
Coupling miR/isomiR and mRNA Expression Signatures Unveils New Molecular Layers of Endometrial Receptivity

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients, Study Design, and Samples
2.2. RNA Extraction and Quality Control
2.3. sRNA-Seq and Data Analysis
2.4. mRNA Sequencing and Data Analysis
2.5. Prediction of miR/isomiR Interactions with Target Transcripts and Transcription Factors
2.6. Gene Ontology and Pathway Analysis
3. Results
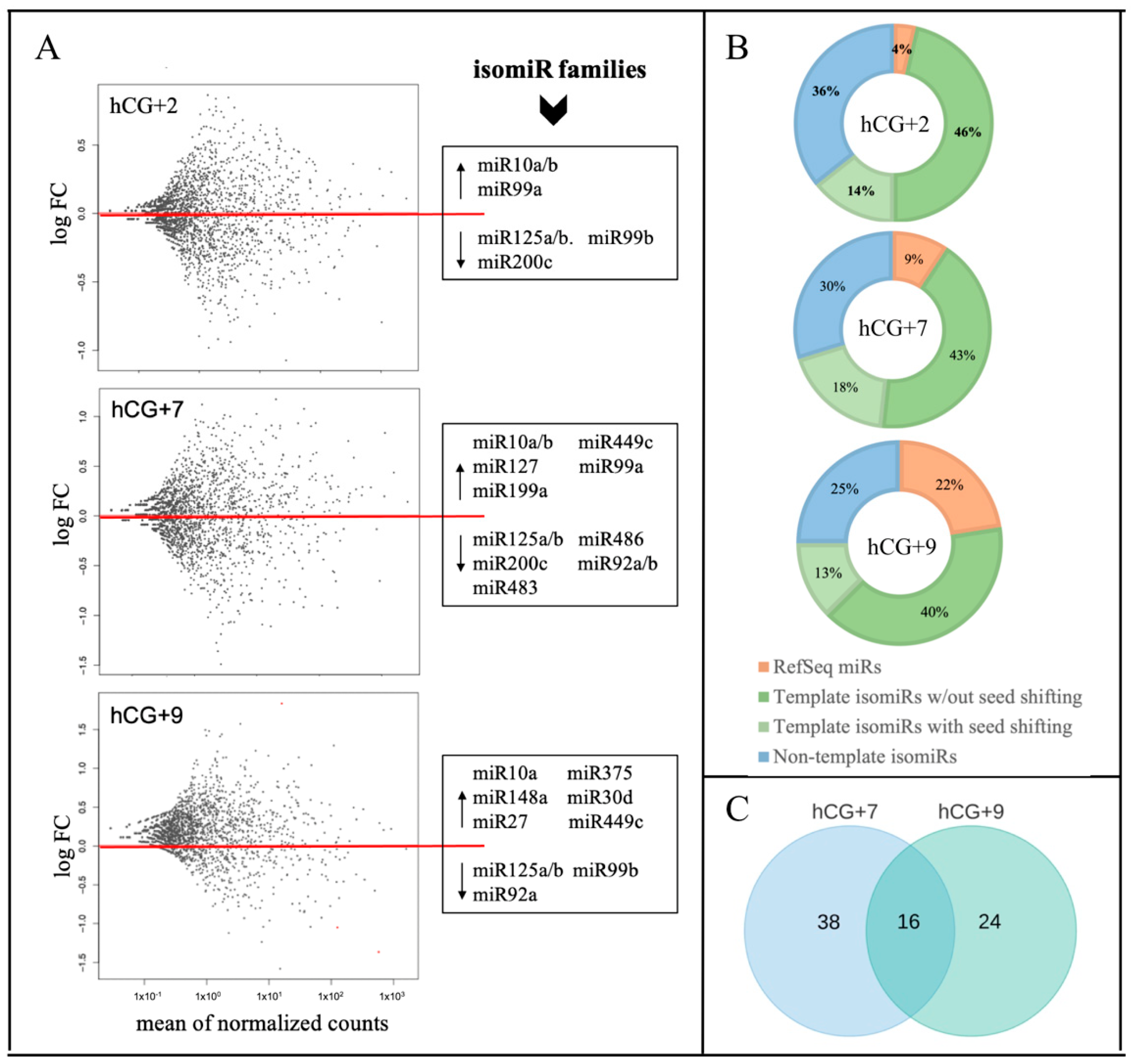
3.1. sRNA-Seq and Identification of DE miRs and isomiRs During the Phase Transitions of Endometrial Cycle
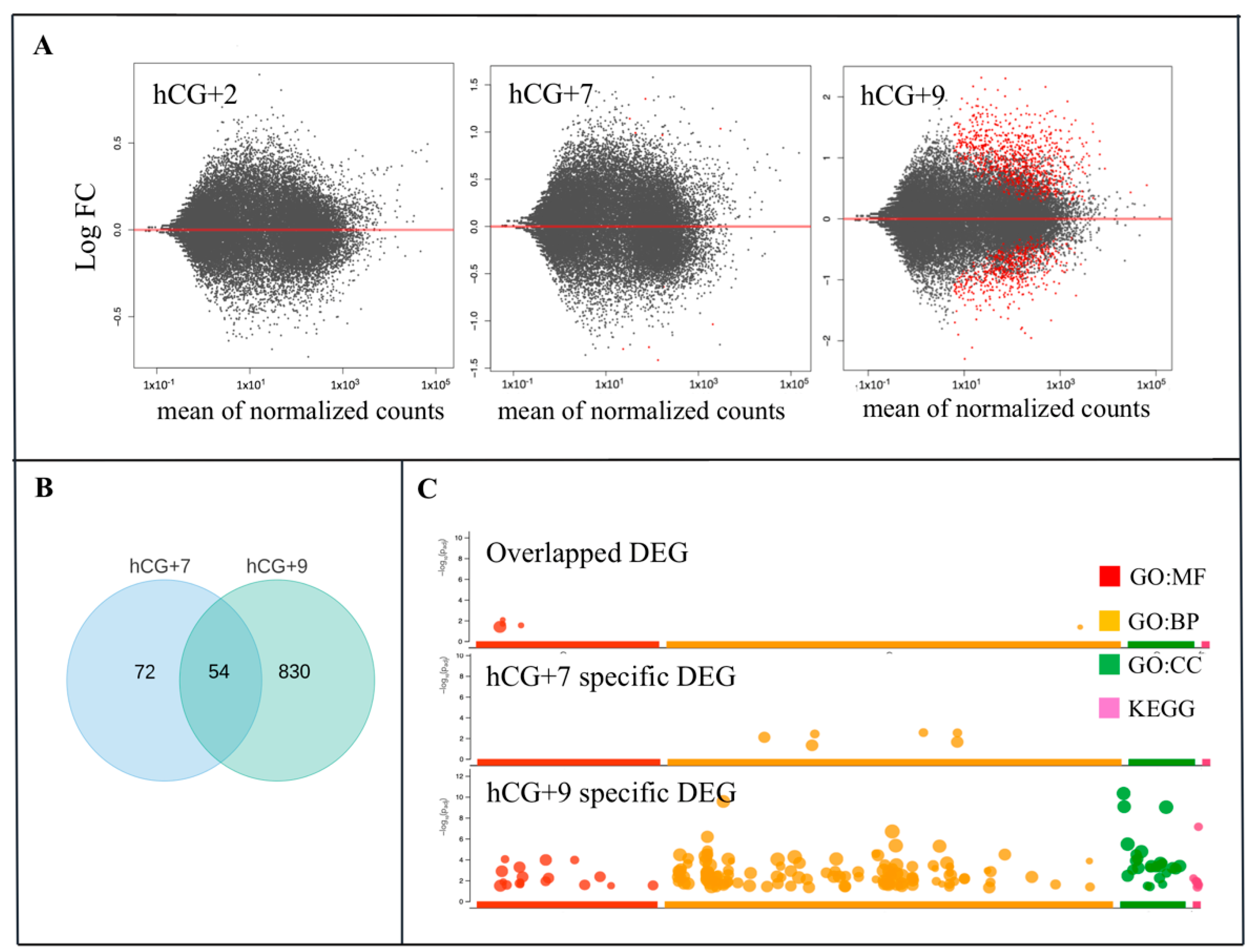
3.2. mRNA-Seq and Identification of DEGs during the Phase Transitions of Endometrial Cycle
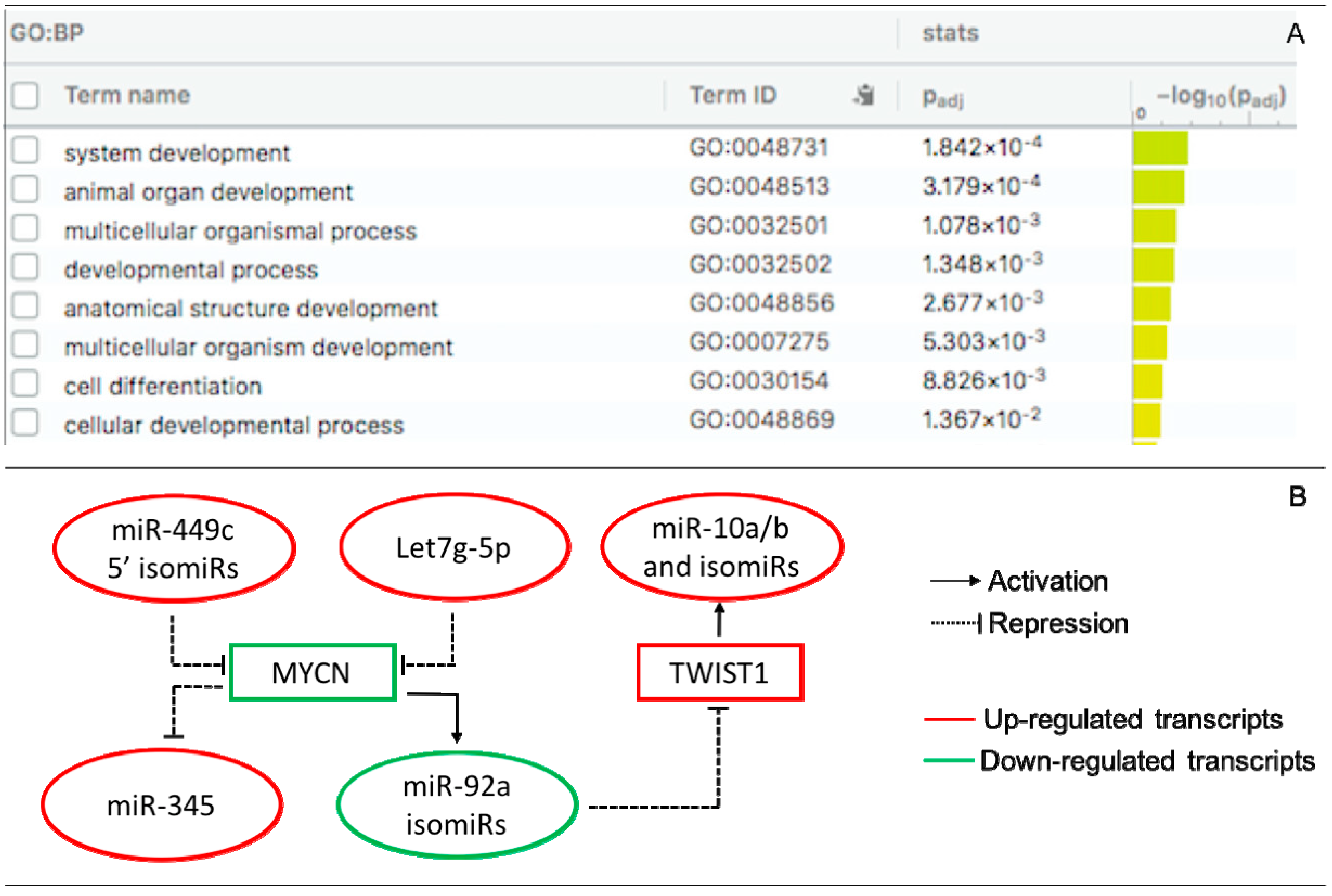
3.3. Interactions of DE miRs and isomiRs with DEGs in the Receptive Endometrium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, R.C.; Ambros, V. An Extensive Class of Small RNAs in Caenorhabditis Elegans. Science 2001, 294, 862–864. [Google Scholar] [CrossRef] [Green Version]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of Novel Genes Coding for Small Expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of Mammalian MicroRNA Host Genes and Transcription Units. Genome Res. 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [Green Version]
- Ha, M.; Kim, V.N. Regulation of MicroRNA Biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA Genes Are Transcribed by RNA Polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Jones-Rhoades, M.W.; Bartel, D.P.; Bartel, B. MicroRNAs and Their Regulatory Roles in Plants. Annu. Rev. Plant Biol. 2006, 57, 19–53. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Rådmark, O.; Kim, S.; et al. The Nuclear RNase III Drosha Initiates MicroRNA Processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Gregory, R.I.; Yan, K.P.; Amuthan, G.; Chendrimada, T.; Doratotaj, B.; Cooch, N.; Shiekhattar, R. The Microprocessor Complex Mediates the Genesis of MicroRNAs. Nature 2004, 432. [Google Scholar] [CrossRef]
- Zhang, H.; Kolb, F.A.; Jaskiewicz, L.; Westhof, E.; Filipowicz, W. Single Processing Center Models for Human Dicer and Bacterial RNase III. Cell 2004, 118, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; Zamore, P.D. MicroPrimer: The Biogenesis and Function of MicroRNA. Development 2005, 132, 4645–4652. [Google Scholar] [CrossRef] [Green Version]
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of MRNA Translation and Stability by MicroRNAs. Annu. Rev. Biochem. 2010, 79, 351–379. [Google Scholar] [CrossRef] [Green Version]
