
A Multiallelic Molecular Beacon-Based Real-Time RT-PCR Assay for the Detection of SARS-CoV-2

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
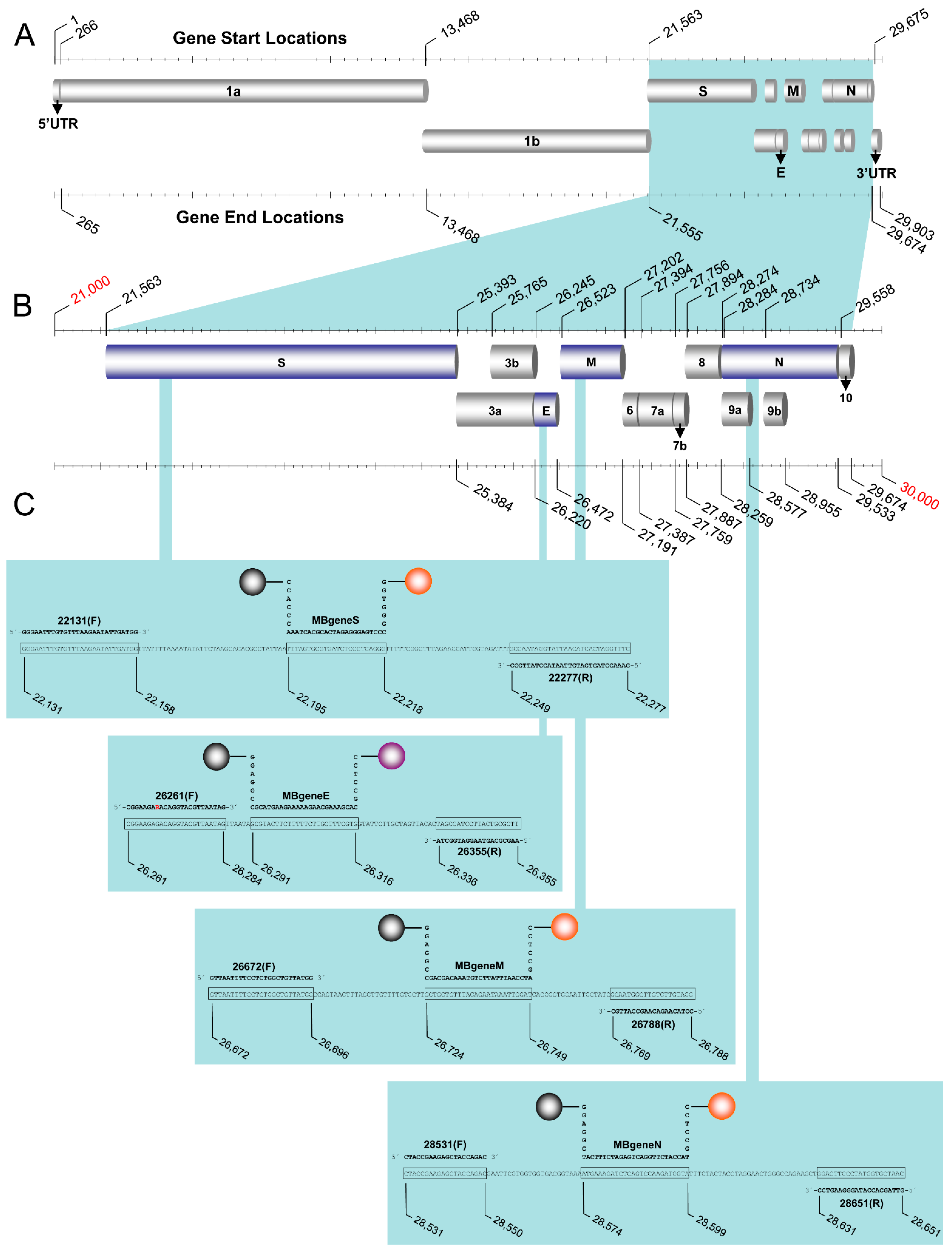
2.1. Molecular Beacon and Primer Design
2.2. Internal Positive Control
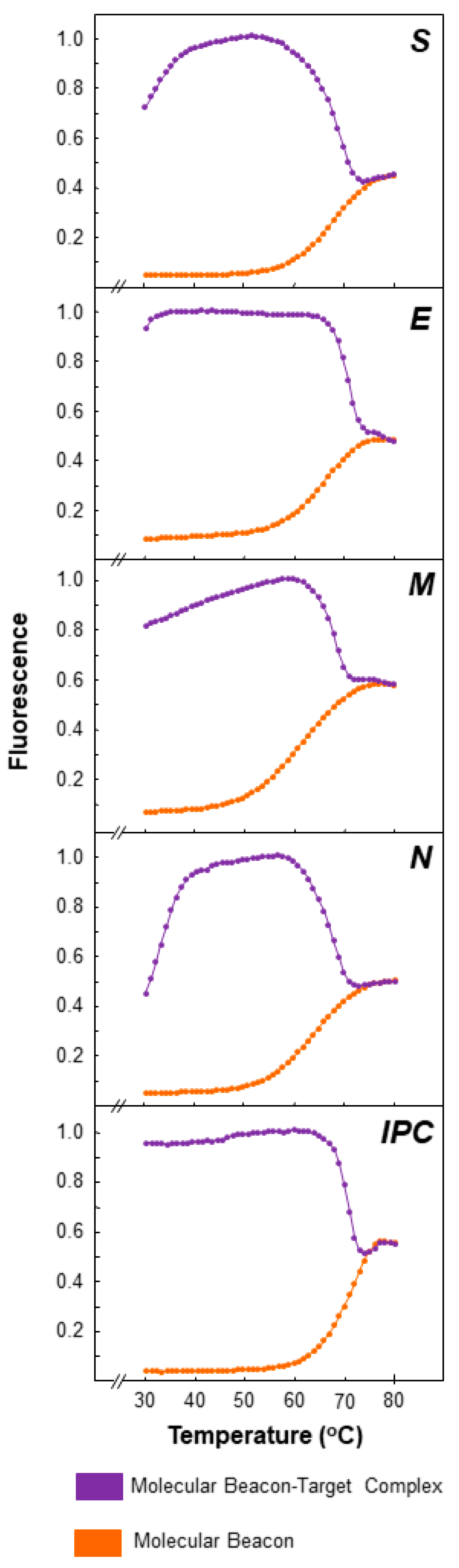
2.3. Thermal Profiles of Molecular Beacons
2.4. RNA Transcripts
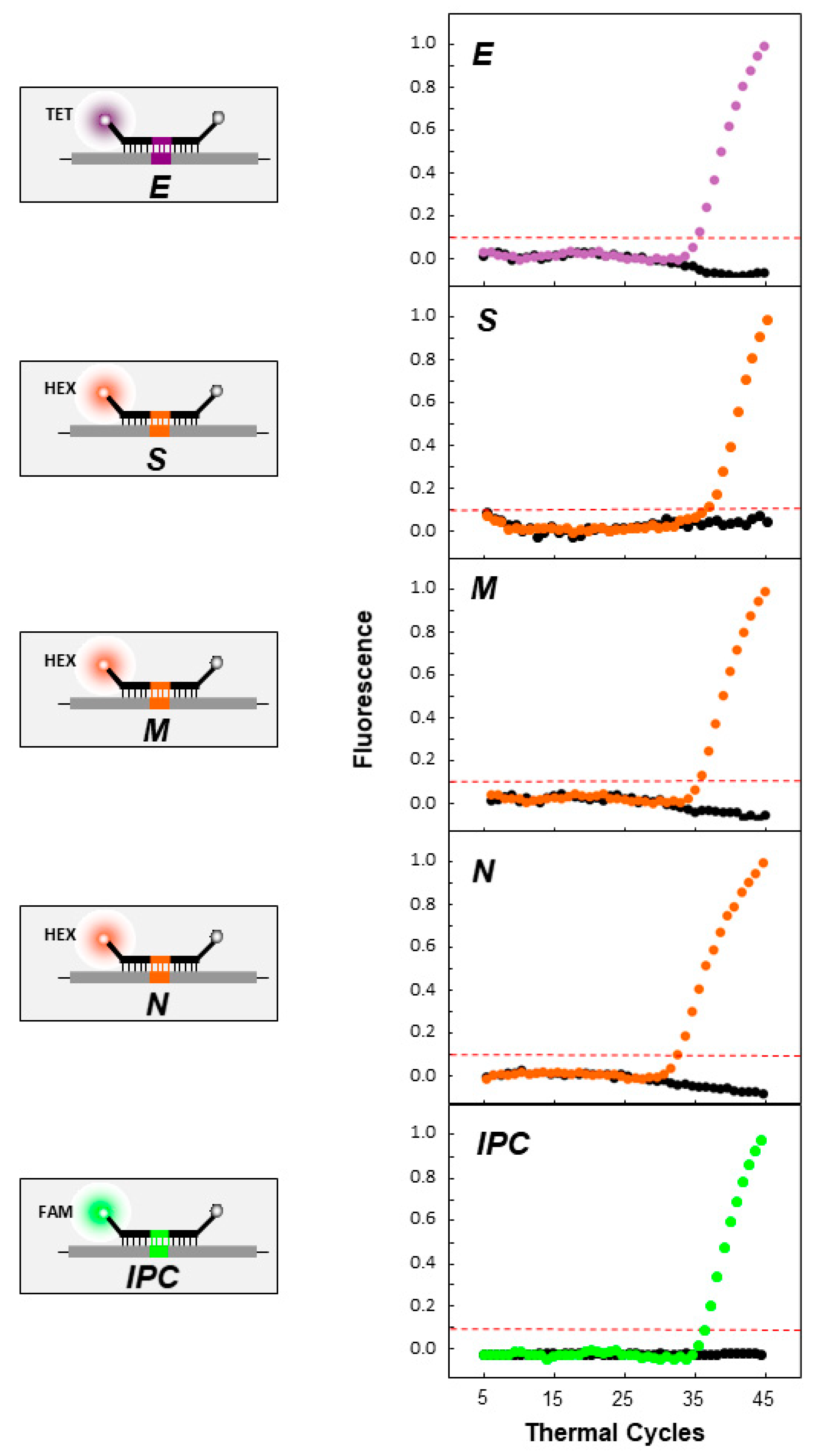
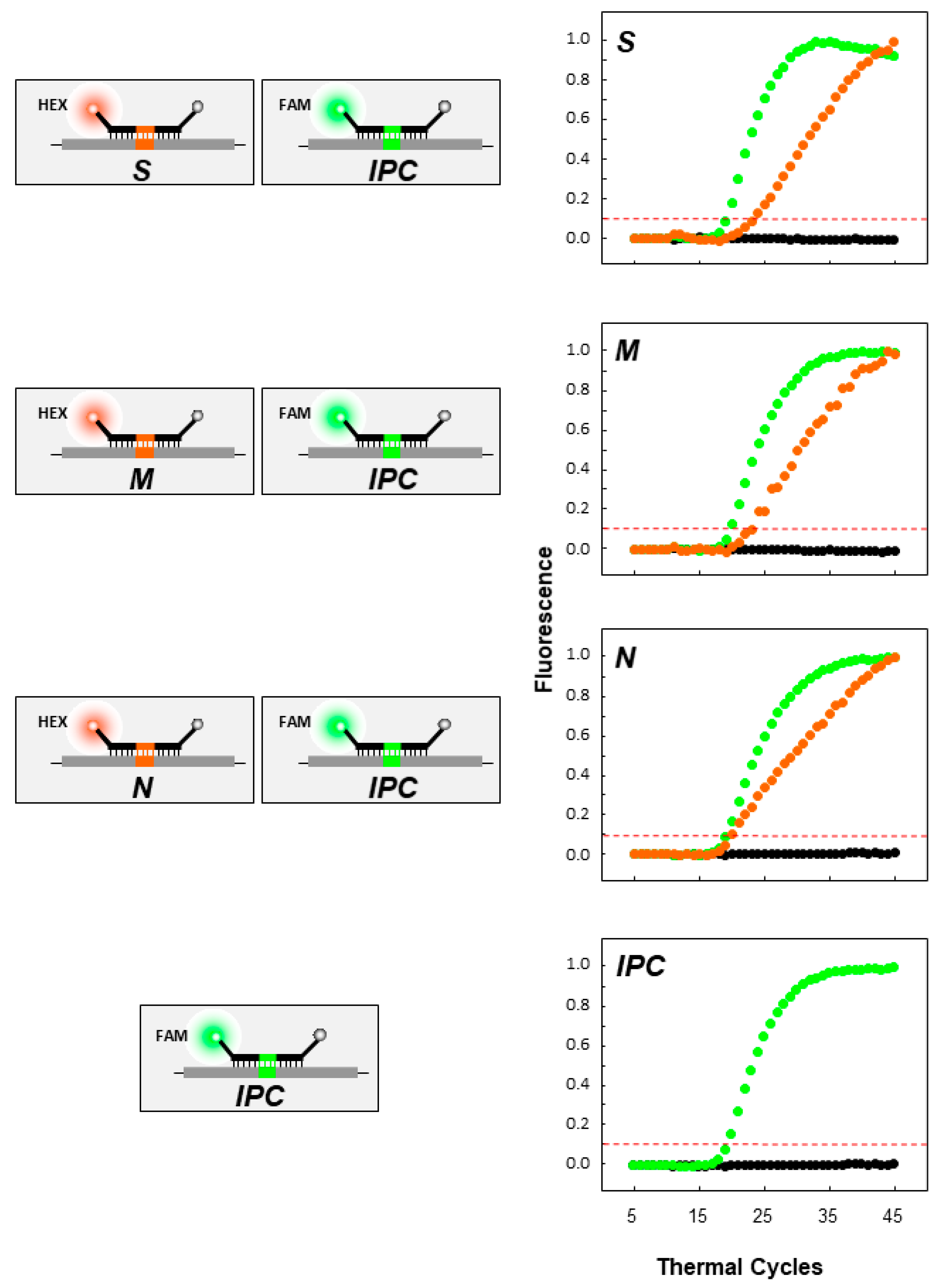
2.5. Real-Time RT-PCR: Uniplex and Duplex Reactions
2.6. Validation and Clinical Samples
3. Results
3.1. Molecular Beacon Thermal Denaturation Profiles
3.2. Real-Time RT-PCR Validation and Testing
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fauci, A.S.; Morens, D.M. The Perpetual Challenge of Infectious Diseases. N. Engl. J. Med. 2012. [Google Scholar] [CrossRef] [Green Version]
- Devaux, C.A. Emerging and re-emerging viruses: A global challenge illustrated by Chikungunya virus outbreaks. World J. Virol. 2012, 1, 11. [Google Scholar] [CrossRef] [PubMed]
- Rabi, F.A.; Al Zoubi, M.S.; Al-Nasser, A.D.; Kasasbeh, G.A.; Salameh, D.M. Sars-cov-2 and coronavirus disease 2019: What we know so far. Pathogens 2020, 9, 231. [Google Scholar] [CrossRef] [PubMed]
- Peiris, J.S.M.; Yuen, K.Y.; Osterhaus, A.D.M.E.; Stöhr, K. The Severe Acute Respiratory Syndrome. N. Engl. J. Med. 2003, 349, 2431–2441. [Google Scholar] [CrossRef] [Green Version]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012. [Google Scholar] [CrossRef] [PubMed]
