Trace Element Concentrations Associated with Mid-Paleozoic Microfossils as Biosignatures to Aid in the Search for Life †

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Farley, K.A.; Williford, K.H. Seeking Signs of Life, and More: NASA’s Mars 2020 Mission. Eos 2017, 98. [Google Scholar] [CrossRef]
- Mustard, J.F.; Adler, M.; Allwood, A.; Bass, D.S.; Beaty, D.W.; Bell, J.F.; Brinckerhoff, W.; Carr, M.; Des Marais, D.J.; Brake, B. Report of the Mars 2020 Science Definition Team. Mars Explor. Program Anal. Group (Mepag) 2013, 150, 155–205. [Google Scholar]
- Muscente, A.D.; Czaja, A.D.; Tuggle, J.; Winkler, C.; Xiao, S. Manganese Oxides Resembling Microbial Fabrics and Their Implications for Recognizing Inorganically Preserved Microfossils. Astrobiology 2018, 18, 249–258. [Google Scholar] [CrossRef]
- Wacey, D.; Noffke, N.; Saunders, M.; Guagliardo, P.; Pyle, D.M. Volcanogenic Pseudo-Fossils from The ∼3.48 Ga Dresser Formation, Pilbara, Western Australia. Astrobiology 2018, 18, 539–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schopf, J.W.; Kudryavtsev, A.B.; Agresti, D.G.; Wdowiak, T.J.; Czaja, A.D. Laser–Raman Imagery of Earth’s Earliest Fossils. Nature 2002, 416, 73. [Google Scholar] [CrossRef]
- Brasier, M.D.; Green, O.R.; Jephcoat, A.P.; Kleppe, A.K.; Van Kranendonk, M.J.; Lindsay, J.F.; Steele, A.; Grassineau, N.V. Questioning the Evidence for Earth’s Oldest Fossils. Nature 2002, 416, 76. [Google Scholar] [CrossRef] [PubMed]
- Javaux, E.J. Challenges in Evidencing the Earliest Traces of Life. Nature 2019, 572, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Cady, S.L.; Farmer, J.D.; Grotzinger, J.P.; Schopf, J.W.; Steele, A. Morphological Biosignatures and the Search for Life on Mars. Astrobiology 2003, 3, 351–368. [Google Scholar] [CrossRef]
- Schopf, J.W. Microfossils of the Early Archean Apex Chert: New Evidence of the Antiquity of Life. Science 1993, 260, 640–646. [Google Scholar] [CrossRef] [Green Version]
- Westall, F.; Hickman-Lewis, K.; Cavalazzi, B. Biosignatures in deep time. In Biosignatures for Astrobiology; Springer: Cham, Switzerland, 2019; pp. 145–164. [Google Scholar]