- Okamura, K.; Hagen, J.W.; Duan, H.; Tyler, D.M.; Lai, E.C. The Mirtron Pathway Generates MicroRNA-Class Regulatory RNAs in Drosophila. Cell 2007, 130. [Google Scholar] [CrossRef] [Green Version]
- Ruby, J.G.; Jan, C.H.; Bartel, D.P. Intronic MicroRNA Precursors That Bypass Drosha Processing. Nature 2007, 448. [Google Scholar] [CrossRef] [Green Version]
- Cifuentes, D.; Xue, H.; Taylor, D.W.; Patnode, H.; Mishima, Y.; Cheloufi, S.; Ma, E.; Mane, S.; Hannon, G.J.; Lawson, N.D.; et al. A Novel MiRNA Processing Pathway Independent of Dicer Requires Argonaute2 Catalytic Activity. Science 2010, 328. [Google Scholar] [CrossRef] [Green Version]
- Cheloufi, S.; dos Santos, C.O.; Chong, M.M.W.; Hannon, G.J. A Dicer-Independent MiRNA Biogenesis Pathway That Requires Ago Catalysis. Nature 2010, 465. [Google Scholar] [CrossRef] [Green Version]
- Burroughs, A.M.; Ando, Y.; de Hoon, M.L.; Tomaru, Y.; Suzuki, H.; Hayashizaki, Y.; Daub, C.O. Deep-Sequencing of Human Argonaute-Associated Small RNAs Provides Insight into MiRNA Sorting and Reveals Argonaute Association with RNA Fragments of Diverse Origin. RNA Biol. 2011, 8. [Google Scholar] [CrossRef]
- Neilsen, C.T.; Goodall, G.J.; Bracken, C.P. IsomiRs—The Overlooked Repertoire in the Dynamic MicroRNAome. Trends Genet. 2012, 28, 544–549. [Google Scholar] [CrossRef]
- Tan, G.C.; Chan, E.; Molnar, A.; Sarkar, R.; Alexieva, D.; Isa, I.M.; Robinson, S.; Zhang, S.; Ellis, P.; Langford, C.F.; et al. 5′ IsomiR Variation Is of Functional and Evolutionary Importance. Nucleic Acids Res. 2014, 42. [Google Scholar] [CrossRef]
- Desvignes, T.; Batzel, P.; Berezikov, E.; Eilbeck, K.; Eppig, J.T.; McAndrews, M.S.; Singer, A.; Postlethwait, J.H. MiRNA Nomenclature: A View Incorporating Genetic Origins, Biosynthetic Pathways, and Sequence Variants. Trends Genet. 2015, 31, 613–626. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Yi, R.; Cullen, B.R. Recognition and Cleavage of Primary MicroRNA Precursors by the Nuclear Processing Enzyme Drosha. EMBO J. 2005, 24, 138–148. [Google Scholar] [CrossRef]
- Wu, H.; Ye, C.; Ramirez, D.; Manjunath, N. Alternative Processing of Primary MicroRNA Transcripts by Drosha Generates 5′ End Variation of Mature MicroRNA. PLoS ONE 2009, 4. [Google Scholar] [CrossRef]
- Park, J.E.; Heo, I.; Tian, Y.; Simanshu, D.K.; Chang, H.; Jee, D.; Patel, D.J.; Kim, V.N. Dicer Recognizes the 5′ End of RNA for Efficient and Accurate Processing. Nature 2011, 475, 201–205. [Google Scholar] [CrossRef]
- Kim, B.; Jeong, K.; Kim, V.N. Genome-Wide Mapping of DROSHA Cleavage Sites on Primary MicroRNAs and Noncanonical Substrates. Mol. Cell 2017, 66, 258–269.e5. [Google Scholar] [CrossRef] [Green Version]
