
Compendium of Plant-Specific CRISPR Vectors and Their Technical Advantages

 ,
,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Size of gRNA Is Important for Efficient Cas9 Endonuclease Activity
3. Polycistronic-tRNA/Csy4-gRNA Is a Need for Editing of Multiple Targets with a Single Construct
4. Importance of Promoter Regulating Cas9 and gRNA towards Editing Efficiency
5. CRISPR On/Off Strategy Uses Inducible Promoter
6. Plant Selectable Markers Are Needed
7. Selectable Marker and Plasmid Backbone Affecting Transformation Efficiency
8. Origin of Replication and Its Importance during Molecular Cloning
9. CRISPR/Cas Toolkits for Plants
9.1. Cas/sgRNA Ribonucleoprotein Complexes as Editing Tools
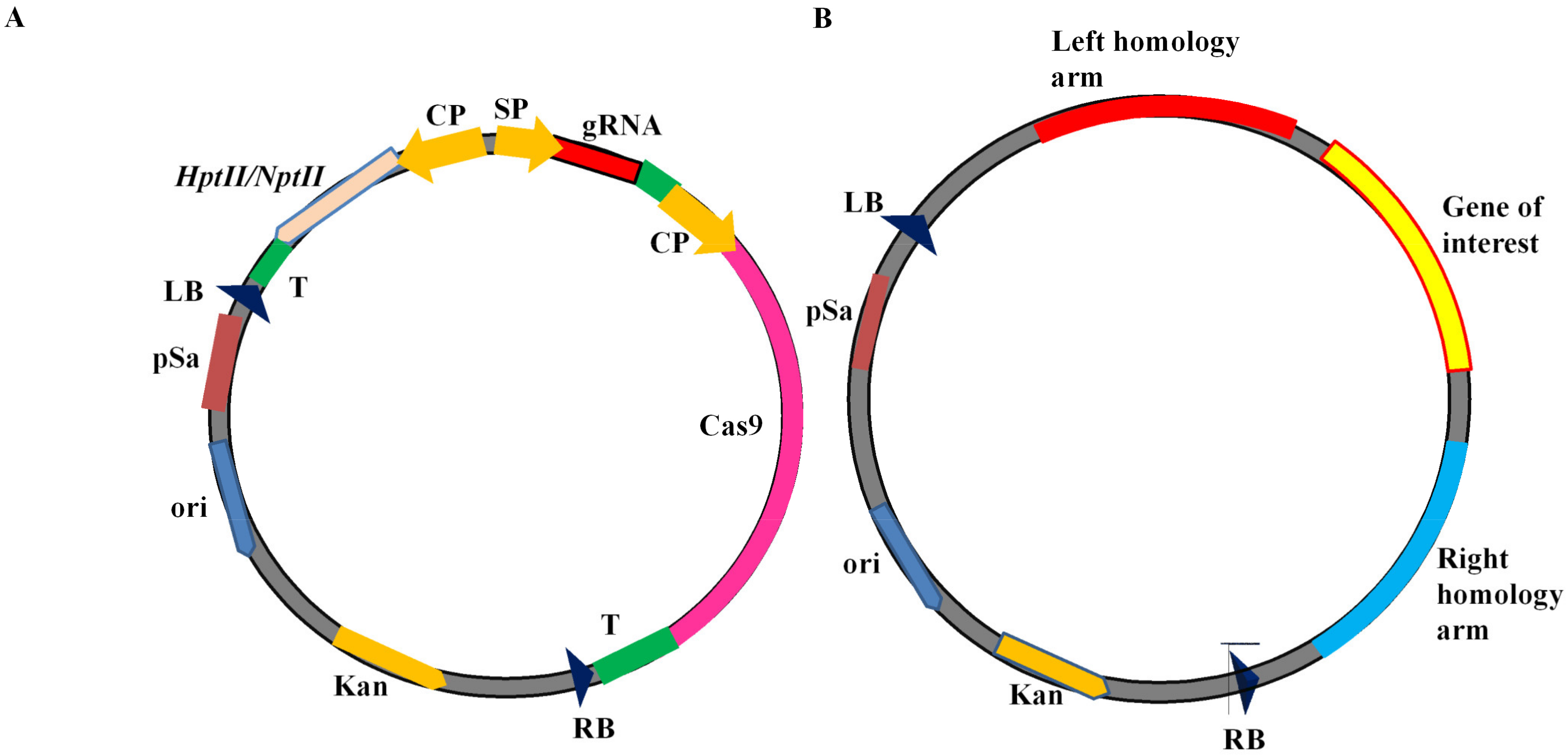
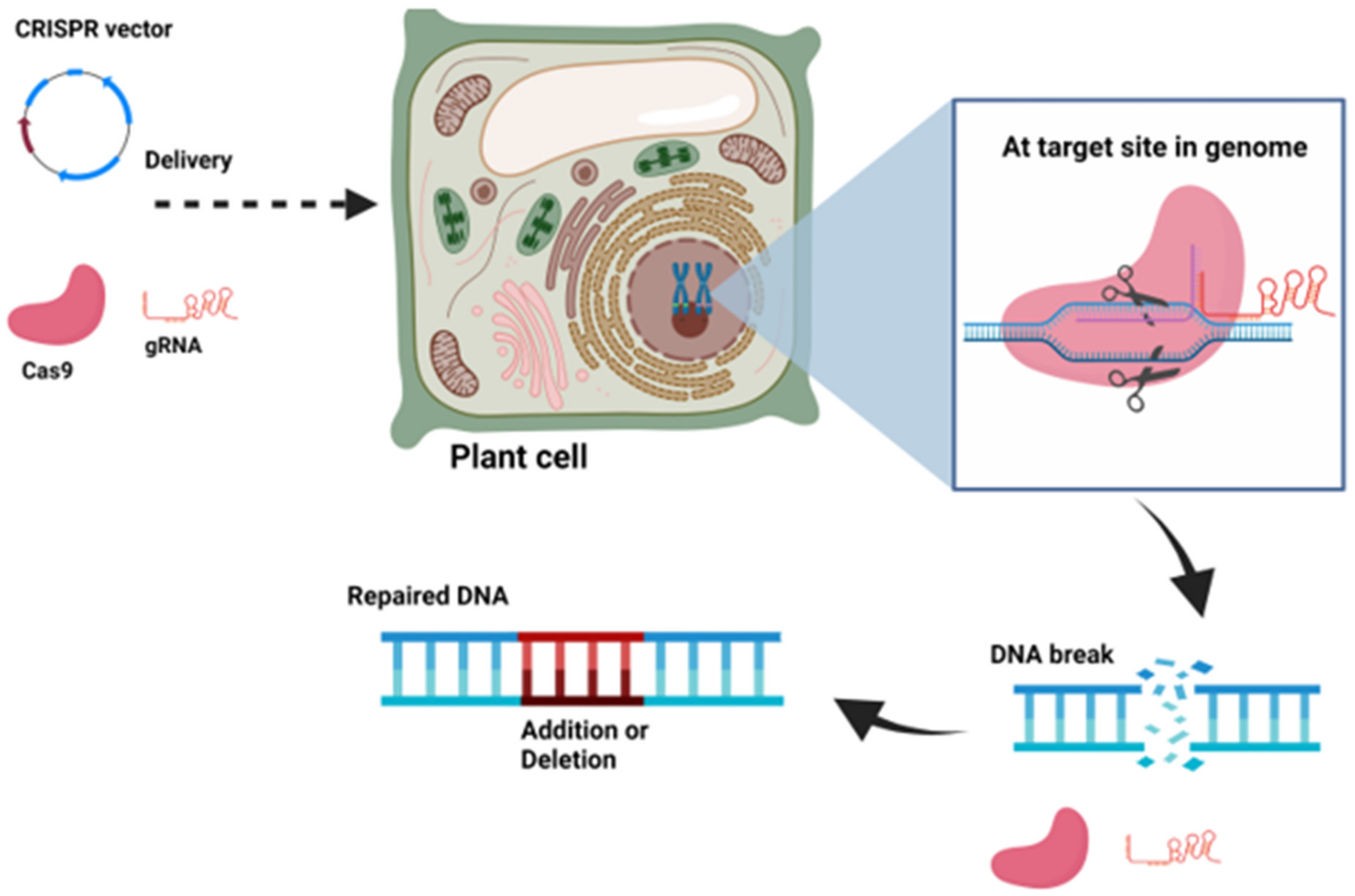
9.2. Knock-Out CRISPR Vectors
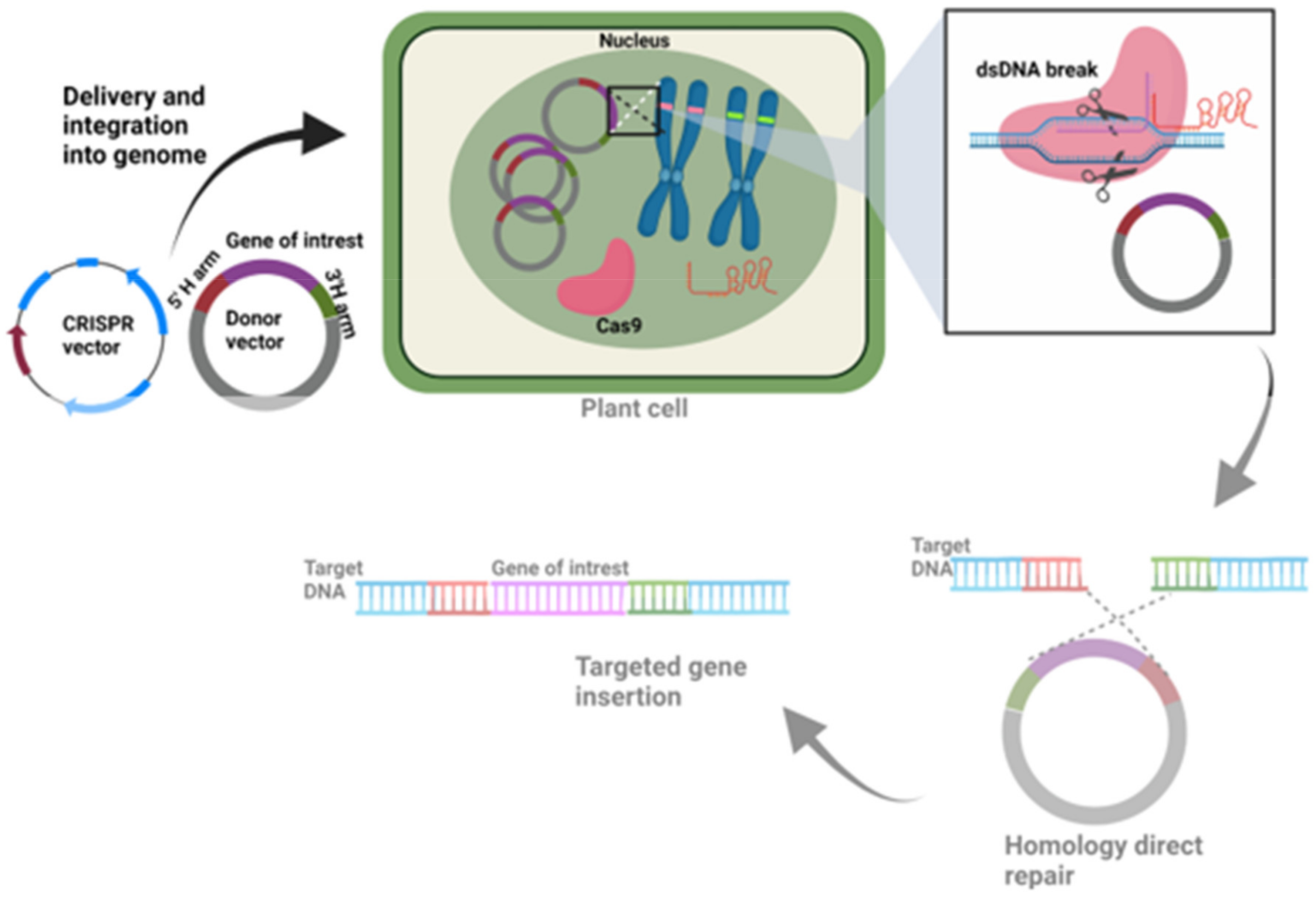
9.3. Knock-In CRISPR Vectors