- Hays, L.E.; Graham, H.V.; Des Marais, D.J.; Hausrath, E.M.; Horgan, B.; McCollom, T.M.; Parenteau, M.N.; Potter-McIntyre, S.L.; Williams, A.J.; Lynch, K.L. Biosignature Preservation and Detection in Mars Analog Environments. Astrobiology 2017, 17, 363–400. [Google Scholar] [CrossRef]
- Tarnas, J.D.; Mustard, J.F.; Lin, H.; Goudge, T.A.; Amador, E.S.; Bramble, M.S.; Kremer, C.H.; Zhang, X.; Itoh, Y.; Parente, M. Orbital Identification of Hydrated Silica in Jezero Crater, Mars. Geophys. Res. Lett. 2019, 46, 12771–12782. [Google Scholar] [CrossRef] [Green Version]
- Goudge, T.A.; Mustard, J.F.; Head, J.W.; Fassett, C.I.; Wiseman, S.M. Assessing the Mineralogy of the Watershed and Fan Deposits of the Jezero Crater Paleolake System, Mars. J. Geophys. Res. Planets 2015, 120, 775–808. [Google Scholar] [CrossRef]
- Osinski, G.R.; Tornabene, L.L.; Banerjee, N.R.; Cockell, C.S.; Flemming, R.; Izawa, M.R.; McCutcheon, J.; Parnell, J.; Preston, L.J.; Pickersgill, A.E. Impact-Generated Hydrothermal Systems on Earth and Mars. Icarus 2013, 224, 347–363. [Google Scholar] [CrossRef] [Green Version]
- Ruff, S.W.; Farmer, J.D. Silica Deposits on Mars with Features Resembling Hot Spring Biosignatures at El Tatio in Chile. Nat. Commun. 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Walter, M.R.; Des Marais, D.J. Preservation of Biological Information in Thermal Spring Deposits: Developing a Strategy for the Search for Fossil Life on Mars. Icarus 1993, 101, 129–143. [Google Scholar] [CrossRef]
- Cady, S.L.; Farmer, J.D. Fossilization Processes in Siliceous Thermal Springs: Trends in Preservation along Thermal Gradients. Ciba Found Symp. 1996, 202, 150–170. [Google Scholar]
- Lynne, B.Y.; Campbell, K.A.; James, B.J.; Browne, P.R.; Moore, J. Tracking Crystallinity in Siliceous Hot-Spring Deposits. Am. J. Sci. 2007, 307, 612–641. [Google Scholar] [CrossRef]
- Schopf, J.W. Evidences of Archean Life. In Chemical Evolution of the Early Precambrian; Elsevier: Cambridge, MA, USA, 1977; pp. 101–105. [Google Scholar]
- Djokic, T.; Van Kranendonk, M.J.; Campbell, K.A.; Walter, M.R.; Ward, C.R. Earliest Signs of Life on Land Preserved in ca. 3.5 Ga Hot Spring Deposits. Nat. Commun. 2017, 8, 15263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damer, B.; Deamer, D. The Hot Spring Hypothesis for an Origin of Life. Astrobiology 2019, 20, 429–452. [Google Scholar] [CrossRef] [Green Version]
