siewert, J.R.; Höfler, H.; Sarbia, M. Coexpression of Cyclooxygenases (COX-1, COX-2) and Vascular Endothelial Growth Factors (VEGF-A, VEGF-C) in Esophageal Adenocarcinoma. Cancer Res. 2005, 65, 5038–5044. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, R.; Kawada, K.; Sakai, Y. Prostaglandin E2/EP Signaling in the Tumor Microenvironment of Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 6254. [Google Scholar] [CrossRef] [Green Version]
- Noda, M.; Tatsumi, Y.; Tomizawa, M.; Takama, T.; Mitsufuji, S.; Sugihara, H.; Kashima, K.; Hattori, T. Effects of etodolac, a selective cyclooxygenase-2 inhibitor, on the expression of E-cadherin-catenin complexes in gastrointestinal cell lines. J. Gastroenterol. 2002, 37, 896–904. [Google Scholar] [CrossRef]
- Saini, M.K.; Sanyal, S.N. Targeting angiogenic pathway for chemoprevention of experimental colon cancer using C-phycocyanin as cyclooxygenase-2 inhibitor. Biochem. Cell Biol. 2014, 92, 206–218. [Google Scholar] [CrossRef]
- Krüger-Genge, A.; Blocki, A.; Franke, R.-P.; Jung, F. Vascular Endothelial Cell Biology: An Update. Int. J. Mol. Sci. 2019, 20, 4411. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Wang, Y.; Yin, Q.; Liu, G.; Liu, H.; Huang, Y.; Liangqian, J. Phycocyanin: A Potential Drug for Cancer Treatment. J. Cancer 2017, 8, 3416–3429. [Google Scholar] [CrossRef] [Green Version]
- Ying, J.; Ruowang, P.; Ji, H.; Lin, C.; Pan, R.; Zhou, L.; Song, Y.; Zhang, E.; Ren, P.; Chen, J.; et al. Transcriptome analysis of phycocyanin inhibitory effects on SKOV-3 cell proliferation. Gene 2016, 585, 58–64. [Google Scholar] [CrossRef]
- Nowell, P.C. The clonal evolution of tumor cell populations. Science 1976, 194, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Loeb, L.A.; Springgate, C.F.; Battula, N. Errors in DNA replication as a basis of malignant changes. Cancer Res. 1974, 34, 2311–2321. [Google Scholar]
- Mroz, E.A.; Rocco, J.W.; Rocco, J. The challenges of tumor genetic diversity. Cancer 2016, 123, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Mroz, E.A.; Tward, A.M.; Hammon, R.J.; Ren, Y.; Rocco, J. Intra-tumor Genetic Heterogeneity and Mortality in Head and Neck Cancer: Analysis of Data from The Cancer Genome Atlas. PLoS Med. 2015, 12, e1001786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mroz, E.A.; Tward, A.D.; Pickering, C.R.; Myers, J.N.; Ferris, R.L.; Rocco, J.W. High intratumor genetic heterogeneity is related to worse outcome in patients with head and neck squamous cell carcinoma. Cancer 2013, 119, 3034–3042. [Google Scholar] [CrossRef] [PubMed]
- Pereira, B.; Chin, S.-F.; Rueda, O.M.; Vollan, H.-K.M.; Provenzano, E.; Bardwell, H.A.; Pugh, M.; Jones, L.A.; Russell, R.; Sammut, S.-J.; et al. The somatic mutation profiles of 2433 breast cancers refine their genomic and transcriptomic landscapes. Nat. Commun. 2016, 7, 11479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Li, B.; Yin, Q.-F.; Wang, Y.-J. Carboxymethyl chitosan nanoparticles coupled with CD59-specific ligand peptide for targeted delivery of C-phycocyanin to HeLa cells. Tumor Biol. 2017, 39. [Google Scholar] [CrossRef] [Green Version]
- Rangasamy, J.; Prabaharan, M.; Nair, S.; Tokura, S.; Tamura, H.; Selvamurugan, N. Novel carboxymethyl derivatives of chitin and chitosan materials and their biomedical applications. Prog. Mater. Sci. 2010, 55, 675–709. [Google Scholar] [CrossRef]