- Yuen, K.S.; Ye, Z.W.; Fung, S.Y.; Chan, C.P.; Jin, D.Y. SARS-CoV-2 and COVID-19: The most important research questions. Cell Biosci. 2020, 10, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, Y.; Chen, Y.; Qin, Q. Unique epidemiological and clinical features of the emerging 2019 novel coronavirus pneumonia (COVID-19) implicate special control measures. J. Med. Virol. 2020, 92, 568–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Costa, V.G.; Moreli, M.L.; Saivish, M.V. The emergence of SARS, MERS and novel SARS-2 coronaviruses in the 21st century. Arch. Virol. 2020, 165, 1517–1526. [Google Scholar] [CrossRef]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Kim, D.; Lee, J.Y.; Yang, J.S.; Kim, J.W.; Kim, V.N.; Chang, H. The Architecture of SARS-CoV-2 Transcriptome. Cell 2020, 181, 914–921. [Google Scholar] [CrossRef] [PubMed]
- Bojkova, D.; Klann, K.; Koch, B.; Widera, M.; Krause, D.; Ciesek, S.; Cinatl, J.; Münch, C. Proteomics of SARS-CoV-2-infected host cells reveals therapy targets. Nature 2020, 583, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Harb, J.G.; Noureldine, H.A.; Chedid, G.; Eldine, M.N.; Abdallah, D.A.; Chedid, N.F.; Nour-Eldine, W. SARS, MERS and COVID-19: Clinical manifestations and organ-system complications: A mini review. Pathog. Dis. 2020, 78, ftaa033. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, J.S.; Smith, D.W. COVID-19: A novel zoonotic disease caused by a coronavirus from China: What we know and what we don’t. Microbiol. Aust. 2020, 41, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Helms, J.; Kremer, S.; Merdji, H.; Clere-Jehl, R.; Schenck, M.; Kummerlen, C.; Collange, O.; Boulay, C.; Fafi-Kremer, S.; Ohana, M.; et al. Neurologic Features in Severe SARS-CoV-2 Infection. N. Engl. J. Med. 2020, 383, 2268–2270. [Google Scholar] [CrossRef]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England. Science (80-) 2021, 372, eabg3055. [Google Scholar] [CrossRef]
- Hu, E. COVID-19 Testing: Challenges, Limitations and Suggestions for Improvement. Preprints 2020, 2020040155. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Y.J.; Wei, C.L.; Ee, L.A.; Vega, V.B.; Thoreau, H.; Yun, S.T.S.; Chia, J.M.; Ng, P.; Chiu, K.P.; Lim, L.; et al. Comparative full-length genome sequence analysis of 14 SARS coronavirus isolates and common mutations associated with putative origins of infection. Lancet 2003, 361, 1779–1785. [Google Scholar] [CrossRef] [Green Version]
- Zella, D.; Giovanetti, M.; Cella, E.; Borsetti, A.; Ciotti, M.; Ceccarelli, G.; D’Ettorre, G.; Pezzuto, A.; Tambone, V.; Campanozzi, L.; et al. The importance of genomic analysis in cracking the coronavirus pandemic. Expert Rev. Mol. Diagn. 2021, 21, 1–16. [Google Scholar] [CrossRef]