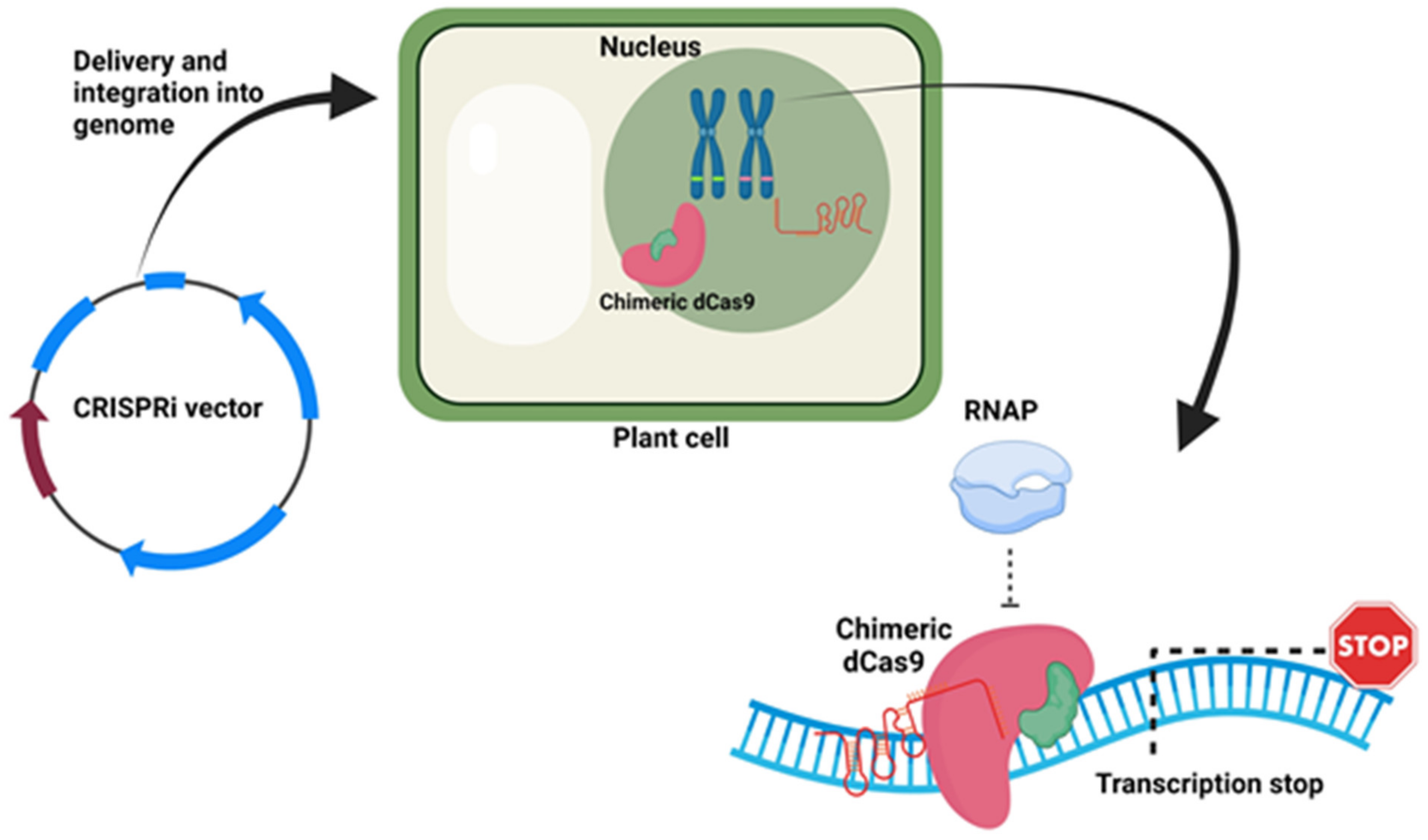
9.4. CRISPRi Vectors
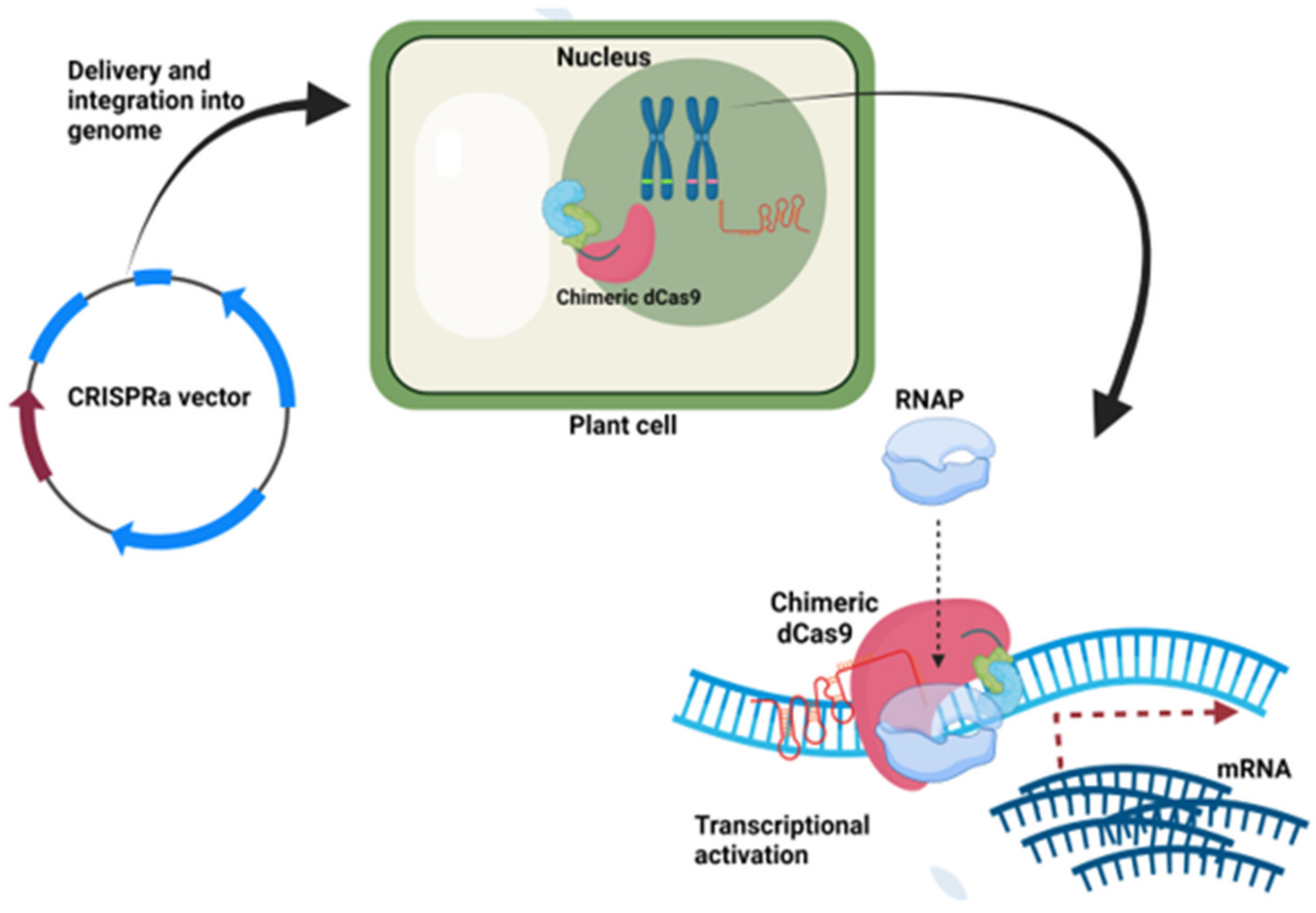
9.5. CRISPRa Vectors
9.6. CRISPR Vectors for Visualization
9.7. Virus-Based CRISPR Vectors
9.8. CRISPR Vectors for Transient Expression
9.9. CRISPR Cloning/Assembly Vectors
9.10. Base Editing Vectors
9.11. Prime Editing Vectors
10. Basis of Choosing the Right Vector
- (i)
- PAM: which PAM (NGG or NTT or NAG, etc.) is present in the target gene? The type of PAM sequence present in the target sequence will determine the Cas endonuclease and the vector carrying it.
- (ii)
- gRNA: single or multiple?
- (iii)
- Copy number of CRISPR vector: high or low?
- (iv)
- Cloning sites and strategies: restriction enzyme based, golden gate assembly, entry-destination based cloning, and ligation independent cloning, etc.
- (v)
- Antibiotic resistance for bacterial and plant selection.