- Heo, I.; Joo, C.; Kim, Y.K.; Ha, M.; Yoon, M.J.; Cho, J.; Yeom, K.H.; Han, J.; Kim, V.N. TUT4 in Concert with Lin28 Suppresses MicroRNA Biogenesis through Pre-MicroRNA Uridylation. Cell 2009, 138, 696–708. [Google Scholar] [CrossRef] [Green Version]
- Hagan, J.P.; Piskounova, E.; Gregory, R.I. Lin28 Recruits the TUTase Zcchc11 to Inhibit Let-7 Maturation in Mouse Embryonic Stem Cells. Nat. Struct. Mol. Biol. 2009, 16, 1021–1025. [Google Scholar] [CrossRef] [Green Version]
- Burroughs, A.M.; Ando, Y.; de Hoon, M.J.L.; Tomaru, Y.; Nishibu, T.; Ukekawa, R.; Funakoshi, T.; Kurokawa, T.; Suzuki, H.; Hayashizaki, Y.; et al. A Comprehensive Survey of 3′ Animal MiRNA Modification Events and a Possible Role for 3′ Adenylation in Modulating MiRNA Targeting Effectiveness. Genome Res. 2010, 20. [Google Scholar] [CrossRef] [Green Version]
- Wyman, S.K.; Knouf, E.C.; Parkin, R.K.; Fritz, B.R.; Lin, D.W.; Dennis, L.M.; Krouse, M.A.; Webster, P.J.; Tewari, M. Post-Transcriptional Generation of MiRNA Variants by Multiple Nucleotidyl Transferases Contributes to MiRNA Transcriptome Complexity. Genome Res. 2011, 21, 1450–1461. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Chendrimada, T.P.; Wang, Q.; Higuchi, M.; Seeburg, P.H.; Shiekhattar, R.; Nishikura, K. Modulation of MicroRNA Processing and Expression through RNA Editing by ADAR Deaminases. Nat. Struct. Mol. Biol. 2006, 13. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, Y.; Zinshteyn, B.; Sethupathy, P.; Iizasa, H.; Hatzigeorgiou, A.G.; Nishikura, K. Redirection of Silencing Targets by Adenosine-to-Inosine Editing of MiRNAs. Science 2007, 315. [Google Scholar] [CrossRef] [Green Version]
- Habig, J.W.; Dale, T.; Bass, B.L. MiRNA Editing-We Should Have Inosine This Coming. Mol. Cell 2007, 25, 792–793. [Google Scholar] [CrossRef] [Green Version]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of MiRNAs and SiRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Coller, J. What Comes First: Translational Repression or MRNA Degradation? The Deepening Mystery of MicroRNA Function. Cell Res. 2012, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphreys, D.T.; Hynes, C.J.; Patel, H.R.; Wei, G.H.; Cannon, L.; Fatkin, D.; Suter, C.M.; Clancy, J.L.; Preiss, T. Complexity of Murine Cardiomyocyte MiRNA Biogenesis, Sequence Variant Expression and Function. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telonis, A.G.; Loher, P.; Jing, Y.; Londin, E.; Rigoutsos, I. Beyond the One-Locus-One-MiRNA Paradigm: MicroRNA Isoforms Enable Deeper Insights into Breast Cancer Heterogeneity. Nucleic Acids Res. 2015, 43. [Google Scholar] [CrossRef] [Green Version]