- Watson, N.F.S.; Durrant, L.G.; Madjd, Z.; Ellis, I.O.; Scholefield, J.H.; Spendlove, I. Expression of the membrane complement regulatory protein CD59 (protectin) is associated with reduced survival in colorectal cancer patients. Cancer Immunol. Immunother. 2005, 55, 973–980. [Google Scholar] [CrossRef]
- Varsano, S.; Rashkovsky, L.; Shapiro, H.; Ophir, D.; Mark-Bentankur, T. Human lung cancer cell lines express cell membrane complement inhibitory proteins and are extremely resistant to complement-mediated lysis; a comparison with normal human respiratory epithelium in vitro, and an insight into mechanism(s) of resistance. Clin. Exp. Immunol. 1998, 113, 173–182. [Google Scholar] [CrossRef]
- Crnogorac-Jurcevic, T.; Efthimiou, E.; Nielsen, T.; Loader, J.; Terris, B.; Stamp, G.; Baron, A.; Scarpa, A.; Lemoine, N.R. Expression profiling of microdissected pancreatic adenocarcinomas. Oncogene 2002, 21, 4587–4594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezzanzanica, D.; Balladore, E.; Turatti, F.; Luison, E.; Alberti, P.; Bagnoli, M.; Figini, M.; Mazzoni, A.; Raspagliesi, F.; Oggionni, M.; et al. CD95-Mediated Apoptosis Is Impaired at Receptor Level by Cellular FLICE-Inhibitory Protein (Long Form) in Wild-Type p53 Human Ovarian Carcinoma. Clin. Cancer Res. 2004, 10, 5202–5214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Huang, B.; Zuo, M.-M.; Guo, R.-Y.; Wei, H. Preparation of the phycoerythrin subunit liposome in a photodynamic experiment on liver cancer cells. Acta Pharmacol. Sin. 2008, 29, 1539–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.; Jiang, B.; Liang, J.; Wen, C.; Shen, X.-C. Phycocyanin functionalized single-walled carbon nanohorns hybrid for near-infrared light-mediated cancer phototheranostics. Carbon 2019, 143, 814–827. [Google Scholar] [CrossRef]
- Du, S.-W.; Zhang, L.-K.; Han, K.; Chen, S.; Hu, Z.; Guan, Y.; Hu, K.; Yin, L.; Wu, B.; Guan, Y. Combined Phycocyanin and Hematoporphyrin Monomethyl Ether for Breast Cancer Treatment via Photosensitizers Modified Fe3O4 Nanoparticles Inhibiting the Proliferation and Migration of MCF-7 Cells. Biomacromolecules 2017, 19, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Fu, X.; Huang, W.; Li, C.; Wang, X.; Huang, B. Photodynamic effect and mechanism study of selenium-enriched phycocyanin from Spirulina platensis against liver tumours. J. Photochem. Photobiol. B Biol. 2018, 180, 89–97. [Google Scholar] [CrossRef]
- Li, B.; Gao, M.; Lv, C.-Y.; Yang, P.; Yin, Q.-F. Study of the synergistic effects of all-transretinoic acid and C-phycocyanin on the growth and apoptosis of A549 cells. Eur. J. Cancer Prev. 2016, 25, 97–101. [Google Scholar] [CrossRef]
- Saini, M.K.; Vaiphei, K.; Sanyal, S.N. Chemoprevention of DMH-induced rat colon carcinoma initiation by combination administration of piroxicam and C-phycocyanin. Mol. Cell. Biochem. 2011, 361, 217–228. [Google Scholar] [CrossRef]
- Saini, M.K.; Sanyal, S.N. Piroxicam and c-phycocyanin prevent colon carcinogenesis by inhibition of membrane fluidity and canonical Wnt/β-catenin signaling while up-regulating ligand dependent transcription factor PPARγ. Biomed. Pharmacother. 2014, 68, 537–550. [Google Scholar] [CrossRef]
- Hashem, M.A.; Shoeeb, S.B.; Abd-Elhakim, Y.M.; Mohamed, W.A. The antitumor activity of Arthrospira platensis and/or cisplatin in a murine model of Ehrlich ascites carcinoma with hematinic and hepato-renal protective action. J. Funct. Foods 2020, 66, 103831. [Google Scholar] [CrossRef]