- Olgers, F. Geology of the Drummond Basin, Queensland; Australian Government Publishing Service: Hawthorn, Australia, 1972; ISBN 0-642-00112-X.
- Walter, M.R.; McLoughlin, S.; Drinnan, A.N.; Farmer, J.D. Palaeontology of Devonian Thermal Spring Deposits, Drummond Basin, Australia. Alcheringa 1998, 22, 285–314. [Google Scholar] [CrossRef]
- Walter, M.R.; Desmarais, D.; Farmer, J.D.; Hinman, N.W. Lithofacies and Biofacies of Mid-Paleozoic Thermal Spring Deposits in the Drummond Basin, Queensland, Australia. Palaios 1996, 11, 497–518. [Google Scholar] [CrossRef] [Green Version]
- Cunneen, R.; Sillitoe, R.H. Paleozoic Hot Spring Sinter in the Drummond Basin, Queensland, Australia. Econ. Geol. 1989, 84, 135–142. [Google Scholar] [CrossRef]
- Gong, J.; Myers, K.D.; Munoz-Saez, C.; Homann, M.; Rouillard, J.; Wirth, R.; Schreiber, A.; van Zuilen, M.A. Formation and Preservation of Microbial Palisade Fabric in Silica Deposits from El Tatio, Chile. Astrobiology 2020, 20, 500–524. [Google Scholar] [CrossRef]
- Ball, J.W.; McMleskey, R.B.; Nordstrom, D.K. Water-Chemistry Data for Selected Springs, Geysers, and Streams in Yellowstone National Park, Wyoming, 2006-2008; US Geological Survey: Reston, VA, USA, 2010. [CrossRef]
- Gangidine, A.; Havig, J.R.; Fike, D.A.; Jones, C.; Hamilton, T.L.; Czaja, A.D. Trace Element Concentrations in Hydrothermal Silica Deposits as a Potential Biosignature. Astrobiology 2020, 20, 525–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havig, J.R. Geochemistry of Hydrothermal Biofilms: Composition of Biofilms in a Siliceous Sinter-Deposition Hot Spring; ProQuest: Ann Arbor, MI, USA, 2009; ISBN 1-109-20136-2. [Google Scholar]
- Martin, R.; Rodgers, K.A.; Browne, P.R.L. Aspects of the Distribution and Movement of Aluminium in the Surface of the Te Kopia Geothermal Field, Taupo Volcanic Zone, New Zealand. Appl. Geochem. 2000, 15, 1121–1136. [Google Scholar] [CrossRef]
- Banfield, J.F.; Moreau, J.W.; Chan, C.S.; Welch, S.A.; Little, B. Mineralogical Biosignatures and the Search for Life on Mars. Astrobiology 2001, 1, 447–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, G.E.; Kamber, B.S. Trace element geochemistry as a tool for interpreting microbialites. In Earliest Life on Earth: Habitats, Environments and Methods of Detection; Springer: Berlin/Heidelberg, Germany, 2011; pp. 127–170. [Google Scholar]