- Telonis, A.G.; Magee, R.; Loher, P.; Chervoneva, I.; Londin, E.; Rigoutsos, I. Knowledge about the Presence or Absence of MiRNA Isoforms (IsomiRs) Can Successfully Discriminate amongst 32 TCGA Cancer Types. Nucleic Acids Res. 2017, 45. [Google Scholar] [CrossRef] [Green Version]
- Salem, O.; Erdem, N.; Jung, J.; Münstermann, E.; Wörner, A.; Wilhelm, H.; Wiemann, S.; Körner, C. The Highly Expressed 5′isomiR of Hsa-MiR-140-3p Contributes to the Tumor-Suppressive Effects of MiR-140 by Reducing Breast Cancer Proliferation and Migration. BMC Genom. 2016, 17. [Google Scholar] [CrossRef] [Green Version]
- Mercey, O.; Popa, A.; Cavard, A.; Paquet, A.; Chevalier, B.; Pons, N.; Magnone, V.; Zangari, J.; Brest, P.; Zaragosi, L.E.; et al. Characterizing IsomiR Variants within the MicroRNA-34/449 Family. FEBS Lett. 2017, 591. [Google Scholar] [CrossRef] [Green Version]
- Loher, P.; Londin, E.R.; Rigoutsos, I. IsomiR Expression Profiles in Human Lymphoblastoid Cell Lines Exhibit Population and Gender Dependencies. Oncotarget 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Altmäe, S.; Martinez-Conejero, J.A.; Esteban, F.J.; Ruiz-Alonso, M.; Stavreus-Evers, A.; Horcajadas, J.A.; Salumets, A. MicroRNAs MiR-30b, MiR-30d, and MiR-494 Regulate Human Endometrial Receptivity. Reprod. Sci. 2013, 20. [Google Scholar] [CrossRef] [Green Version]
- Vilella, F.; Moreno-Moya, J.M.; Balaguer, N.; Grasso, A.; Herrero, M.; Martínez, S.; Marcilla, A.; Simón, C. Hsa-MiR-30d, Secreted by the Human Endometrium, Is Taken up by the Pre-Implantation Embryo and Might Modify Its Transcriptome. Development 2015, 142. [Google Scholar] [CrossRef] [Green Version]
- Sha, A.G.; Liu, J.L.; Jiang, X.M.; Ren, J.Z.; Ma, C.H.; Lei, W.; Su, R.W.; Yang, Z.M. Genome-Wide Identification of Micro-Ribonucleic Acids Associated with Human Endometrial Receptivity in Natural and Stimulated Cycles by Deep Sequencing. Fertil. Steril. 2011, 96. [Google Scholar] [CrossRef]
- Altmäe, S.; Koel, M.; Võsa, U.; Adler, P.; Suhorutšenko, M.; Laisk-Podar, T.; Kukushkina, V.; Saare, M.; Velthut-Meikas, A.; Krjutškov, K.; et al. Meta-Signature of Human Endometrial Receptivity: A Meta-Analysis and Validation Study of Transcriptomic Biomarkers. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Sigurgeirsson, B.; Åmark, H.; Jemt, A.; Ujvari, D.; Westgren, M.; Lundeberg, J.; Gidlöf, S. Comprehensive RNA Sequencing of Healthy Human Endometrium at Two Time Points of the Menstrual Cycle. Biol. Reprod. 2017, 96. [Google Scholar] [CrossRef]