11. Cloning Strategies in Vectors Derived from Different Backbone
12. Conclusions
Author Contributions
Funding
Institutional Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beumer, K.J.; Trautman, J.K.; Christian, M.; Dahlem, T.J.; Lake, C.M.; Hawley, R.S.; Grunwald, D.J.; Voytas, D.; Carroll, D. Comparing Zinc Finger Nucleases and Transcription Activator-Like Effector Nucleases for Gene Targeting in Drosophila. G3 Genes Genomes Genet. 2013, 3, 1717–1725. [Google Scholar] [CrossRef] [Green Version]
- Razzaq, A.; Saleem, F.; Kanwal, M.; Mustafa, G.; Yousaf, S.; Arshad, H.M.I.; Hameed, M.K.; Khan, M.S.; KhanJoyia, F.A. Mod-ern trends in plant genome editing: An inclusive review of the CRISPR/Cas9 Toolbox. Int. J. Mol. Sci. 2019, 20, 4045. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-J.; Orlova, N.; Oakes, B.L.; Ma, E.; Spinner, H.B.; Baney, K.L.M.; Chuck, J.; Tan, D.; Knott, G.J.; Harrington, L.B.; et al. CasX enzymes comprise a distinct family of RNA-guided genome editors. Nature 2019, 566, 218–223. [Google Scholar] [CrossRef]
- Pausch, P.; Al-Shayeb, B.; Bisom-Rapp, E.; Tsuchida, C.A.; Li, Z.; Cress, B.F.; Knott, G.J.; Jacobsen, S.E.; Banfield, J.F.; Doudna, J.A. CRISPR-CasΦ from huge phages is a hypercompact genome editor. Science 2020, 369, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.K.; Kumar, J.; Alok, A.; Tuli, R. RNA-guided genome editing for target gene mutations in wheat. G3 Genes Genomes Genet. 2013, 3, 2233–2238. [Google Scholar] [CrossRef] [Green Version]
- Kaur, N.; Alok, A.; Shivani; Kumar, P.; Kaur, N.; Awasthi, P.; Chaturvedi, S.; Pandey, P.; Pandey, A.; Pandey, A.K.; et al. CRISPR/Cas9 directed editing of lycopene epsilon-cyclase modulates metabolic flux for β-carotene biosynthesis in banana fruit. Metab. Eng. 2020, 59, 76–86. [Google Scholar] [CrossRef]
- Alok, A.; Kumar, J.; Jogam, P.; Sandhya, D. CRISPR/Cas9-mediated gene editing tool and fathomless genetic and meta-bolic engineering applications in plants. In Recent Trends and Techniques in Plant Metabolic Engineering; Springer: Singapore, 2018; pp. 167–179. [Google Scholar]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.; Lin, C.-Y.; Gootenberg, J.; Konermann, S.; Trevino, A.E.; Scott, D.A.; Inoue, A.; Matoba, S.; Zhang, Y.; et al. Double Nicking by RNA-Guided CRISPR Cas9 for Enhanced Genome Editing Specificity. Cell 2013, 154, 1380–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mali, P.; Aach, J.; Stranges, P.; Esvelt, K.; Moosburner, M.; Kosuri, S.; Yang, L.; Church, G.M. CAS9 transcriptional activators for target specificity screening and paired nickases for cooperative genome engineering. Nat. Biotechnol. 2013, 31, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, P.; A Scott, D.; Weinstein, J.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.; Fine, E.; Wu, X.; Shalem, O.; et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2013, 31, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Nishimasu, H.; Ran, F.A.; Hsu, P.; Konermann, S.; Shehata, S.I.; Dohmae, N.; Ishitani, R.; Zhang, F.; Nureki, O. Crystal Structure of Cas9 in Complex with Guide RNA and Target DNA. Cell 2014, 156, 935–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimasu, H.; Cong, L.; Yan, W.X.; Ran, F.A.; Zetsche, B.; Li, Y.; Kurabayashi, A.; Ishitani, R.; Zhang, F.; Nureki, O. Crystal Structure of Staphylococcus aureus Cas9. Cell 2015, 162, 1113–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esvelt, K.M.; Mali, P.; Braff, J.L.; Moosburner, M.; Yaung, S.; Church, G.M. Orthogonal Cas9 proteins for RNA-guided gene regulation and editing. Nat. Methods 2013, 10, 1116–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonfara, I.; Le Rhun, A.; Chylinski, K.; Makarova, K.S.; Lécrivain, A.-L.; Bzdrenga, J.; Koonin, E.V.; Charpentier, E. Phylogeny of Cas9 determines functional exchangeability of dual-RNA and Cas9 among orthologous type II CRISPR-Cas systems. Nucleic Acids Res. 2013, 42, 2577–2590. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Lee, S.; Kim, Y.; Park, J.; Min, S.; Choi, J.W.; Huang, T.P.; Yoon, S.; Liu, D.R.; Kim, H.H. High-throughput analysis of the activities of xCas9, SpCas9-NG and SpCas9 at matched and mismatched target sequences in human cells. Nat. Biomed. Eng. 2020, 4, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Kuang, Y.; Ren, B.; Yan, D.; Yan, F.; Spetz, C.; Sun, W.; Wang, G.; Zhou, X.; Zhou, H. SpRY greatly expands the genome editing scope in rice with highly flexible PAM recognition. Genome Biol. 2021, 22, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kleinstiver, B.; Prew, M.S.; Tsai, S.Q.; Topkar, V.V.; Nguyen, N.T.; Zheng, Z.; Gonzales, A.P.W.; Li, Z.; Peterson, R.T.; Yeh, J.-R.J.; et al. Engineered CRISPR-Cas9 nucleases with altered PAM specificities. Nature 2015, 523, 481–485. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.H.; Miller, S.; Geurts, M.H.; Tang, W.; Chen, L.; Sun, N.; Zeina, C.M.; Gao, X.; Rees, H.A.; Lin, Z.; et al. Evolved Cas9 variants with broad PAM compatibility and high DNA specificity. Nature 2018, 556, 57–63. [Google Scholar] [CrossRef]