- Heim, C.; Simon, K.; Ionescu, D.; Reimer, A.; De Beer, D.; Quéric, N.-V.; Reitner, J.; Thiel, V. Assessing the Utility of Trace and Rare Earth Elements as Biosignatures in Microbial Iron Oxyhydroxides. Front. Earth Sci. 2015, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Simões, M.F.; Ottoni, C.A.; Antunes, A. Biogenic Metal Nanoparticles: A New Approach to Detect Life on Mars? Life 2020, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Hickman-Lewis, K.; Cavalazzi, B.; Sorieul, S.; Gautret, P.; Foucher, F.; Whitehouse, M.J.; Jeon, H.; Georgelin, T.; Cockell, C.S.; Westall, F. Metallomics in Deep Time and the Influence of Ocean Chemistry on the Metabolic Landscapes of Earth’s Earliest Ecosystems. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Allwood, A.; Clark, B.; Flannery, D.; Hurowitz, J.; Wade, L.; Elam, T.; Foote, M.; Knowles, E. Texture-Specific Elemental Analysis of Rocks and Soils with PIXL: The Planetary Instrument for X-Ray Lithochemistry on Mars 2020. In Proceedings of the 2015 IEEE Aerospace Conference, Big Sky, MT, USA, 7–14 March 2015; pp. 1–13. [Google Scholar]
- Sforna, M.-C.; Daye, M.; Philippot, P.; Somogyi, A.; Van Zuilen, M.A.; Medjoubi, K.; Gérard, E.; Jamme, F.; Dupraz, C.; Braissant, O. Patterns of Metal Distribution in Hypersaline Microbialites during Early Diagenesis: Implications for the Fossil Record. Geobiology 2017, 15, 259–279. [Google Scholar] [CrossRef]
- Korotev, R.L.; Zeigler, R.A.; Jolliff, B.L.; Irvin, A.J.; Bunch, T.E. Compositional and Lithological Diversity among Brecciated Lunar Meteorites of Intermediate Iron Concentration. Meteorit. Planet. Sci. 2009, 44, 1287–1322. [Google Scholar] [CrossRef]
- Wilson, R.G. Secondary Ion Mass Spectrometry Sensitivity Factors versus Ionization Potential and Electron Affinity for Many Elements in HgCdTe and CdTe Using Oxygen and Cesium Ion Beams. J. Appl. Phys. 1988, 63, 5121–5125. [Google Scholar] [CrossRef]
- Johnson, D.B. Biohydrometallurgy and the Environment: Intimate and Important Interplay. Hydrometallurgy 2006, 83, 153–166. [Google Scholar] [CrossRef]