- Charkiewicz, R.; Nikliński, J.; Biecek, P.; Kiśluk, J.; Pancewicz, S.; Moniuszko-Malinowska, A.M.; Flisiak, R.; Krętowski, A.J.; Dzięcioł, J.; Moniuszko, M.; et al. The first SARS-CoV-2 genetic variants of concern (VOC) in Poland: The concept of a comprehensive approach to monitoring and surveillance of emerging variants. Adv. Med. Sci. 2021, 66, 237–245. [Google Scholar] [CrossRef]
- Rambaut, A.; Holmes, E.C.; O’Toole, Á.; Hill, V.; McCrone, J.T.; Ruis, C.; du Plessis, L.; Pybus, O.G. A dynamic nomenclature proposal for SARS-CoV-2 lineages to assist genomic epidemiology. Nat. Microbiol. 2020, 5, 1403–1407. [Google Scholar] [CrossRef] [PubMed]
- Bal, A.; Destras, G.; Gaymard, A.; Stefic, K.; Marlet, J.; Eymieux, S.; Regue, H.; Semanas, Q.; D’Aubarede, C.; Billaud, G.; et al. Two-step strategy for the identification of SARS-CoV-2 variant of concern 202012/01 and other variants with spike deletion H69–V70, France, August to December 2020. Eurosurveillance 2021, 23, 2100008. [Google Scholar] [CrossRef]
- Leelawong, M.; Mitchell, S.L.; Fowler, R.C.; Hughes, S.; Griffith, M.P.; Marsh, J.W.; Harrison, L.H.; Rakeman, J.L. SARS-CoV-2 N gene mutations impact detection by clinical molecular diagnostics: Reports in two cities in the United States. Diagn. Microbiol. Infect. Dis. 2021, 101, 115468. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Miller, J.A.; Verghese, M.; Sibai, M.; Solis, D.; Mfuh, K.O.; Jiang, B.; Iwai, N.; Mar, M.; Huang, C.; et al. Multiplex SARS-CoV-2 Genotyping RT-PCR for Population-Level Variant Screening and Epidemiologic Surveillance. J. Clin. Microbiol. 2021, 59, e00859-12. [Google Scholar] [CrossRef] [PubMed]