- Rekker, K.; Altmäe, S.; Suhorutshenko, M.; Peters, M.; Martinez-Blanch, J.F.; Codoñer, F.M.; Vilella, F.; Simón, C.; Salumets, A.; Velthut-Meikas, A. A Two-Cohort RNA-Seq Study Reveals Changes in Endometrial and Blood Mirnome in Fertile and Infertile Women. Genes 2018, 9, 574. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Kim, H.R.; Lim, E.J.; Park, M.; Yoon, J.A.; Kim, Y.S.; Kim, E.K.; Shin, J.E.; Kim, J.H.; Kwon, H.; et al. Integrative Analyses of Uterine Transcriptome and MicroRNAome Reveal Compromised LIF-STAT3 Signaling and Progesterone Response in the Endometrium of Patients with Recurrent/Repeated Implantation Failure (RIF). PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Shen, H.; Fan, L.J.; Guan, J.; Zheng, X.B.; Chen, X.; Liang, R.; Zhang, X.W.; Cui, Q.H.; Sun, K.K.; et al. Endometrial MicroRNA Signature during the Window of Implantation Changed in Patients with Repeated Implantation Failure. Chin. Med J. 2017, 130. [Google Scholar] [CrossRef]
- Faridani, O.R.; Abdullayev, I.; Hagemann-Jensen, M.; Schell, J.P.; Lanner, F.; Sandberg, R. Single-Cell Sequencing of the Small-RNA Transcriptome. Nat. Biotechnol. 2016, 34. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Ech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy Platform for Accessible, Reproducible and Collaborative Biomedical Analyses: 2018 Update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Glogovitis, I.; Yahubyan, G.; Würdinger, T.; Koppers-Lalic, D.; Baev, V. MiRGalaxy: Galaxy-Based Framework for Interactive Analysis of MicroRNA and IsomiR Sequencing Data. Cancers 2021, 13, 5663. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and Memory-Efficient Alignment of Short DNA Sequences to the Human Genome. Genome Biol. 2009, 10. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Teder, H.; Koel, M.; Paluoja, P.; Jatsenko, T.; Rekker, K.; Laisk-Podar, T.; Kukuškina, V.; Velthut-Meikas, A.; Fjodorova, O.; Peters, M.; et al. TAC-Seq: Targeted DNA and RNA Sequencing for Precise Biomarker Molecule Counting. NPJ Genom. Med. 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, X. MiRDB: An Online Database for Prediction of Functional MicroRNA Targets. Nucleic Acids Res. 2020, 48. [Google Scholar] [CrossRef] [Green Version]
- Tong, Z.; Cui, Q.; Wang, J.; Zhou, Y. TransmiR v2.0: An Updated Transcription Factor-MicroRNA Regulation Database. Nucleic Acids Res. 2019, 47. [Google Scholar] [CrossRef] [Green Version]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res. 2019, 47. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Kim, C.H.; Kim, D.Y.; Ahn, J.W.; Kim, S.H.; Chae, H.D.; Kang, B.M. Human Chorionic Gonadotropin-Administered Natural Cycle versus Spontaneous Ovulatory Cycle in Patients Undergoing Two Pronuclear Zygote Frozen-Thawed Embryo Transfer. Obstet. Gynecol. Sci. 2018, 61. [Google Scholar] [CrossRef]