- Zhang, X.; Fan, T.; Li, S.; Guan, F.; Zhang, J.; Liu, H. C-Phycocyanin elicited antitumor efficacy via cell-cycle arrest, apoptosis induction, and invasion inhibition in esophageal squamous cell carcinoma. J. Recept. Signal Transduct. 2019, 39, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, L.; Yin, Q.; Liu, H.; Liu, G.; Zhu, G.; Li, B. The Targeted Antitumor Effects of C- PC/CMC-CD59sp Nanoparticles on HeLa Cells in Vitro and in Vivo. J. Cancer 2017, 8, 3001–3013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Xu, X.; Jiang, L.; Ji, H.; Zhu, F.; Jin, B.; Han, J.; Dong, X.; Yang, F.; Li, B. Targeted Antitumor Mechanism of C-PC/CMC-CD55sp Nanospheres in HeLa Cervical Cancer Cells. Front. Pharmacol. 2020, 11, 906. [Google Scholar] [CrossRef] [PubMed]
- Gantar, M.; Dhandayuthapani, S.; Rathinavelu, A. Phycocyanin Induces Apoptosis and Enhances the Effect of Topotecan on Prostate Cell Line LNCaP. J. Med. Food 2012, 15, 1091–1095. [Google Scholar] [CrossRef]
- Ji, H.; Liu, G.; Han, J.; Zhu, F.; Dong, X.; Li, B. C-phycocyanin inhibits epithelial-to-mesenchymal transition in Caski cells. Cancer Cell Int. 2020, 20, 1–11. [Google Scholar] [CrossRef]
- Romay, C.; Ledón, N.; González, R. Further studies on anti-inflammatory activity of phycocyanin in some animal models of inflammation. Inflamm. Res. 1998, 47, 334–338. [Google Scholar] [CrossRef]
- Naidu, K.A.; Sarada, R.; Manoj, G.; Khan, M.; Swamy, M.M.; Viswanatha, S.; Murthy, K.N.; Ravishankar, G.; Srinivas, L. Toxicity Assessment Of Phycocyanin—A Blue Colorant From Blue Green Alga Spirulina platensis. Food Biotechnol. 1999, 13, 51–66. [Google Scholar] [CrossRef]
- Ou, Y.; Lin, L.; Pan, Q.; Yang, X.; Cheng, X. Preventive effect of phycocyanin from Spirulina platensis on alloxan-injured mice. Environ. Toxicol. Pharmacol. 2012, 34, 721–726. [Google Scholar] [CrossRef]
- Gupta, M.; Dwivedi, U.N.; Khandelwal, S. C-Phycocyanin: An effective protective agent against thymic atrophy by tributyltin. Toxicol. Lett. 2011, 204, 2–11. [Google Scholar] [CrossRef]
- Vadiraja, B.; Gaikwad, N.; Madyastha, K. Hepatoprotective Effect of C-Phycocyanin: Protection for Carbon Tetrachloride andR-(+)-Pulegone-Mediated Hepatotoxicty in Rats. Biochem. Biophys. Res. Commun. 1998, 249, 428–431. [Google Scholar] [CrossRef] [Green Version]
- Krüger-Genge, A.; Steinbrecht, S.; Jung, C.G.H.; Waldeck, P.; Küpper, J.-H.; Storsberg, J.; Jung, F. Effect of an Aqueous Extract of Spirulina Platensis on the Endothelialization Potential of Human Venous Endothelial Cells (HUVEC). Clin. Hemorheol. Microcirc. 2020, accepted. [Google Scholar]
- Toss, A.; Cortesi, L. Molecular Mechanisms of PARP Inhibitors in BRCA-related Ovarian Cancer. J. Cancer Sci. Ther. 2013, 5, 409–416. [Google Scholar] [CrossRef]
- Morales, J.C.; Li, L.; Fattah, F.J.; Dong, Y.; Bey, E.A.; Patel, M.; Gao, J.; Boothman, D.A. Review of Poly (ADP-ribose) Polymerase (PARP) Mechanisms of Action and Rationale for Targeting in Cancer and Other Diseases. Crit. Rev. Eukaryot. Gene Expr. 2014, 24, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Reddy, M.C.; Subhashini, J.; Mahipal, S.; Bhat, V.B.; Reddy, P.S.; Kiranmai, G.; Madyastha, K.; Reddanna, P. C-Phycocyanin, a selective cyclooxygenase-2 inhibitor, induces apoptosis in lipopolysaccharide-stimulated RAW 264.7 macrophages. Biochem. Biophys. Res. Commun. 2003, 304, 385–392. [Google Scholar] [CrossRef]
- Chelariu-Raicu, A.; Coleman, R.L. Breast cancer (BRCA) gene testing in ovarian cancer. Chin. Clin. Oncol. 2020, 9, 63. [Google Scholar] [CrossRef]