- Lynne, B.Y.; Campbell, K.A.; Moore, J.N.; Browne, P.R.L. Diagenesis of 1900-Year-Old Siliceous Sinter (Opal-A to Quartz) at Opal Mound, Roosevelt Hot Springs, Utah, USA. Sediment. Geol. 2005, 179, 249–278. [Google Scholar] [CrossRef]
- Hickman-Lewis, K.; Gourcerol, B.; Westall, F.; Manzini, D.; Cavalazzi, B. Reconstructing Palaeoarchaean Microbial Biomes Flourishing in the Presence of Emergent Landmasses Using Trace and Rare Earth Element Systematics. Precambrian Res. 2020, 342, 105689. [Google Scholar] [CrossRef]
- Huerta-Diaz, M.A.; Delgadillo-Hinojosa, F.; Siqueiros-Valencia, A.; Valdivieso-Ojeda, J.; Reimer, J.J.; Segovia-Zavala, J.A. Millimeter-scale Resolution of Trace Metal Distributions in Microbial Mats from a Hypersaline Environment in Baja California, Mexico. Geobiology 2012, 10, 531–547. [Google Scholar] [CrossRef] [PubMed]
- Allwood, A.C.; Kamber, B.S.; Walter, M.R.; Burch, I.W.; Kanik, I. Trace Elements Record Depositional History of an Early Archean Stromatolitic Carbonate Platform. Chem. Geol. 2010, 270, 148–163. [Google Scholar] [CrossRef]




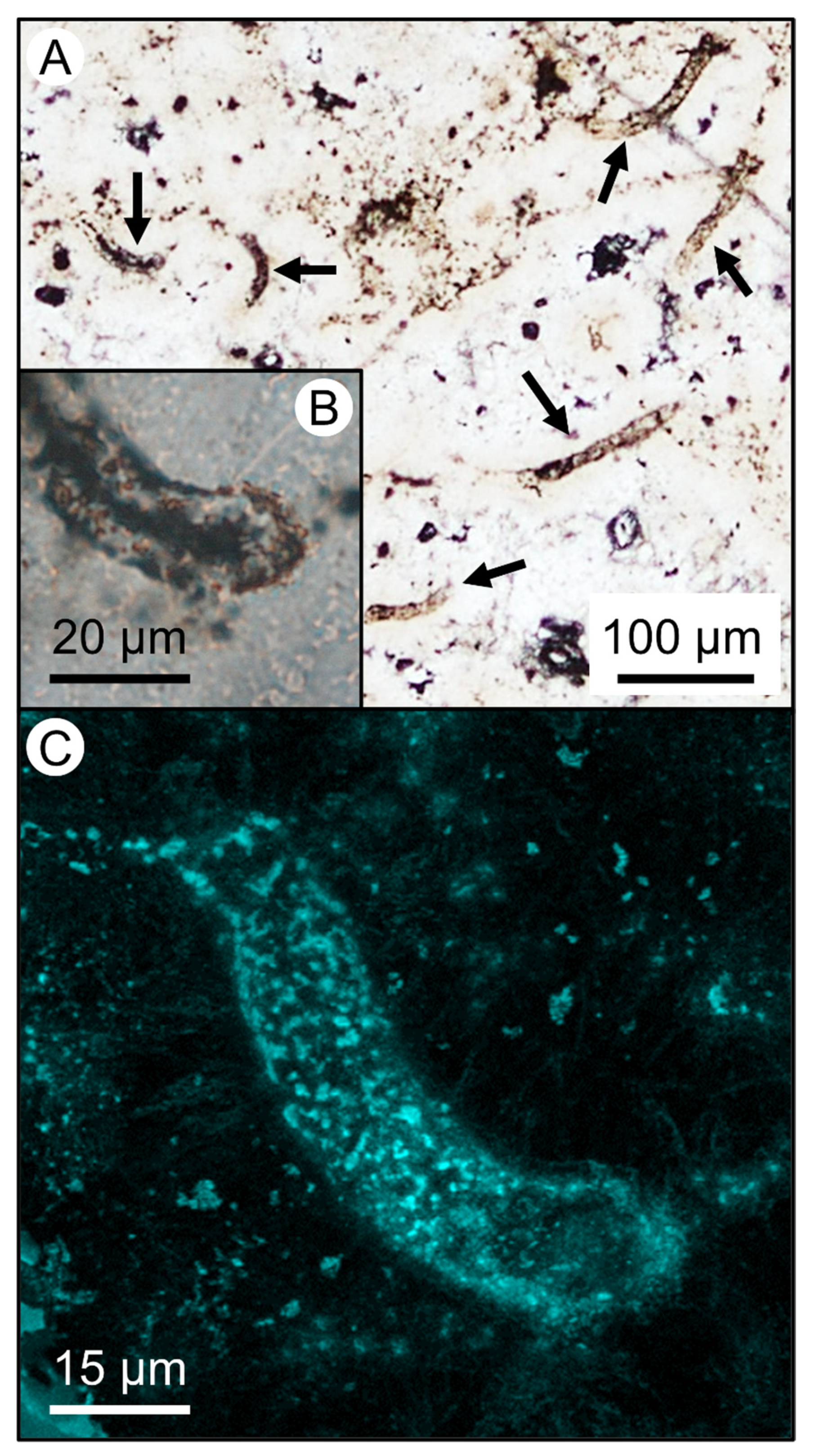{kind=link}
{kind=link}
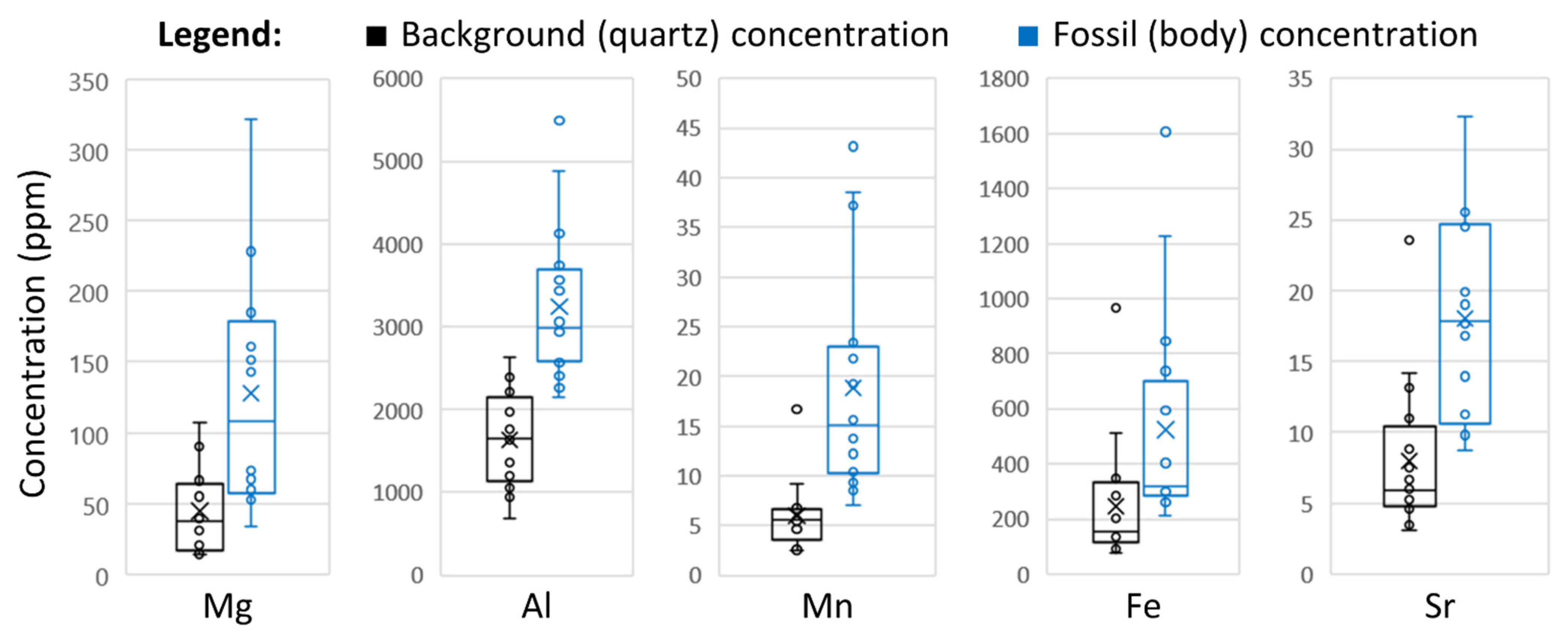{kind=link}
| Component | Mass % |
|---|---|
| SiO2 | 95.7379 |
| Al2O3 | 1.6859 |
| Fe2O3 | 0.2517 |
| MgO | 0.2249 |
| Sr | 0.012 |
| MnO | 0.0085 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gangidine, A.; Walter, M.R.; Havig, J.R.; Jones, C.; Sturmer, D.M.; Czaja, A.D. Trace Element Concentrations Associated with Mid-Paleozoic Microfossils as Biosignatures to Aid in the Search for Life. Life 2021, 11, 142. https://doi.org/10.3390/life11020142
Gangidine A, Walter MR, Havig JR, Jones C, Sturmer DM, Czaja AD. Trace Element Concentrations Associated with Mid-Paleozoic Microfossils as Biosignatures to Aid in the Search for Life. Life. 2021; 11(2):142. https://doi.org/10.3390/life11020142
Chicago/Turabian StyleGangidine, Andrew, Malcolm R. Walter, Jeff R. Havig, Clive Jones, Daniel M. Sturmer, and Andrew D. Czaja. 2021. "Trace Element Concentrations Associated with Mid-Paleozoic Microfossils as Biosignatures to Aid in the Search for Life" Life 11, no. 2: 142. https://doi.org/10.3390/life11020142