- Dai, J.; Wang, J.; Yang, L.; Xiao, Y.; Ruan, Q. MIR-125a Regulates Angiogenesis of Gastric Cancer by Targeting Vascular Endothelial Growth Factor A. Int. J. Oncol. 2015, 47. [Google Scholar] [CrossRef]
- Jin, F.; Wang, Y.; Zhu, Y.; Li, S.; Liu, Y.; Chen, C.; Wang, X.; Zen, K.; Li, L. The MIR-125a/HK2 Axis Regulates Cancer Cell Energy Metabolism Reprogramming in Hepatocellular Carcinoma. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Wade, S.M.; Ohnesorge, N.; McLoughlin, H.; Biniecka, M.; Carter, S.P.; Trenkman, M.; Cunningham, C.C.; McGarry, T.; Canavan, M.; Kennedy, B.N.; et al. Dysregulated MiR-125a Promotes Angiogenesis through Enhanced Glycolysis. EBioMedicine 2019, 47. [Google Scholar] [CrossRef] [Green Version]
- Zhao, A.; Zeng, Q.; Xie, X.; Zhou, J.; Yue, W.; Li, Y.; Pei, X. MicroRNA-125b Induces Cancer Cell Apoptosis through Suppression of Bcl-2 Expression. J. Genet. Genom. 2012, 39. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.F.; He, T.Z.; Liu, C.M.; Cui, Y.; Song, P.P.; Jin, X.H.; Ma, X. MiR-125b Expression Affects the Proliferation and Apoptosis of Human Glioma Cells by Targeting Bmf. Cell Physiol. Biochem. 2009, 23. [Google Scholar] [CrossRef] [PubMed]
- Bonauer, A.; Carmona, G.; Iwasaki, M.; Mione, M.; Koyanagi, M.; Fischer, A.; Burchfield, J.; Fox, H.; Doebele, C.; Ohtani, K.; et al. MicroRNA-92a Controls Angiogenesis and Functional Recovery of Ischemic Tissues in Mice. Science 2009, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, K.; Ito, H.; Yoshitomi, H.; Yamamoto, K.; Fukuda, A.; Yoshikawa, J.; Furu, M.; Ishikawa, M.; Shibuya, H.; Matsuda, S. Inhibition of MiR-92a Enhances Fracture Healing via Promoting Angiogenesis in a Model of Stabilized Fracture in Young Mice. J. Bone Miner. Res. 2014, 29. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, L.P.; Grazul-Bilska, A.T.; Redmer, D.A. Angiogenesis in the Female Reproductive Organs: Pathological Implications. Int. J. Exp. Pathol. 2002, 83. [Google Scholar] [CrossRef]
- Jiajie, T.; Yanzhou, Y.; Hoi-Hung, A.C.; Chen, Z.J.; Wai-Yee, C. Conserved MiR-10 Family Represses Proliferation and Induces Apoptosis in Ovarian Granulosa Cells. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Sandbothe, M.; Buurman, R.; Reich, N.; Greiwe, L.; Vajen, B.; Gürlevik, E.; Schäffer, V.; Eilers, M.; Kühnel, F.; Vaquero, A.; et al. The MicroRNA-449 Family Inhibits TGF-β-Mediated Liver Cancer Cell Migration by Targeting SOX4. J. Hepatol. 2017, 66. [Google Scholar] [CrossRef]
