
Isolation and Characterization of Bovine RVA from Northeast China, 2017–2020

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Primers
2.3. RT-PCR
2.4. Cell Culture and Virus Isolation
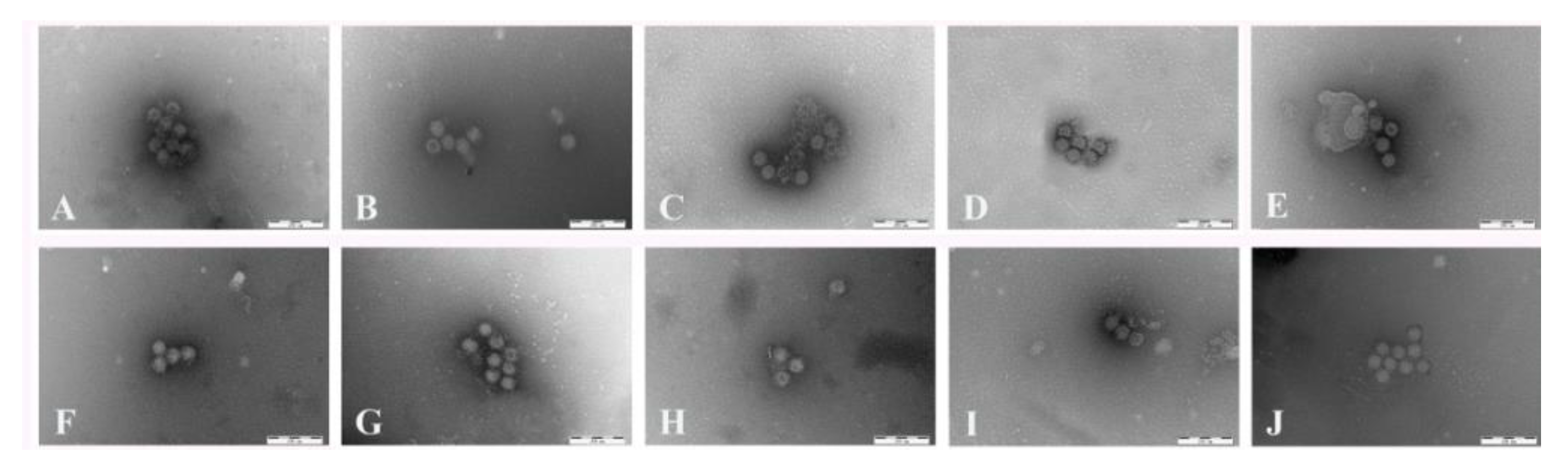
2.5. Immunoelectron Microscopy
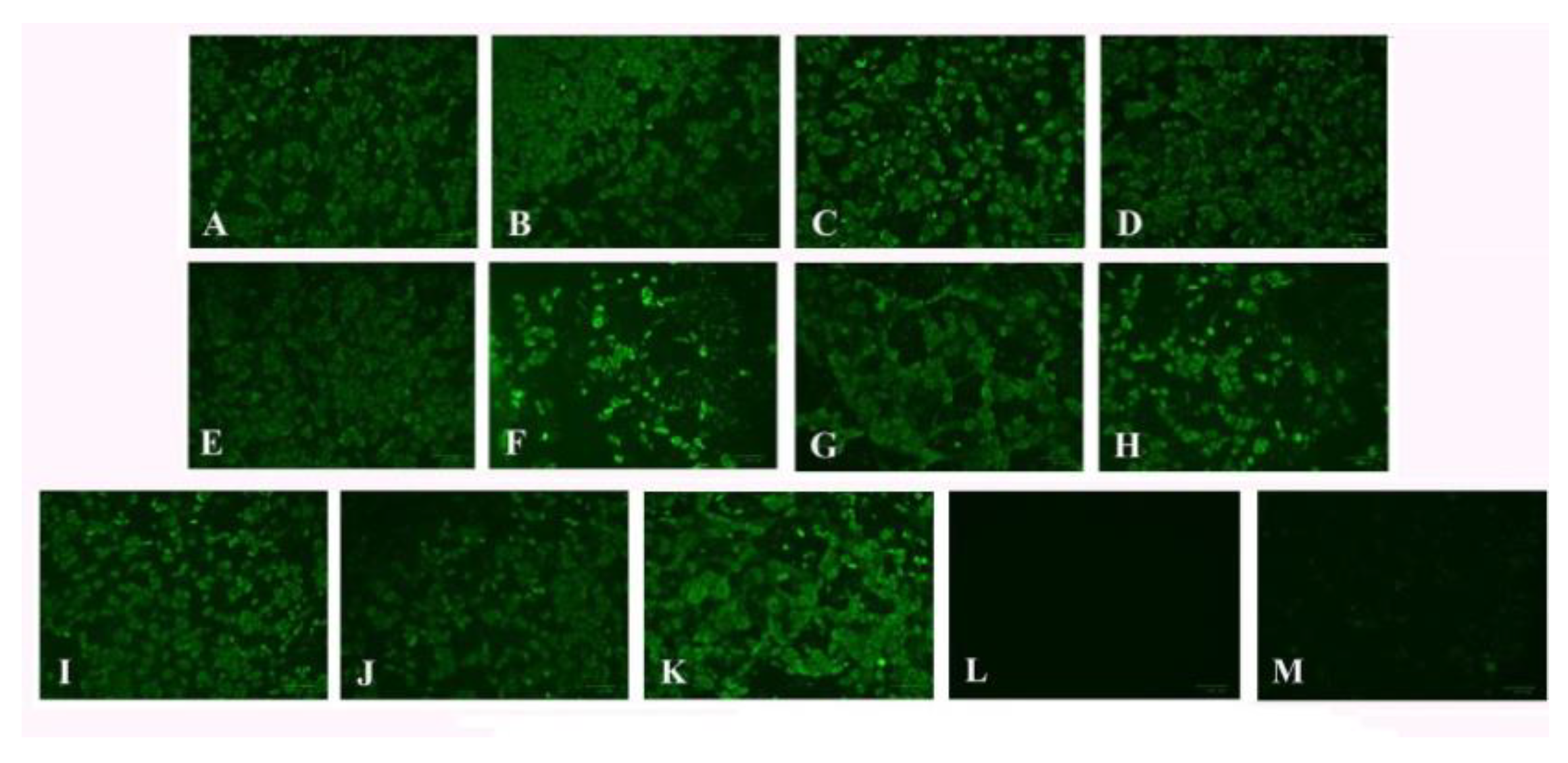
2.6. Indirect Immunofluorescence Assay
2.7. Sequence Alignments and Phylogenetic Analyses
3. Results
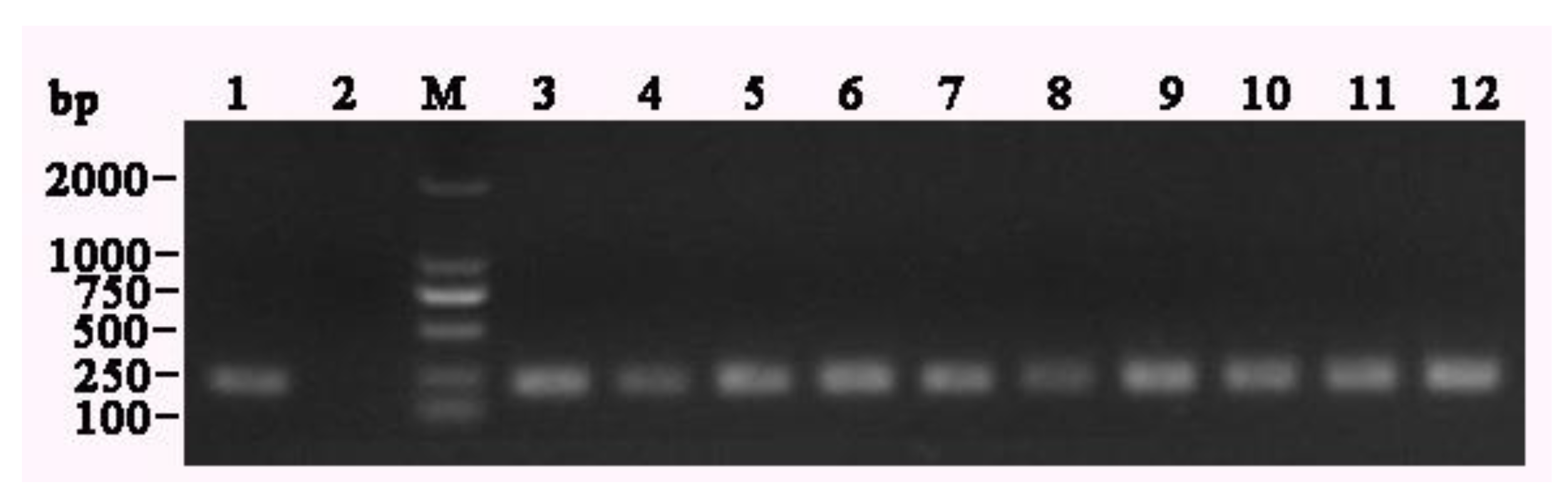
3.1. Molecular Characterization
3.2. Virus Isolation
3.3. Immunoelectron Microscopy
3.4. Indirect Immunofluorescence Assay
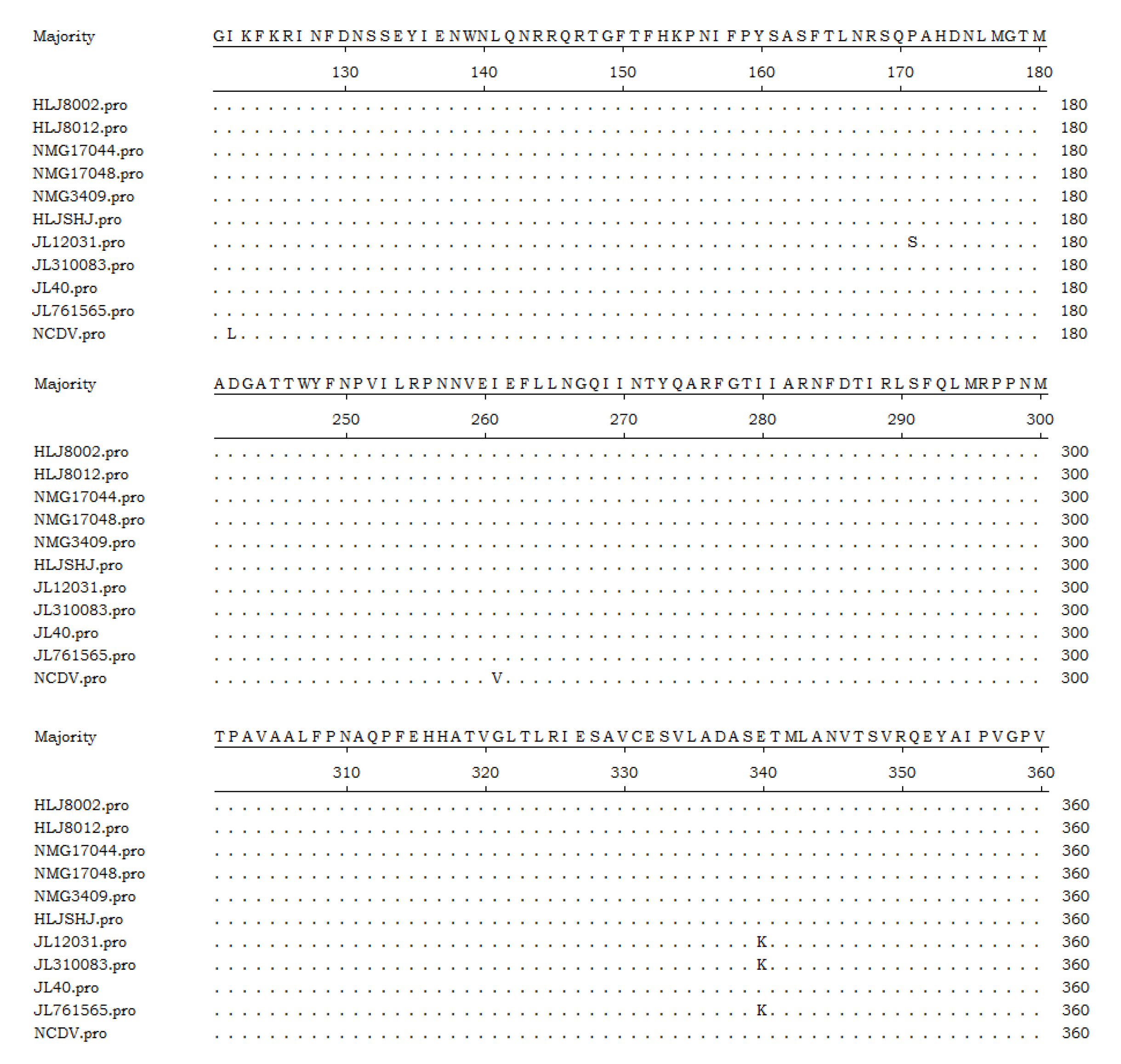
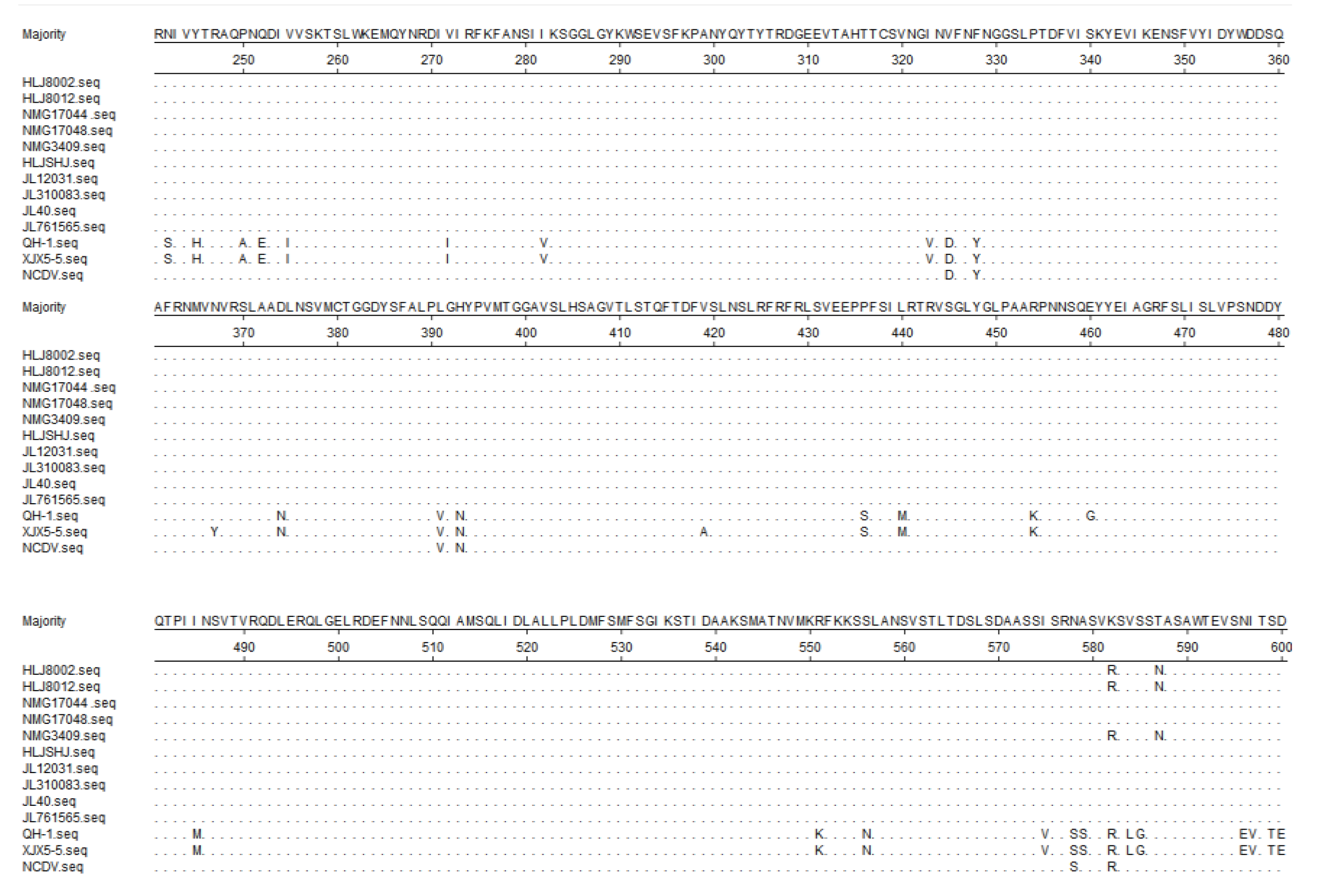
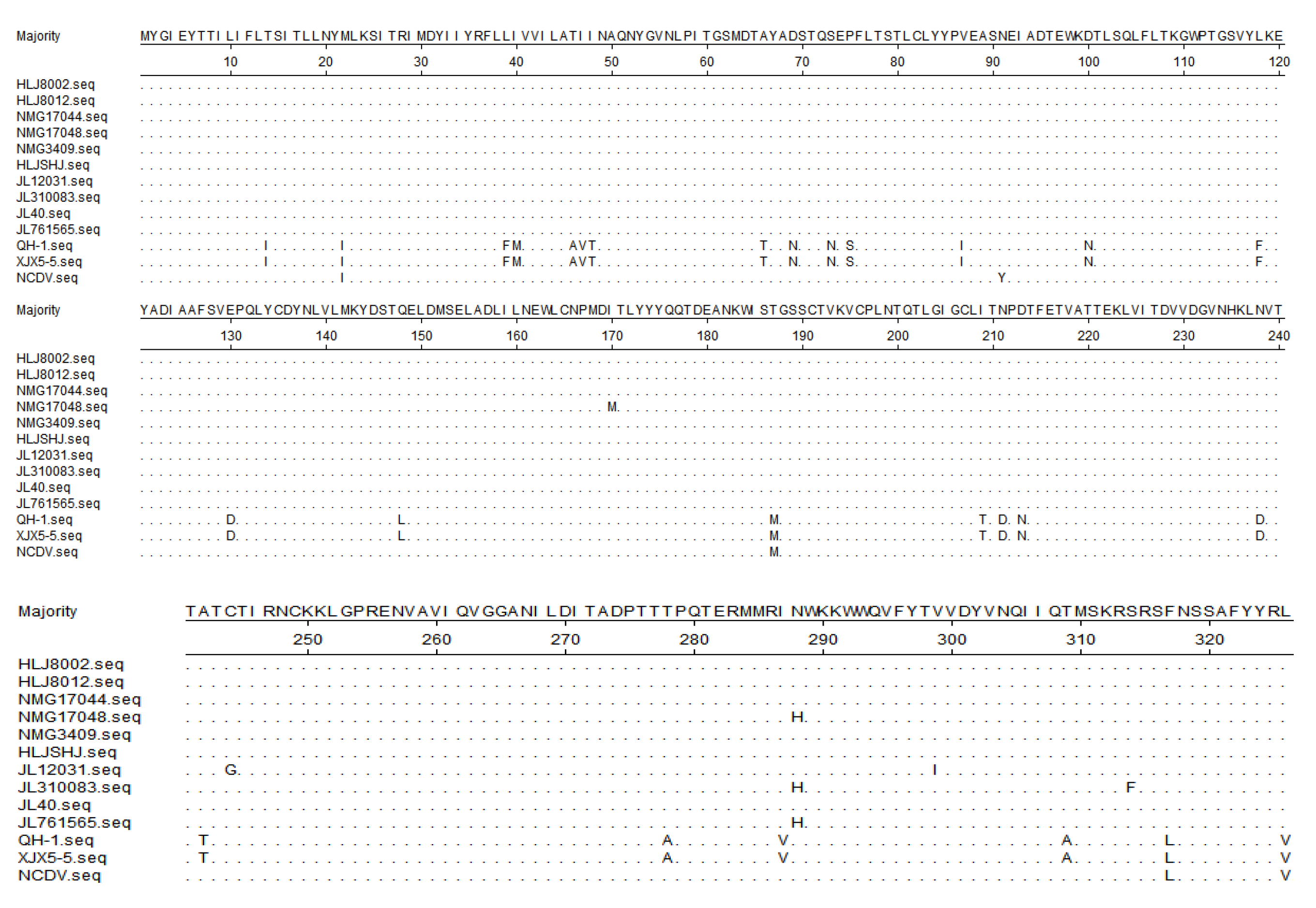
3.5. Sequence Analysis of the Target Genes
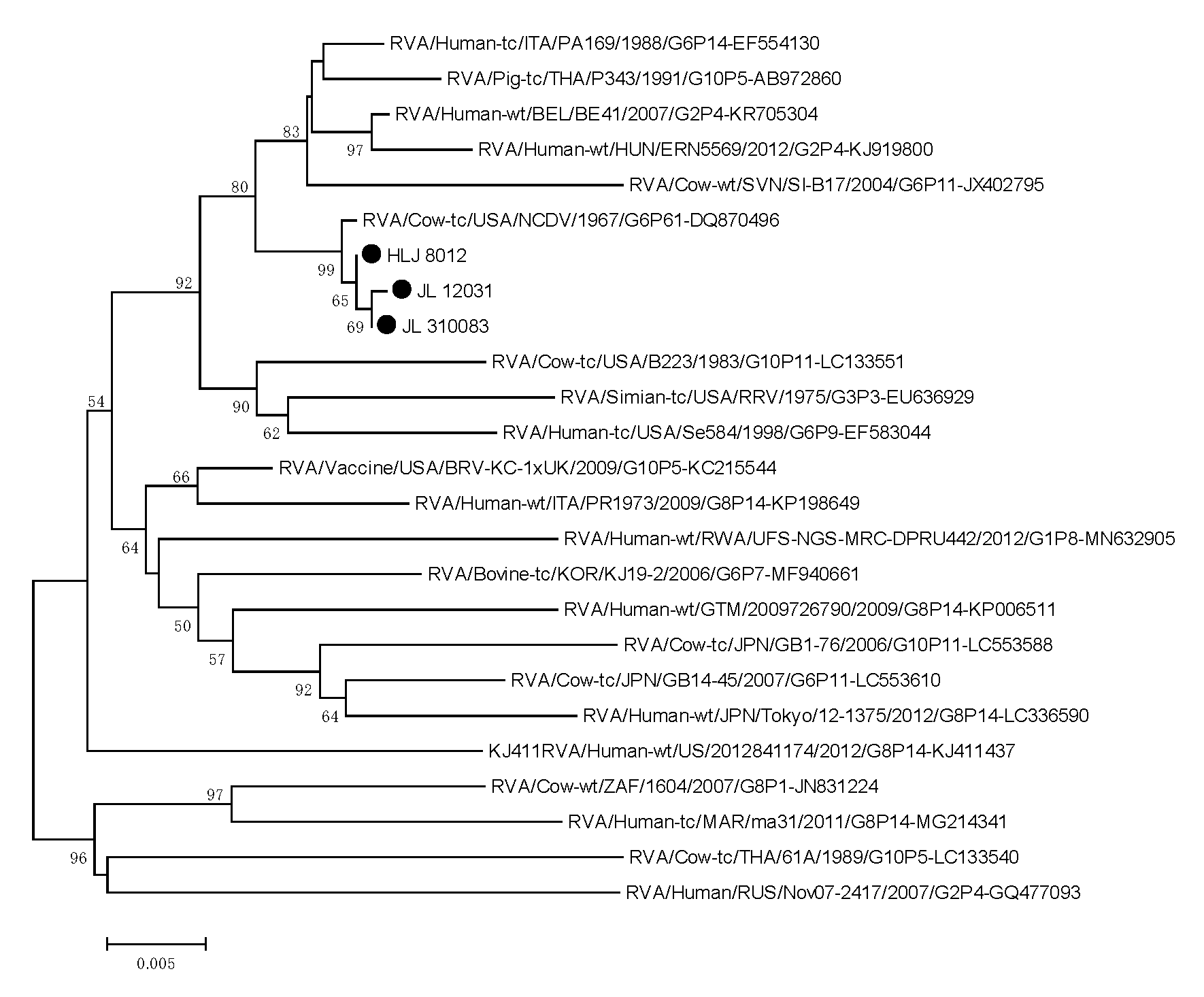
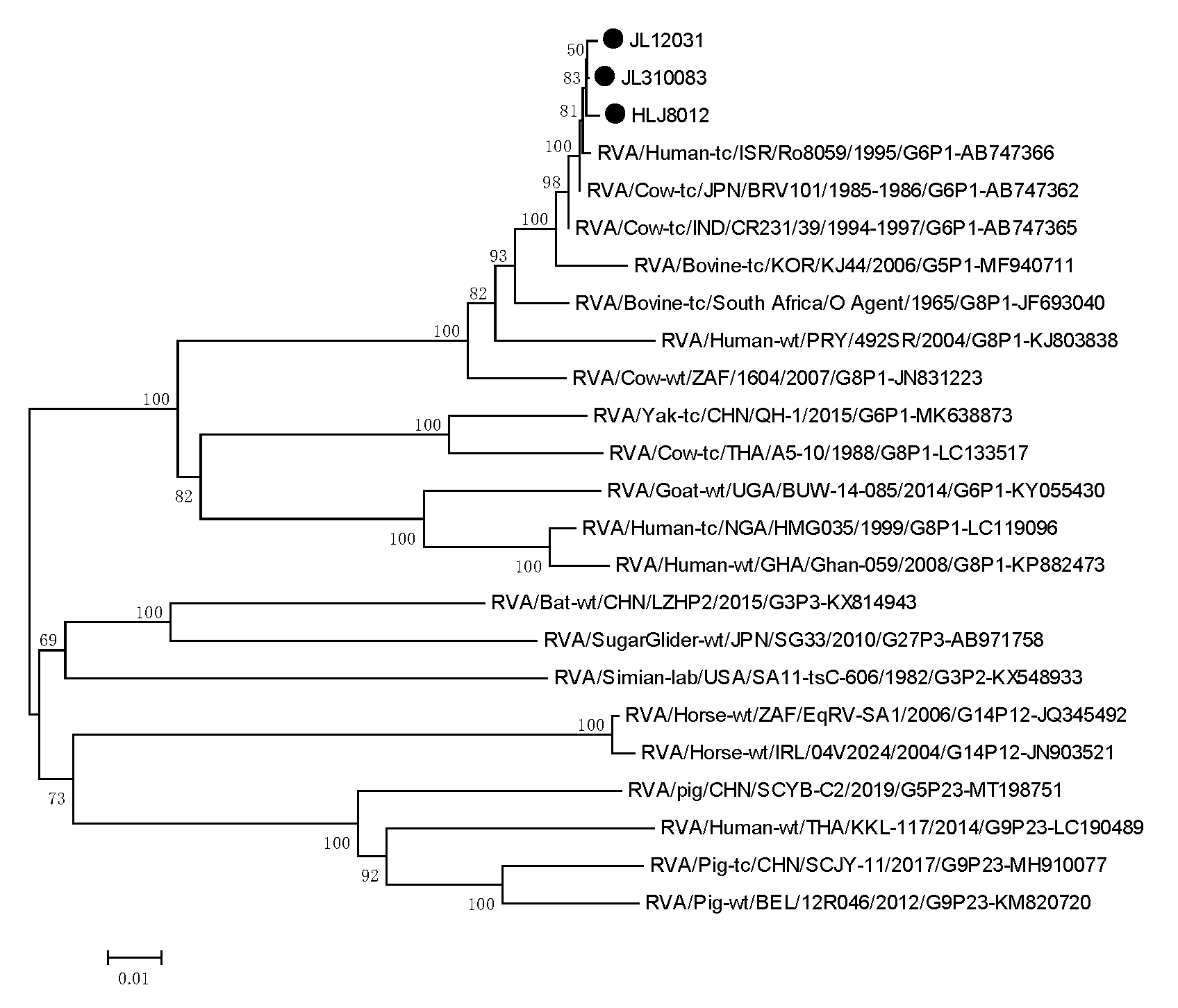
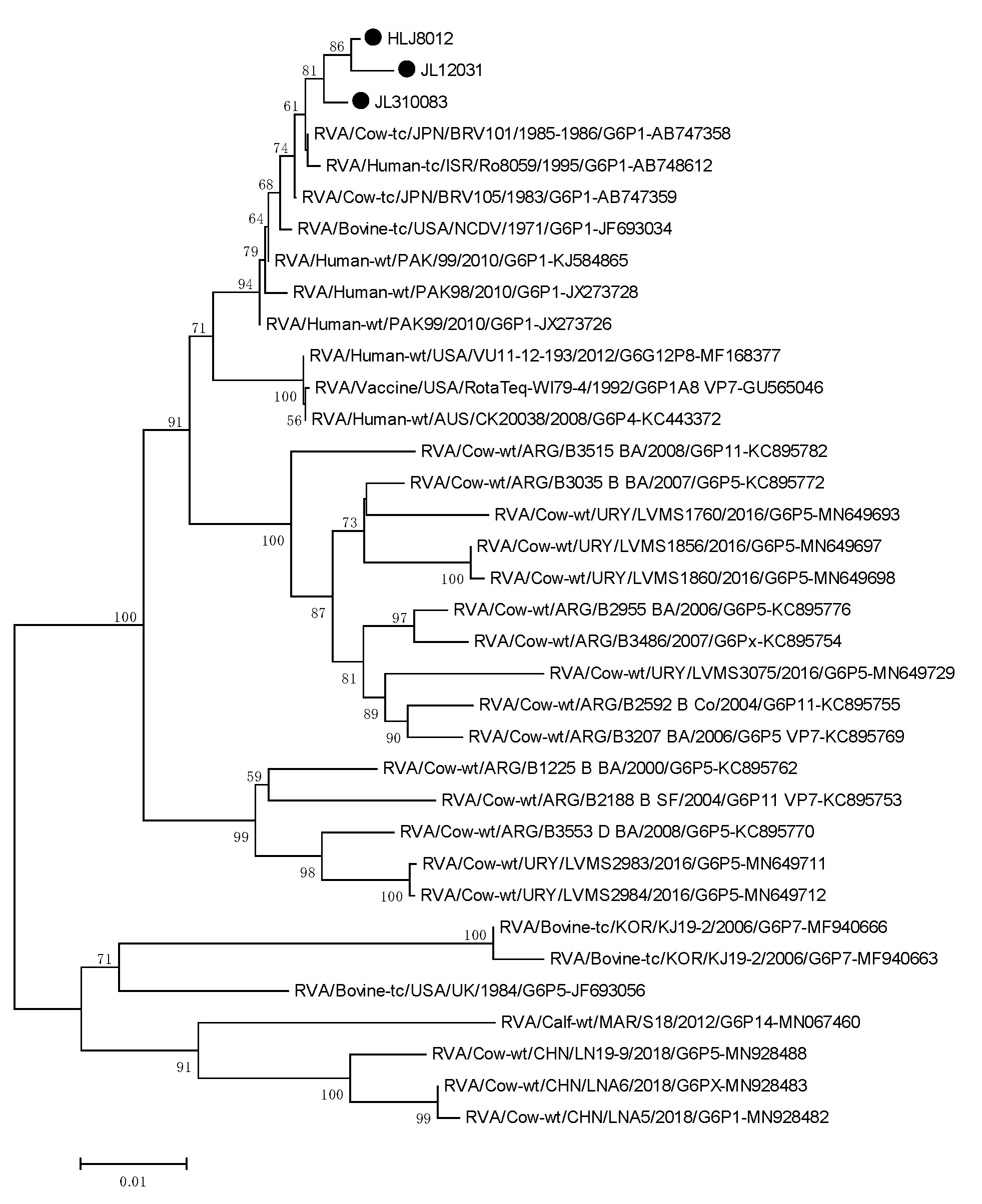
3.6. Phylogenetic Analysis of the Target Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elkady, G.; Zhu, J.; Peng, Q.; Chen, M.; Guo, A. Isolation and whole protein characterization of species A and B bovine rotaviruses from Chinese calves. Infect. Genet. Evol. 2021, 89, 104715. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Otto, P.H.; Ciarlet, M.; Desselberger, U.; Ranst, M.V.; Johne, R. VP6-sequence-based cutoff values as a criterion for rotavirus species demarcation. Arch. Virol. 2012, 157, 1177–1182. [Google Scholar] [CrossRef]
- Estes, M.K.; Greenberg, H.B. Rotaviruses. In Fields Virology; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2013; pp. 1347–1401. [Google Scholar]