- Tahan, S.; Parikh, B.A.; Droit, L.; Wallace, M.A.; Burnham, C.-A.D.; Wang, D. SARS-CoV-2 E Gene Variant Alters Analytical Sensitivity Characteristics of Viral Detection Using a Commercial Reverse Transcription-PCR Assay. J. Clin. Microbiol. 2021, 59, e00075-21. [Google Scholar] [CrossRef] [PubMed]
- Hadjinicolaou, A.V.; Farcas, G.A.; Demetriou, V.L.; Mazzulli, T.; Poutanen, S.M.; Willey, B.M.; Low, D.E.; Butany, J.; Asa, S.L.; Kain, K.C.; et al. Development of a molecular-beacon-based multi-allelic real-time RT-PCR assay for the detection of human coronavirus causing severe acute respiratory syndrome (SARS-CoV): A general methodology for detecting rapidly mutating viruses. Arch. Virol. 2011, 156, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple Sequence Alignment Using ClustalW and ClustalX. Curr. Protoc. Bioinforma. 2003, 00, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Kramer, F.R.; Marras, S.A.E.; Tyagi, S. Inventing molecular beacons. In The PCR Revolution: Basic Technologies and Applications; Cambridge University Press: Cambridge, UK, 2009; ISBN 9780511818974. [Google Scholar]