- Guilinger, J.P.; Thompson, D.B.; Liu, D.R. Fusion of catalytically inactive Cas9 to FokI nuclease improves the specificity of genome modification. Nat. Biotechnol. 2014, 32, 577–582. [Google Scholar] [CrossRef]
- Endo, M.; Mikami, M.; Toki, S. Biallelic Gene Targeting in Rice. Plant Physiol. 2015, 170, 667–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zetsche, B.; Gootenberg, J.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Cox, D.B.; Yan, D.B.T.C.W.X.; Manteiga, D.B.T.C.J.C.; Schneider, M.W.; Yamano, T.; Nishimasu, H.; Nureki, T.Y.H.N.O.; Crosetto, N.; Zhang, L.G.D.B.T.C.W.X.Y.M.W.S.F. Engineered Cpf1 variants with altered PAM specificities. Nat. Biotechnol. 2017, 35, 789–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michno, J.-M.; Wang, X.; Liu, J.; Curtin, S.J.; Kono, T.; Stupar, R.M. CRISPR/Cas mutagenesis of soybean and Medicago truncatula using a new web-tool and a modified Cas9 enzyme. GM Crop. Food 2015, 6, 243–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nekrasov, V.; Staskawicz, B.; Weigel, D.; Jones, J.D.G.; Kamoun, S. Targeted mutagenesis in the model plant Nicotiana ben-thamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 691–693. [Google Scholar] [CrossRef] [PubMed]
- Eid, A.; Ali, Z.; Mahfouz, M.M. High efficiency of targeted mutagenesis in arabidopsis via meiotic promoter-driven ex-pression of Cas9 endonuclease. Plant Cell Rep. 2016, 35, 1555–1558. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.W.; Kim, J.; Kwon, S.I.; Corvalán, C.; Cho, S.W.; Kim, H.; Kim, S.-G.; Kim, S.-T.; Choe, S.; Kim, J.-S.; et al. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nat. Biotechnol. 2015, 33, 1162–1164. [Google Scholar] [CrossRef] [PubMed]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.K.; Cigan, A.M. Genome editing in maize directed by CRISPR–Cas9 ribonucleoprotein complexes. Nat. Commun. 2016, 7, 13274. [Google Scholar] [CrossRef]
- Malnoy, M.; Viola, R.; Jung, M.-H.; Koo, O.-J.; Kim, S.; Kim, J.-S.; Velasco, R.; Kanchiswamy, C.N. DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar] [CrossRef]
- Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 14261. [Google Scholar] [CrossRef]
- Xie, K.; Yang, Y. RNA-Guided Genome Editing in Plants Using a CRISPR–Cas System. Mol. Plant 2013, 6, 1975–1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarty, N.S.; Graham, A.E.; Studená, L.; Ledesma-Amaro, R. Multiplexed CRISPR technologies for gene editing and tran-scriptional regulation. Nat. Commun. 2020, 11, 1281. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol. 2020, 38, 824–844. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.; Lander, E.S.; Zhang, F. Development and Applications of CRISPR-Cas9 for Genome Engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, K.; Minkenberg, B.; Yang, Y. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proc. Natl. Acad. Sci. USA 2015, 112, 3570–3575. [Google Scholar] [CrossRef] [Green Version]
- Xing, H.-L.; Dong, L.; Wang, Z.-P.; Zhang, H.-Y.; Han, C.-Y.; Liu, B.; Wang, X.-C.; Chen, Q.-J. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014, 14, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čermák, T.; Curtin, S.J.; Gil Humanes, J.; Čegan, R.; Kono, T.; Konečná, E.; Belanto, J.J.; Starker, C.; Mathre, J.W.; Greenstein, R.L.; et al. A Multipurpose Toolkit to Enable Advanced Genome Engineering in Plants. Plant Cell 2017, 29, 1196–1217. [Google Scholar] [CrossRef] [Green Version]
- Haurwitz, R.E.; Sternberg, S.H.; Doudna, J.A. Csy4 relies on an unusual catalytic dyad to position and cleave CRISPR RNA. EMBO J. 2012, 31, 2824–2832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Przybilski, R.; Richter, C.; Gristwood, T.; Clulow, J.S.; Vercoe, R.B.; Fineran, P.C. Csy4 is responsible for CRISPR RNA processing inPectobacterium atrosepticum. RNA Biol. 2011, 8, 517–528. [Google Scholar] [CrossRef] [Green Version]
- Nissim, L.; Perli, S.D.; Fridkin, A.; Perez-Pinera, P.; Lu, T.K. Multiplexed and Programmable Regulation of Gene Networks with an Integrated RNA and CRISPR/Cas Toolkit in Human Cells. Mol. Cell 2014, 54, 698–710. [Google Scholar] [CrossRef] [Green Version]