- Matthijnssens, J.M.; Ciarlet, S.M.; Mcdonald, H.; Attoui, K.; Bányai, J.R.; Brister, J.; Buesa, M.D.; Esona, M.K.; Estes, J.R. Gentsch Uniformity of Rotavirus Strain Nomenclature Proposed by the Rotavirus Classification Working Group (RCWG). Arch. Virol. 2011, 156, 1397–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snodgrass, D.R.; Terzolo, H.R.; Sherwood, D.; Campbell, I.; Menzies, J.D.; Synge, B.A. Aetiology of diarrhoea in young calves. Vet. Rec. 1986, 119, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Windeyer, M.C.; Leslie, K.E.; Godden, S.M.; Hodgins, D.C.; Lissemore, K.D.; Leblanc, S.J. Factors associated with morbidity, mortality, and growth of dairy heifer calves up to 3 months of age. Prev. Vet. Med. 2014, 113, 231–240. [Google Scholar] [CrossRef]
- Karayel, I.; Marton, S.; Coskun, N.; Bányai, K.; Alkan, F. Putative vaccine breakthrough event associated with heterotypic rotavirus infection in newborn calves, Turkey, 2015. Vet. Microbiol. 2017, 201, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Alkan, F.; Ozkul, A.; Oguzoglu, T.C.; Timurkan, M.O.; Caliskan, E.; Martella, V.; Burgu, I. Distribution of G (VP7) and P (VP4) genotypes of group A bovine rotaviruses from Turkish calves with diarrhea, 1997–2008. Vet. Microbiol. 2010, 141, 231–237. [Google Scholar] [CrossRef]
- Swiatek, D.L.; Palombo, E.A.; Lee, A.; Coventry, M.J.; Britz, M.L.; Kirkwood, C.D. Detection and analysis of bovine rotavirus strains circulating in Australian calves during 2004 and 2005. Vet. Microbiol. 2010, 140, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Badaracco, A.; Garaicoechea, L.; Matthijnssens, J.; Uriarte, E.L.; Odeón, A.; Bilbao, G.; Fernandez, F.; Parra, G.I.; Parre, O.V. Phylogenetic analyses of typical bovine rotavirus genotypes G6, G10, P[5] and P[11] circulating in Argentinean beef and dairy herds. Infect. Genet. Evol. 2013, 18, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, T.S.; Lorenzetti, E.; Alfieri, A.F.; Alfieri, A.A. Phylogenetic analysis of a G6P[5] bovine rotavirus strain isolated in a neonatal diarrhea outbreak in a beef cattle herd vaccinated with G6P[1] and G10P[11] genotypes. Arch. Virol. 2015, 160, 447–451. [Google Scholar] [CrossRef]