- Vet, J.A.M.; Van der Rijt, B.J.M.; Blom, H.J. Molecular beacons: Colorful analysis of nucleic acids. Expert Rev. Mol. Diagn. 2002, 2, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Hu, Y.; Song, Z.G.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. Complete genome characterisation of a novel coronavirus associated with severe human respiratory disease in Wuhan, China. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W6. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.A.; Sacco, O.; Mancino, E.; Cristiani, L.; Midulla, F. Differences and similarities between SARS-CoV and SARS-CoV-2: Spike receptor-binding domain recognition and host cell infection with support of cellular serine proteases. Infection 2020, 48, 665–669. [Google Scholar] [CrossRef]
- Rahman, M.S.; Hoque, M.N.; Islam, M.R.; Islam, I.; Mishu, I.D.; Rahaman, M.M.; Sultana, M.; Hossain, M.A. Mutational insights into the envelope protein of SARS-CoV-2. Gene Rep. 2021, 22, 100997. [Google Scholar] [CrossRef]
- Bianchi, M.; Benvenuto, D.; Giovanetti, M.; Angeletti, S.; Ciccozzi, M.; Pascarella, S. Sars-CoV-2 Envelope and Membrane Proteins: Structural Differences Linked to Virus Characteristics? Biomed Res. Int. 2020, 2020, 6. [Google Scholar] [CrossRef]
- Chazal, N. Coronavirus, the King Who Wanted More Than a Crown: From Common to the Highly Pathogenic SARS-CoV-2, Is the Key in the Accessory Genes? Front. Microbiol. 2021, 12, 1970. [Google Scholar] [CrossRef] [PubMed]
- Ayass bioscience VOC – Variants of Concern Sequencing. Available online: https://ayassbioscience.com/voc/ (accessed on 28 July 2021).