- Qi, W.; Zhu, T.; Tian, Z.; Li, C.; Zhang, W.; Song, R. High-efficiency CRISPR/Cas9 multiplex gene editing using the glycine tRNA-processing system-based strategy in maize. BMC Biotechnol. 2016, 16, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, F.; Xie, K.; Chen, Y.; Yang, Y.; Mao, Y. Polycistronic tRNA and CRISPR guide-RNA enables highly efficient multiplexed genome engineering in human cells. Biochem. Biophys. Res. Commun. 2016, 482, 889–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Zheng, X.; Qi, Y.; Zhang, D.; Cheng, Y.; Tang, A.; Voytas, D.F.; Zhang, Y. A Single Transcript CRISPR-Cas9 System for Efficient Genome Editing in Plants. Mol. Plant 2016, 9, 1088–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, W.; Chen, D.; Kaufmann, K. Efficient multiplex mutagenesis by RNA-guided Cas9 and its use in the characterization of regulatory elements in the AGAMOUS gene. Plant Methods 2016, 12, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.F.; Norville, J.E.; Aach, J.; McCormack, M.; Zhang, D.; Bush, J.; Church, G.M.; Sheen, J. Multiplex and homologous recom-bination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nat. Biotechnol. 2013, 31, 688–691. [Google Scholar] [CrossRef] [PubMed]
- McCracken, S.; Fong, N.; Yankulov, K.; Ballantyne, S.; Pan, G.; Greenblatt, J.; Patterson, S.D.; Wickens, M.; Bentley, D. The C-terminal domain of RNA polymerase II couples mRNA processing to transcription. Nature 1997, 385, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Hamm, J.; Mattaj, I. Monomethylated cap structures facilitate RNA export from the nucleus. Cell 1990, 63, 109–118. [Google Scholar] [CrossRef]
- Ranganathan, V.; Wahlin, K.; Maruotti, J.; Zack, D.J. Expansion of the CRISPR–Cas9 genome targeting space through the use of H1 promoter-expressed guide RNAs. Nat. Commun. 2014, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.-C.; Hsu, M.-N.; Chang, C.-W.; Lin, M.-W.; Hwu, J.R.; Tu, Y.; Hu, Y.-C. Synthetic switch to minimize CRISPR off-target effects by self-restricting Cas9 transcription and translation. Nucleic Acids Res. 2018, 47, e13. [Google Scholar] [CrossRef] [Green Version]
- Nandy, S.; Pathak, B.; Zhao, S.; Srivastava, V. Heat-shock-inducible CRISPR/Cas9 system generates heritable muta-tions in rice. Plant Direct 2019, 3, e00145. [Google Scholar] [CrossRef] [Green Version]
- Zuo, J.; Niu, Q.W.; Chua, N.H. Technical advance: An estrogen receptor-based transactivator XVE mediates highly induc-ible gene expression in transgenic plants. Plant J. 2000, 2, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.-J.; Uchii, S.; Watanabe, S.; Ezura, H. A Highly Efficient Transformation Protocol for Micro-Tom, a Model Cultivar for Tomato Functional Genomics. Plant Cell Physiol. 2006, 47, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.K.; Solanke, A.U.; Jani, D.; Singh, Y.; Sharma, A.K. A simple and efficient Agrobacterium-mediated procedure for transformation of tomato. J. Biosci. 2009, 34, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Alok, A.; Kaur, J.; Tiwari, S. Functional characterization of wheat myo-inositol oxygenase promoter under different abiotic stress conditions in Arabidopsis thaliana. Biotechnol. Lett. 2020, 42, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, J.G.; Alves, S.C.; Smedley, M.; Snape, J.W.; Harwood, W.A. High-throughput Agrobacterium-mediated barley transformation. Plant Methods 2008, 4, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smedley, M.A.; Harwood, W.A. Gateway®-Compatible Plant Transformation Vectors. In Agrobacterium Protocols; Springer: New York, NY, USA; pp. 3–16. [CrossRef]
- Hayta, S.; Smedley, M.A.; Demir, S.U.; Blundell, R.; Hinchliffe, A.; Atkinson, N.; Harwood, W.A. An efficient and reproducible Agrobacterium-mediated transformation method for hexaploid wheat (Triticum aestivum L.). Plant Methods 2019, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Acanda, Y.; Canton, M.; Zale, J. Efficient Biolistic Transformation of Immature Citrus Rootstocks Using Phosphomannose-isomerase Selection. Plants 2019, 8, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellens, R.; Mullineaux, P.; Klee, H. Technical Focus:A guide to Agrobacterium binary Ti vectors. Trends Plant Sci. 2000, 5, 446–451. [Google Scholar] [CrossRef]
- Hellens, R.P.; Edwards, E.A.; Leyland, N.R.; Bean, S.; Mullineaux, P.M. pGreen: A versatile and flexible binary Ti vector for Agrobacterium-mediated plant transformation. Plant Mol. Biol. 2000, 42, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liang, Z.; Zong, Y.; Wang, Y.; Liu, J.; Chen, K.; Qiu, J.-L.; Gao, C. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 2016, 7, 12617. [Google Scholar] [CrossRef] [Green Version]