- Cortese, V.S. Neonatal immunology. Vet. Clin. N. Am. Food Anim. Pract. 2009, 25, 221–227. [Google Scholar] [CrossRef]
- Bartels, C.; Holzhauer, M.; Jorritsma, R.; Swart, W.; Lam, T. Prevalence, prediction and risk factors of enteropathogens in normal and non-normal faeces of young Dutch dairy calves. Prev. Vet. Med. 2010, 93, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Meganck, V.; Hoflack, G.; Piepers, S.; Opsomer, G. Evaluation of a protocol to reduce the incidence of neonatal calf diarrhoea on dairy herds. Prev. Vet. Med. 2015, 118, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Bilcke, J.; Ciarlet, M.; Martella, V.; Bányai, K.; Rahman, M.; Zeller, M.; Beutels, P.; Damme, P.V.; Ranst, M.V. Rotavirus disease and vaccination: Impact on genotype diversity. Future Microbiol. 2009, 4, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Bányai, K.; Matthijnssens, J.; Buonavoglia, C.; Ciarlet, M. Zoonotic aspects of rotaviruses. Vet. Microbiol. 2010, 140, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Badaracco, A.; Garaicoechea, L.; Rodríguez, D.; Uriarte, E.L.; Odeón, A.; Bilbao, G.; Galarza, R.; Abdala, A.; Fernandez, F.; Parre, O.V. Bovine rotavirus strains circulating in beef and dairy herds in Argentina from 2004 to 2010. Vet. Microbiol. 2012, 158, 394–399. [Google Scholar] [CrossRef]
- Usonis, V.; Ivaskeviciene, I.; Desselberger, U.; Rodrigo, C. The unpredictable diversity of co-circulating rotavirus types in Europe and the possible impact of universal mass vaccination programmes on rotavirus genotype incidence. Vaccine 2012, 30, 4596–4605. [Google Scholar] [CrossRef]