- Wu, C.-E.; Yeh, D.-W.; Pan, Y.-R.; Huang, W.-K.; Chen, M.-H.; Chang, J.W.-C.; Chen, J.-S.; Wang, Y.-C.; Chun-Nan, Y. Chromosomal Instability May Not Be a Predictor for Immune Checkpoint Inhibitors from a Comprehensive Bioinformatics Analysis. Life 2020, 10, 276. [Google Scholar] [CrossRef]
- Agata, S.; Tognazzo, S.; Alducci, E.; Matricardi, L.; Moserle, L.; Barana, D.; Montagna, M. Segregation analysis of the BRCA2 c.9227G>T variant in multiple families suggests a pathogenic role in breast and ovarian cancer predisposition. Sci. Rep. 2020, 10, 1–6. [Google Scholar] [CrossRef]
- Verdaguer, H.; Acosta, D.; Macarulla, T. A new targeted treatment for patients with a germline BRCA mutation: Olaparib in pancreatic cancer. Futur. Oncol. 2020, 16, 2691–2700. [Google Scholar] [CrossRef]
- Abida, W.; Patnaik, A.; Campbell, D.; Shapiro, J.; Bryce, A.H.; McDermott, R.; Sautois, B.; Vogelzang, N.J.; Bambury, R.M.; Voog, E.; et al. Rucaparib in Men With Metastatic Castration-Resistant Prostate Cancer Harboring a BRCA1 or BRCA2 Gene Alteration. J. Clin. Oncol. 2020, 38, 3763–3772. [Google Scholar] [CrossRef]
- Deng, C.-X. BRCA1: Cell cycle checkpoint, genetic instability, DNA damage response and cancer evolution. Nucleic Acids Res. 2006, 34, 1416–1426. [Google Scholar] [CrossRef] [Green Version]
- Risdon, E.N.; Chau, C.H.; Price, D.K.; Sartor, O.; Figg, W.D. PARP Inhibitors and Prostate Cancer: To Infinity and Beyond BRCA. Oncologist 2021, 26, e115–e129. [Google Scholar] [CrossRef] [PubMed]
- Westerink, W.M.; Stevenson, J.C.; Horbach, G.J.; Schoonen, W.G. The development of RAD51C, Cystatin A, p53 and Nrf2 luciferase-reporter assays in metabolically competent HepG2 cells for the assessment of mechanism-based genotoxicity and of oxidative stress in the early research phase of drug development. Mutat. Res. Toxicol. Environ. Mutagen. 2010, 696, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Malik, Z.; Singh, S.S.; Chirom, K.; Ishrat, R.; Malik, Z. Exploring novel key regulators in breast cancer network. PLoS ONE 2018, 13, e0198525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chornokur, G.; Lin, H.-Y.; Tyrer, J.P.; Lawrenson, K.; Dennis, J.; Amankwah, E.K.; Qu, X.; Tsai, Y.-Y.; Jim, H.S.L.; Chen, Z.; et al. Common Genetic Variation in Cellular Transport Genes and Epithelial Ovarian Cancer (EOC) Risk. PLoS ONE 2015, 10, e0128106. [Google Scholar] [CrossRef] [Green Version]



| Tumor Type | Cell Line | Phycocyanin Concentration | Application Time (h) | Proliferation (%) | IC50 | Morphology | Ref. | ||
|---|---|---|---|---|---|---|---|---|---|
| Cervical carcinoma | HeLa | 80; 200 | µg/mL | 72; 24 | −32; −20 | -; 1104 | µg/mL | epithelial | [57,58] |
| Human colorectal adenocarcinoma | HCT116 | 50 | µg/mL | 48 | −73 | 18.8 | µg/mL | epithelial | [59] |
| Pancreatic adenocarcinoma | Capan-1 | 100 | µM | 72 | −80 | 6.2 | µM | epithelial | [55] |
| Pancreatic adenocarcinoma Pancreatic adenocarcinoma | BxPC3 PA-TU-8902 | 100 0.3 | µM g/L | 72 24 | −100 −82 | 15.1 - | µM - | epithelial epithelial | [55,60] |
| Human ductal pancreas carcinoma | PANC-1 | 100 | µM | 72 | −70 | 12.2 | µM | epithelial | [55] |
| Hepatoblastoma | HepG2 | 100 | µM | 72 | −76 | 13 | µM | epithelial | [55] |
| Hepatoblastoma | HepG2 | 7; 50 | µg/mL | 24; 48 | −61; −75 | 1.75; 22.3 | µg/mL | epithelial | [59,61] |
| Prostate carcinoma | DU145 | 100 | µM | 72 | −70 | 18 | µM | epithelial | [55] |