- Schwab, M. MYCN in Neuronal Tumours. Cancer Lett. 2004, 204. [Google Scholar] [CrossRef]
- Schulte, J.H.; Horn, S.; Otto, T.; Samans, B.; Heukamp, L.C.; Eilers, U.C.; Krause, M.; Astrahantseff, K.; Klein-Hitpass, L.; Buettner, R.; et al. MYCN Regulates Oncogenic MicroRNAs in Neuroblastoma. Int. J. Cancer 2008, 122. [Google Scholar] [CrossRef]
- Haug, B.H.; Henriksen, J.R.; Buechner, J.; Geerts, D.; Tømte, E.; Kogner, P.; Martinsson, T.; Flægstad, T.; Sveinbjørnsson, B.; Einvik, C. MYCN-Regulated MiRNA-92 Inhibits Secretion of the Tumor Suppressor DICKKOPF-3 (DKK3) in Neuroblastoma. Carcinogenesis 2011, 32. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.L.; Chang, H.Y.; Chang, J.Y.; Hsu, W.M.; Huang, H.C.; Juan, H.F. Unveiling MYCN Regulatory Networks in Neuroblastoma via Integrative Analysis of Heterogeneous Genomics Data. Oncotarget 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Qin, Q.; Xu, Y.; He, T.; Qin, C.; Xu, J. Normal and Disease-Related Biological Functions of Twist1 and Underlying Molecular Mechanisms. Cell Res. 2012, 22. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, J.K.; Kessler, C.A.; Handwerger, S. Critical Role for TWIST1 in the Induction of Human Uterine Decidualization. Endocrinology 2011, 152. [Google Scholar] [CrossRef] [Green Version]
- Kuokkanen, S.; Chen, B.; Ojalvo, L.; Benard, L.; Santoro, N.; Pollard, J.W. Genomic Profiling of MicroRNAs and Messenger RNAs Reveals Hormonal Regulation in MicroRNA Expression in Human Endometrium. Biol. Reprod. 2010, 82. [Google Scholar] [CrossRef] [Green Version]
- Evans, G.E.; Martínez-Conejero, J.A.; Phillipson, G.T.M.; Simón, C.; McNoe, L.A.; Sykes, P.H.; Horcajadas, J.A.; Lam, E.Y.N.; Print, C.G.; Sin, I.L.; et al. Gene and Protein Expression Signature of Endometrial Glandular and Stromal Compartments during the Window of Implantation. Fertil. Steril. 2012, 97. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| DE miRs/isomiRs | DE Targets * | ||||
|---|---|---|---|---|---|
| Family Name | miR/isomiR Name | miR/isomiR Sequence | |||
| let-7g-5p ↑ | A | let-7g-5p | CUGUACAGGCCACUGCCUUGC | ↓ | MYCN2, KCNC2, FAXC |
| miR-10a/b ↑ | A | miR-10b-5p | UACCCUGUAGAACCGAAUUUGUG | ↓ | GABRB2 |
| B | miR-10a-5p_t_+1_-1 | ACCCUGUAGAUCCGAAUUUGU | ↓ | CPED1 | |
| miR-23a ↑ | A | miR-23a -3p_t_0_-1 | AUCACAUUGCCAGGGAUUUC | ↓ | FUT94, ROBO23, CRISPLD1, FREM1, ANO42, FBN2, KCNIP42, GUCY1A2, SH3BGR, ZNF730 |
| miR-27b ↑ | A | miR-27b-3p | UUCACAGUGGCUAAGUUCUGC | ↓ | EYA43, SFRP1, RARA2, BMP33, CCN42, SLC9A4, CYP39A12 |
| B | miR-27b-3p_t_+1_0 | UCACAGUGGCUAAGUUCUGC | ↓ | CYP39A12, LBH2, EYA43, RARA2, CCN42 | |