- Fritzen, J.; Lorenzetti, E.; Oliveira, M.V.; Bon, V.R.; Ayres, H.; Alfieri, A.F.; Alfieri, A.A. Cross-sectional study of the G and P genotypes of rotavirus A field strains circulating in regularly vaccinated dairy cattle herds. Trop. Anim. Health Prod. 2019, 51, 887–892. [Google Scholar] [CrossRef]
- Matthijnssens, J.; Ciarlet, M.; Heiman, E.; Arijs, I.; Delbeke, T.; McDonald, S.M.; Palombo, E.A.; Iturriza-Gomara, M.; Maes, P.; Patton, J.T.; et al. Full Genome-Based Classification of Rotaviruses Reveals a Common Origin between Human Wa-Like and Porcine Rotavirus Strains and Human DS-1-Like and Bovine Rotavirus Strains. J. Virol. 2008, 82, 3204–3219. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Banik, S.; Faruque, A.; Taniguchi, K.; Sack, D.A.; Van Ranst, M.; Azim, T. Detection and characterization of human group C rotaviruses in Bangladesh. J. Clin. Microbiol. 2005, 43, 4460–4465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, H.A.; Parwani, A.V.; Rosen, B.I.; Lucchelli, A.; Saif, L.J. Detection of rotavirus serotypes G1, G2, G3, and G11 in feces of diarrheic calves by using polymerase chain reaction-derived cDNA probes. J. Clin. Microbiol. 1993, 31, 2491–2496. [Google Scholar] [CrossRef] [Green Version]
- Garaicoechea, L.; Bok, K.; Jones, L.R.; Combessies, G.; Odeón, A.; Fernandez, F.; Parre, O.V. Molecular characterization of bovine rotavirus circulating in beef and dairy herds in Argentina during a 10-year period (1994–2003). Vet. Microbiol. 2006, 118, 1–11. [Google Scholar] [CrossRef]