| Large cell lung cancer Lung adenocarcinoma | H460 A549 | 100 50 | µM µg/mL | 72 24 | −95 −38 | 14 99.2 | µM µg/mL | epithelial epithelial | [55,62] |
| Alveolar adenocarcinoma | A549 | 60 | µg/mL | 48 | −64 | - | - | epithelial | [63] |
| Nsc broncho carcinoma | H1299 | 4.8 | µM | 24 | −11.3 | - | - | epithelial | [33] |
| Nsc broncho carcinoma | H460 | 4.8 | µM | 24 | −3.7 | - | - | epithelial | [33] |
| Nsc broncho carcinoma | LTEP-A2 | 4.8 | µM | 4 | −14.5 | - | - | epithelial | [33] |
| Human colorectal adenocarcinoma | HT-29 | 50; 200 | µg/mL | 48; 72 | −63; −100 | - | - | epithelial | [63,64] |
| Triple negative breast cancer | MDA-MB-231 | 20 | µM | 6 | −82 | 5.98 | µM | epithelial | [65] |
| Triple negative breast cancer | MDA-MB-231 | 294 | µg/mL | 24 | −30 | 294 | µg/mL | epithelial | [57] |
| Ductal carcinoma | BT-474 | 20 | µM | 6 | −80 | 8.45 | µM | epithelial | [65] |
| Breast cancer | MCF-7 | 20; 100 | µM | 6; 72 | −58; −65 | 15.4; 33 | µM | epithelial | [4,65] |
| Breast cancer | MCF-7 | 5.66 | µg/mL | 48 | - | 5.66 | µg/mL | epithelial | [66] |
| Breast cancer | MCF-7 | 40 | µM | 72 | −53 | - | - | epithelial | [49] |
| Mammary gland adenocarcinoma | SKBR 3 | 20 | µM | 6 | −60 | 17.7 | µM | epithelial | [65] |
| Squamous cell carcinoma, cervix | SiHa | 376 | µg/mL | 24 | −35 | 376 | µg/mL | epithelial | [57] |
| Malignant melanoma | A375 | 40 | µM | 72 | −54 | - | - | epithelial | [49] |
| Chronic myelogenous leukemia Histiocytic tumor | K562 AK-5 | 100 60 | µM µM | 96 72 | −49 −70 | - 60 | - µM | lymphoblast macrophage | [42,67] |
| Cell Origin | Cell Type | Phycocyanin Concentration | Appl. Time (h) | Prolif. (%) | IC50 | Morphology | Ref. | ||
|---|---|---|---|---|---|---|---|---|---|
| Ovary (Chin. Hamster) | CHO | 20 | µM | 6 | ≈ | - | - | epithelial | [58] |
| Mammary gland | MCF-10A | 25 | µg/mL | 24 | ↑ | >20 | - | epithelial | [65] |
| Skin | HSF | 1000 | µg/mL | 24 | ≈ | - | - | fibroblast | [62] |
| Skin | CCD-986sk | 40 | µM | 72 | +42 | - | - | fibroblast | [35] |
| Skin | Hs68 | 100 | µM | 72 | ≈ | - | - | fibroblast | [49] |
| Liver | LO2 | 100 | µM | 72 | ≈ | - | - | [55] | |
| Liver | QSG-7701 | 100 | µM | 72 | ≈ | - | - | epithelial | [55] |
| Human heart ventricle | AC-16 | 100 | µM | 72 | ≈ | - | - | [55] | |
| Kidney cortex | HK-2 | 100 | µM | 72 | ≈ | - | - | epithelial | [55] |
| Peripheral blood | NK-82 | 100 | µM | 72 | ≈ | - | - | NK cells | [55] |
| Endothelium of umbilical vein | HUVEC | 100 | µM | 72 | ≈ | - | - | epithelial | [55] |
| Endothelium of umbilical vein | HUVEC | 50 | µg/mL | 80 | +31 | - | - | epithelial | [111] |
| Connective tissue (Mouse) | L929 | 596 | µg/mL | 24 | −25 | 596 | µg/mL | fibroblast | [57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braune, S.; Krüger-Genge, A.; Kammerer, S.; Jung, F.; Küpper, J.-H. Phycocyanin from Arthrospira platensis as Potential Anti-Cancer Drug: Review of In Vitro and In Vivo Studies. Life 2021, 11, 91. https://doi.org/10.3390/life11020091
Braune S, Krüger-Genge A, Kammerer S, Jung F, Küpper J-H. Phycocyanin from Arthrospira platensis as Potential Anti-Cancer Drug: Review of In Vitro and In Vivo Studies. Life. 2021; 11(2):91. https://doi.org/10.3390/life11020091
Chicago/Turabian StyleBraune, Steffen, Anne Krüger-Genge, Sarah Kammerer, Friedrich Jung, and Jan-Heiner Küpper. 2021. "Phycocyanin from Arthrospira platensis as Potential Anti-Cancer Drug: Review of In Vitro and In Vivo Studies" Life 11, no. 2: 91. https://doi.org/10.3390/life11020091