- Chen, T.; Fei, C.-Y.; Chen, Y.-P.; Sargsyan, K.; Liang, J.-J.; Liao, C.-C.; Lin, Y.-L.; Chang, C.-P.; Yuan, H.S.; Lim, C. Correction to “Synergistic Inhibition of SARS-CoV-2 Replication Using Disulfiram/Ebselen and Remdesivir”. ACS Pharmacol. Transl. Sci. 2021, 12, 1246. [Google Scholar] [CrossRef] [PubMed]
- Bier, C.; Edelmann, A.; Theil, K.; Schwarzer, R.; Deichner, M.; Gessner, A.; Hiergeist, A.; Rentschler, U.; Gohl, P.; Kuchta, A.; et al. Multi-site Evaluation of SARS-CoV-2 Spike Mutation Detection Using a Multiplex Real-time RT-PCR Assay. medRxiv 2021. [Google Scholar] [CrossRef]
- Etievant, S.; Bal, A.; Escuret, V.; Brengel-Pesce, K.; Bouscambert, M.; Cheynet, V.; Generenaz, L.; Oriol, G.; Destras, G.; Billaud, G.; et al. Performance Assessment of SARS-CoV-2 PCR Assays Developed by WHO Referral Laboratories. J. Clin. Med. 2020, 9, 1871. [Google Scholar] [CrossRef] [PubMed]
- Chrysostomou, A.C.; Vrancken, B.; Koumbaris, G.; Themistokleous, G.; Aristokleous, A.; Masia, C.; Eleftheriou, C.; Iοannou, C.; Stylianou, D.C.; Ioannides, M.; et al. A Comprehensive Molecular Epidemiological Analysis of SARS-CoV-2 Infection in Cyprus from April 2020 to January 2021: Evidence of a Highly Polyphyletic and Evolving Epidemic. Viruses 2021, 13, 1098. [Google Scholar] [CrossRef]
- GISAID GISAID Initiative. Available online: https://www.gisaid.org/ (accessed on 24 February 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation 1 | TargetGene | Sequence | Position 2 | Amplicon Length (nts) 3 | Gene Accession Number 4 |
|---|---|---|---|---|---|
| PCR Primers | |||||
| 22131 (F) | S | GGGAATTTGTGTTTAAGAATATTGATGG | 22,131–22,158 | MN908947.3 | |
| 22277 (R) | S | GAAACCTAGTGATGTTAATACCTATTGGC | 22,249–22,277 | MN908947.3 | |
| 26261 (F) | E | CGGAAGARACAGGTACGTTAATAG | 26,261–26,284 | MN908947.3 | |
| 26355 (R) | E | AAGCGCAGTAAGGATGGCTA | 26,336–26,355 | MN908947.3 | |
| 26672 (F) | M | GTTAATTTTCCTCTGGCTGTTATGG | 26,672–26,696 | MN908947.3 | |
| 26788 (R) | M | CCTACAAGACAAGCCATTGC | 26,769–26,788 | MN908947.3 | |
| 28531 (F) | N | CTACCGAAGAGCTACCAGAC | 28,531–28,550 | MN908947.3 | |
| 28651 (R) | N | GTTAGCACCATAGGGAAGTCC | 28,631–28,651 | MN908947.3 | |
| Target Amplicons | |||||
| TgeneS | S | GGGAATTTGTGTTTAAGAATATTGATGGTTATTTTAAAATATATTCTAAGCACA CGCCTATTAATTTAGTGCGTGATCTCCCTCAGGGTTTTTCGGCTTTAGAACCAT GGTAGATTTGCCAATAGGTATTAACATCACTAGGTTTC | 22,131–22,277 | 147 | MN908947.3 |
| TgeneE | E | CGGAAGAGACAGGTACGTTAATAGTTAATAGCGTACTTCTTTTTCTTGCTTTCG TGGTATTCTTGCTAGTTACACTAGCCATCCTTACTGCGCTT | 26,261–26,355 | 95 | MN908947.3 |
| TgeneM | M | GTTAATTTTCCTCTGGCTGTTATGGCCAGTAACTTTAGCTTGTTTTGTGCTTGC TGCTGTTTACAGAATAAATTGGATCACCGGTGGAATTGCTATCGCAATGGCTTG TCTTGTAGG | 26,672–26,788 | 117 | MN908947.3 |