- Park, S.H.; Saif, L.J.; Jeong, C.; Lim, G.K.; Park, S.I.; Kim, H.H.; Park, S.J.; Kim, Y.J.; Jeong, J.H.; Kang, M.I. Molecular characterization of novel G5 bovine rotavirus strains. J. Clin. Microbiol. 2006, 44, 4101–4112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, P.J.; Cullinane, A.; Martella, V.; O’Shea, H. Molecular characterization of equine rotavirus in Ireland. J. Clin. Microbiol. 2008, 118, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Malik, Y.S.; Sharma, K.; Vaid, N.; Chakravarti, S.; Chandrashekar, K.M.; Basera, S.S.; Singh, R.; Prasad, G.; Gulati, B.R.; Bhilegaonkar, K.N.; et al. Frequency of group A rotavirus with mixed G and P genotypes in bovines: Predominance of G3 genotype and its emergence in combination with G8/G10 types. J. Vet. Sci. 2012, 13, 271–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, N.; Yuanwei, W.; Bin, Z.; Hua, Y.; Cheng, T. High prevalence and genomic characteristics of G6P[1] Bovine Rotavirus A in yak in China. J. Gen. Virol. 2020, 101, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, R.; Singh, K.P.; Singh, V.; Malik, Y.; Kamdi, B.; Singh, R.; Kashyap, G. Immunohistochemical and molecular detection of natural cases of bovine rotavirus and coronavirus infection causing enteritis in dairy calves. Elsevier Public Health Emerg. Collect. 2020, 138, 103814. [Google Scholar] [CrossRef]
- Bertoni, E.; Aduriz, M.; Bok, M.; Vega, C.; Saif, L.; Aguirre, D.; Cimino, R.O.; Miño, S.; Parreño, V. First report of group A rotavirus and bovine coronavirus associated with neonatal calf diarrhea in the northwest of Argentina. Trop. Anim. Health Prod. 2020, 52, 2761–2768. [Google Scholar] [CrossRef]
- Wei, X.; Wang, W.; Dong, Z.; Cheng, F.; Zhang, J. Detection of Infectious Agents Causing Neonatal Calf Diarrhea on Two Large Dairy Farms in Yangxin County, Shandong Province, China. Front. Vet. Sci. 2021, 7, 589126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Su, D.; Meng, X.; Liang, R.; Feng, Y. Cryptosporidiosis outbreak caused by Cryptosporidium parvum subtype IIdA20G1 in neonatal calves. Transbound Emerg. Dis. 2021, 1–8. [Google Scholar] [CrossRef]