| miR-30d ↑ | A | miR-30d-5p_t_0_+2 | UGUAAACAUCCCCGACUGGAAG | ↓ | RTKN22, ANO42, SPOCK32, RAPGEF4, GRIA2, OVOL1, PAPOLB, GCLC2, CAMK4, NAP1L2, SEMA3A |
| B | miR-30d-5p_t_+1_+2 | GUAAACAUCCCCGACUGGAAGCU | ↓ | RTKN22, ROBO23, GCLC2, NDNF, ANG, SPOCK32, IFIT, FHOD3, MYH15, CSMD3, FUT94, ESCO2 | |
| miR-31 ↑ | A | miR-31-5p | AGGCAAGAUGCUGGCAUAGCU | ↓ | LBH2 |
| miR-92a ↓ | A | miR-92a-3p_nont_0_+2_AA | UAUUGCACUUGUCCCGGCCUGUAA | ↑ | GRHL1, TTC9, MTF12, IDH1, SOX11, MMP10, REXO1, IRS2, TWIST1, LRRC1, PIK3AP1 |
| miR-125a/b ↓ | A | miR-125a-5p_t_0_-2 | UCCCUGAGACCCUUUAACCUGU | ↑ | CYP24A1, MTF12, SLC7A1, ANKRD33B, PHACTR3, TMPRSS13, SOD2, IGSF11 |
| B | miR-125a-5p_t_+1_-2 | CCCUGAGACCCUUUAACCUGU | ↑ | TRNP1, DPP4, STAC2, CPT1A, ADCY1, ADAMTS8 | |
| miR-127 ↑ | A | miR-127-3p_nont_0_+2_AU | UCGGAUCCGUCUGAGCUUGGCUAU | ↓ | ATP1A2 |
| miR-141 ↑ | A | miR-141-3p | UAACACUGUCUGGUAAAGAUGG | ↓ | ELMOD13, NAP1L2, CNTN1, HCN1, MCIDAS2, TMEM130 |
| miR-148a ↑ | A | miR-148a-3p_t_0_+1 | UCAGUGCACUACAGAACUUUGUC | ↓ | ADGRB3, ROBO23, BMP33, ISM1 |
| miR-199a ↑ | A | miR-199a-5p | CCCAGUGUUCAGACUACCUGUUC | ↓ | HMCN1 |
| B | miR-199a-5p_t_+1_0 | CCAGUGUUCAGACUACCUGUUC | ↓ | PBK, EYA43, KCNIP4 | |
| miR-200c ↓ | A | miR-200c-3p_t_0_-3 | UAAUACUGCCGGGUAAUGAU | ↑ | PPP1R9B, NTRK2, KYNU, PITPNM3, PROK2, CDYL2, KLF6, SLC39A14, ADH1B, |
| miR-449c ↑ | A | miR-449c-5p_t_0_-2 | UAGGCAGUGUAUUGCUAGCGGCU | ↓ | ELMOD13, NEXMIF, FUT94 |
| B | miR-449c-5p_t_+1_-1 | AGGCAGUGUAUUGCUAGCGGCUG | ↓ | MYCN2. FUT94, ELMOD13, BMP33, ASIC2, NCEH1, MCIDAS2, | |
| miR-486 ↓ | A | miR-486-5p | UCCUGUACUGAGCUGCCCCGAG | ↑ | FGF7, CEMIP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikolova, M.; Naydenov, M.; Glogovitis, I.; Apostolov, A.; Saare, M.; Boggavarapu, N.; Salumets, A.; Baev, V.; Yahubyan, G. Coupling miR/isomiR and mRNA Expression Signatures Unveils New Molecular Layers of Endometrial Receptivity. Life 2021, 11, 1391. https://doi.org/10.3390/life11121391
Nikolova M, Naydenov M, Glogovitis I, Apostolov A, Saare M, Boggavarapu N, Salumets A, Baev V, Yahubyan G. Coupling miR/isomiR and mRNA Expression Signatures Unveils New Molecular Layers of Endometrial Receptivity. Life. 2021; 11(12):1391. https://doi.org/10.3390/life11121391
Chicago/Turabian StyleNikolova, Maria, Mladen Naydenov, Ilias Glogovitis, Apostol Apostolov, Merli Saare, Nageswara Boggavarapu, Andres Salumets, Vesselin Baev, and Galina Yahubyan. 2021. "Coupling miR/isomiR and mRNA Expression Signatures Unveils New Molecular Layers of Endometrial Receptivity" Life 11, no. 12: 1391. https://doi.org/10.3390/life11121391