| TgeneN | N | CTACCGAAGAGCTACCAGACGAATTCGTGGTGGTGACGGTAAAATGAAAGATCTGGTGGTGACGGTAAAATGAAAGATCTCAGTCCAAGATGGTATTTCTACTACCTA GGAACTGGGCCAGAAGCTGGACTTCCCTATGGTGCTAAC | 28,531–28,651 | 121 | MN908947.3 |
| TIPC | N/A | GGGAATTTGTGTTTAAGAATATTGATGGTTAGCTGCTGTTTACAGTCCAAGATG GTAGTATTCTTGCTAGTTACACTAGCCATCCTTACTGCGCTT | N/A | 96 | N/A |
| Molecular Beacons5 | |||||
| MBgeneS | S | HEX-GGTGGGCCCTGAGGGAGATCACGCACTAAACCCACC-DABCYL | 22,195–22,218 | MN908947.3 | |
| MBgeneE | E | TET-CCTCCGCACGAAAGCAAGAAAAAGAAGTACGCCGGAGG-DABCYL | 26,291–26,316 | MN908947.3 | |
| MBgeneM | M | HEX-CCTCCGATCCAATTTATTCTGTAAACAGCAGCCGGAGG-DABCYL | 26,724–26,749 | MN908947.3 | |
| MBgeneN | N | HEX-CCTCCGTACCATCTTGGACTGAGATCTTTCATCGGAGG-DABCYL | 28,574–28,599 | MN908947.3 | |
| MBIPC | N/A | FAM-GCCCACGTACCATCTTGGACTGTAAACAGCAGCCGTGGGC-DABCYL | N/A | N/A |
| Sample | ID of Specimen Analyzed | RT-PCR Result 4 | |||
|---|---|---|---|---|---|
| S | E | M | N | ||
| EVAg 1 | |||||
| 1 | Purified RNA of SARS-CoV-2 cell culture supernatant | + | + | + | + |
| WHO 2 | |||||
| 2 | 2020-01 (SARS-CoV-2) | + | + | + | + |
| 3 | 2020-02 (SARS-CoV-2) | + | + | + | + |
| 4 | 2020-03 (NEGATIVE) | − | − | − | − |
| 5 | 2020-04 (OC43) | − | − | − | − |
| 6 | 2020-05 (SARS-CoV-2) | + | + | + | + |
| 7 | Positive control (SARS-CoV-2) | + | + | + | + |
| QCMD 3 | |||||
| 8 | CVOP20S2-01 (SARS-CoV-2) | + | + | + | + |
| 9 | CVOP20S2-02 (NL63) | − | − | − | − |
| 10 | CVOP20S2-03 (SARS-CoV-2) | + | + | + | + |
| 11 | CVOP20S2-04 (OC43) | − | − | − | − |
| 12 | CVOP20S2-05 (NEGATIVE) | − | − | − | − |
| 13 | CVOP20S2-06 (SARS-CoV-2) | + | + | + | + |
| 14 | CVOP20S2-07 (SARS-CoV-2) | + | + | + | + |
| 15 | CVOP20S2-08 (SARS-CoV-2) | + | + | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrysostomou, A.C.; Hezka Rodosthenous, J.; Topcu, C.; Papa, C.; Aristokleous, A.; Stathi, G.; Christodoulou, C.; Eleftheriou, C.; Stylianou, D.C.; Kostrikis, L.G. A Multiallelic Molecular Beacon-Based Real-Time RT-PCR Assay for the Detection of SARS-CoV-2. Life 2021, 11, 1146. https://doi.org/10.3390/life11111146
Chrysostomou AC, Hezka Rodosthenous J, Topcu C, Papa C, Aristokleous A, Stathi G, Christodoulou C, Eleftheriou C, Stylianou DC, Kostrikis LG. A Multiallelic Molecular Beacon-Based Real-Time RT-PCR Assay for the Detection of SARS-CoV-2. Life. 2021; 11(11):1146. https://doi.org/10.3390/life11111146
Chicago/Turabian StyleChrysostomou, Andreas C., Johana Hezka Rodosthenous, Cicek Topcu, Christina Papa, Antonia Aristokleous, Georgia Stathi, Christina Christodoulou, Christina Eleftheriou, Dora C. Stylianou, and Leondios G. Kostrikis. 2021. "A Multiallelic Molecular Beacon-Based Real-Time RT-PCR Assay for the Detection of SARS-CoV-2" Life 11, no. 11: 1146. https://doi.org/10.3390/life11111146