- Gillhuber, J.; Rügamer, D.; Pfister, K.; Scheuerle, M. Giardiosis and other enteropathogenic infections: A study on diarrhoeic calves in Southern Germany. BMC Res. Notes 2014, 7, 112. [Google Scholar] [CrossRef] [Green Version]
- Uhde, F.L.; Kaufmann, T.; Sager, H.; Albini, S.; Zanoni, R.; Schelling, E.; Meylan, M. Prevalence of four enteropathogens in the faeces of young diarrhoeic dairy calves in Switzerland. Vet. Rec. J. Br. Vet. Assoc. 2008, 163, 362–366. [Google Scholar]
- Dall Agnol, A.M.; Lorenzetti, E.; Leme, R.A.; Ladeia, W.A.; Mainardi, R.M.; Bernardi, A.; Headley, S.A.; Freire, R.L.; Pereira, U.P.; Alfieri, A.F. Severe outbreak of bovine neonatal diarrhea in a dairy calf rearing unit with multifactorial etiology. Brazilian journal of microbiology. Braz. Soc. Microbiol. 2021, 52, 2547–2553. [Google Scholar] [CrossRef] [PubMed]
- Brunauer, M.; Roch, F.F.; Conrady, B. Prevalence of Worldwide Neonatal Calf Diarrhoea Caused by Bovine Rotavirus in Combination with Bovine Coronavirus, Escherichia coli K99 and Cryptosporidium spp.: A Meta-Analysis. Animals 2021, 11, 1014. [Google Scholar] [CrossRef]
- Ciarlet, M.; Hyser, J.M.; Estes, M.K. Sequence analysis of the VP4, VP6, VP7, and NSP4 gene products of the bovine rotavirus WC3. Virus Genes 2002, 24, 107–118. [Google Scholar] [CrossRef]
- Favacho, A.R.M.; Kurtenbach, E.; Sardi, S.I.; Gouvea, V.S. Cloning, expression, and purification of recombinant bovine rotavirus hemagglutinin, VP8*, in Escherichia coli. Protein Expr. Purif. 2006, 46, 196–203. [Google Scholar] [CrossRef]
- Fernandez, F.M.; Conner, M.E.; Hodgins, D.C.; Parwani, A.V.; Nielsen, P.R.; Crawford, S.E.; Estes, M.K.; Saif, L.J. Passive immunity to bovine rotavirus in newborn calves fed colostrum supplements from cows immunized with recombinant SA11 rotavirus core-like particle (CLP) or virus-like particle (VLP) vaccines. Vaccine 1998, 16, 507–516. [Google Scholar] [CrossRef]
- Greenberg, H.B.; Valdesuso, J.O.S.E.; van Wyke, K.A.T.H.L.E.E.N.; Midthun, K.A.R.E.N.; Walsh, M.; McAuliffe, V.; Wyatt, R.G.; Kalica, A.R.; Flores, J.; Hoshino, Y. Production and preliminary characterization of monoclonal antibodies directed at two surface proteins of rhesus rotavirus. J. Virol. 1983, 47, 267–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aminu, M.; Page, N.A.; Ahmad, A.A.; Umoh, J.U.; Dewar, J.; Steele, A.D. Diversity of Rotavirus VP7 and VP4 Genotypes in Northwestern Nigeria. J. Infect. Dis. 2010, 202 (Suppl. S1), S198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalilian, S.; Teimoori, A.; Makvandi, M. In Silico Characterization of Epitopes from Human Rotavirus VP7 Genotype G9 Design for Vaccine Development. Iran. J. Allergy Asthma Immunol. 2019, 18, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Midgley, S.E.; Bányai, K.; Buesa, J.; Halaihel, N.; Bttiger, B. Diversity and zoonotic potential of rotaviruses in swine and cattle across Europe. Vet. Microbiol. 2011, 156, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, A.F.; Alfieri, A.A.; Barreiros, M.; Leite, J.; Richtzenhain, L.J. G and P genotypes of group A rotavirus strains circulating in calves in Brazil, 1996–1999. Vet. Microbiol. 2004, 99, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Reidy, N.; Lennon, G.; Fanning, S.; Power, E.; O’Shea, H. Molecular characterisation and analysis of bovine rotavirus strains circulating in Ireland 2002–2004. Vet. Microbiol. 2006, 117, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Xiaoying, L.; Nan, Y.; Hua, Y.; Yuanwei, W.; Bin, Z.; Cheng, T. Detection and molecular characteristics of bovine rotavirus A in dairy calves in China. J. Vet. Sci. 2021, 22, e69. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Area | Number of Farms | Number of Samples | Health Management | Age of the Cattles |
|---|---|---|---|---|
| Heilongjiang | 3 | 81 | Diarrhea | 3–8 weeks |
| Neimenggu | 5 | 78 | Diarrhea | 3–8 weeks |
| Jilin | 4 | 74 | Diarrhea | 3–8 weeks |
| Name | Primers (5′–3′) | Product Size (bp) |
|---|---|---|
| BRV-1 | CGATAATGTATGTATGGACG | 211 |
| BRV-2 | TGCTGAATAAGGGAAAATG | |
| BRV-VP4-F | GGCTTTAAAATGGCTTCACTCAT | 2362 |
| BRV-VP4-R | GGTCACATCCTCTGTCAGTTGCT | |
| BRV-VP6-F | GGCTTTTAAACGAAGTCTTCA | 1356 |
| BRV-VP6-R | GGTCACATCCTCTCACTACG | |
| BRV-VP7-F | GGCTTTAAAAGCGAGAATTTCCGTT | 1062 |
| BRV-VP7-R | GGTCACATCATACAACTCTAACT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, X.; Wu, W.; Teng, F.; Yan, Y.; Li, G.; Wang, L.; Wang, X.; Wang, R.; Zhou, H.; Jiang, Y.; et al. Isolation and Characterization of Bovine RVA from Northeast China, 2017–2020. Life 2021, 11, 1389. https://doi.org/10.3390/life11121389
Cheng X, Wu W, Teng F, Yan Y, Li G, Wang L, Wang X, Wang R, Zhou H, Jiang Y, et al. Isolation and Characterization of Bovine RVA from Northeast China, 2017–2020. Life. 2021; 11(12):1389. https://doi.org/10.3390/life11121389
Chicago/Turabian StyleCheng, Xi, Wei Wu, Fei Teng, Yue Yan, Guiwei Li, Li Wang, Xiaona Wang, Ruichong Wang, Han Zhou, Yanping Jiang, and et al. 2021. "Isolation and Characterization of Bovine RVA from Northeast China, 2017–2020" Life 11, no. 12: 1389. https://doi.org/10.3390/life11121389