- Waltz, E. Gene-edited CRISPR mushroom escapes US regulation. Nature 2016, 532, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Ren, Q.; Yang, L.; Bao, Y.; Zhong, Z.; He, Y.; Liu, S.; Qi, C.; Liu, B.; Wang, Y.; et al. Single transcript unit CRISPR 2.0 systems for robust Cas9 and Cas12a mediated plant genome editing. Plant Biotechnol. J. 2018, 17, 1431–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Zhang, Y.; Lu, M.-H.; Chai, Y.-P.; Jiang, Y.-Y.; Zhou, Y.; Wang, X.-C.; Chen, Q.-J. A Novel Ternary Vector System United with Morphogenic Genes Enhances CRISPR/Cas Delivery in Maize. Plant Physiol. 2019, 181, 1441–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grützner, R.; Martin, P.; Horn, C.; Mortensen, S.; Cram, E.J.; Lee-Parsons, C.W.; Stuttmann, J.; Marillonnet, S. High-efficiency genome editing in plants mediated by a Cas9 gene containing multiple introns. Plant Commun. 2020, 2, 100135. [Google Scholar] [CrossRef] [PubMed]
- Čermák, T.; Baltes, N.J.; Čegan, R.; Zhang, Y.; Voytas, D.F. High-frequency, precise modification of the tomato genome. Genome Biol. 2015, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Danilo, B.; Perrot, L.; Botton, E.; Nogué, F.; Mazier, M. The DFR locus: A smart landing pad for targeted transgene insertion in tomato. PLoS ONE 2018, 13, e0208395. [Google Scholar] [CrossRef]
- Dahan-Meir, T.; Filler-Hayut, S.; Melamed-Bessudo, C.; Bocobza, S.; Czosnek, H.; Aharoni, A.; Levy, A.A. Efficient in planta gene targeting in tomato using geminiviral replicons and the CRISPR/Cas9 system. Plant J. 2018, 95, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, Z.; Shami, A.; Sedeek, K.; Kamel, R.; Alhabsi, A.; Tehseen, M.; Hassan, N.; Butt, H.; Kababji, A.; Hamdan, S.M.; et al. Fusion of the Cas9 endonuclease and the VirD2 relaxase facilitates homology-directed repair for precise genome engineering in rice. Commun. Biol. 2020, 3, 1–13. [Google Scholar] [CrossRef]
- Lowder, L.G.; Zhang, D.; Baltes, N.J.; Paul, J.W.; Tang, X.; Zheng, X.; Voytas, D.; Hsieh, T.-F.; Zhang, Y.; Qi, Y. A CRISPR/Cas9 Toolbox for Multiplexed Plant Genome Editing and Transcriptional Regulation. Plant Physiol. 2015, 169, 971–985. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Vilar, M.; Bernabé-Orts, J.M.; Fernandez-Del-Carmen, A.; Ziarsolo, P.; Blanca, J.; Granell, A.; Orzaez, D. A modular toolbox for gRNA–Cas9 genome engineering in plants based on the GoldenBraid standard. Plant Methods 2016, 12, 10. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Naseri, A.; Reyes-Gutierrez, P.; Wolfe, S.A.; Zhang, S.; Pederson, T. Multicolor CRISPR labeling of chromosomal loci in human cells. Proc. Natl. Acad. Sci. USA 2015, 112, 3002–3007. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, S.; Matsunaga, S. Visualization of Chromatin Loci with Transiently Expressed CRISPR/Cas9 in Plants. Cytologia 2017, 82, 559–562. [Google Scholar] [CrossRef]
- Wang, X.; Ye, L.; Lyu, M.; Ursache, R.; Löytynoja, A.; Mähönen, A.P. An inducible genome editing system for plants. Nat. Plants 2020, 6, 766–772. [Google Scholar] [CrossRef]
- Sretenovic, S.; Yin, D.; Levav, A.; Selengut, J.D.; Mount, S.M.; Qi, Y. Expanding plant genome-editing scope by an engineered iSpyMacCas9 system that targets A-rich PAM sequences. Plant Commun. 2020, 2, 100101. [Google Scholar] [CrossRef]
- Zhong, Z.; Zhang, Y.; You, Q.; Tang, X.; Ren, Q.; Liu, S.; Yang, L.; Wang, Y.; Liu, X.; Liu, B.; et al. Plant Genome Editing Using FnCpf1 and LbCpf1 Nucleases at Redefined and Altered PAM Sites. Mol. Plant 2018, 11, 999–1002. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ren, Q.; Tang, X.; Liu, S.; Malzahn, A.A.; Zhou, J.; Wang, J.; Yin, D.; Pan, C.; Yuan, M.; et al. Expanding the scope of plant genome engineering with Cas12a orthologs and highly multiplexable editing systems. Nat. Commun. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Hahn, F.; Korolev, A.; Loures, L.S.; Nekrasov, V. A modular cloning toolkit for genome editing in plants. BMC Plant Biol. 2020, 20, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ren, Q.; Sretenovic, S.; Liu, S.; Tang, X.; Huang, L.; He, Y.; Liu, L.; Guo, Y.; Zhong, Z.; Liu, G.; et al. PAM-less plant genome editing using a CRISPR–SpRY toolbox. Nat. Plants 2021, 7, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Lowder, L.G.; Zhang, T.; Malzahn, A.A.; Zheng, X.; Voytas, D.F.; Zhong, Z.; Chen, Y.; Ren, Q.; Li, Q.; et al. A CRISPR–Cpf1 system for efficient genome editing and transcriptional repression in plants. Nat. Plants 2017, 3, 17018. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Lucke, M.; Jang, Y.-T.; Zhu, W.; Symeonidi, E.; Wang, C.; Fitz, J.; Xi, W.; Schwab, R.; Weigel, D. An efficient CRISPR vector toolbox for engineering large deletions in Arabidopsis thaliana. Plant Methods 2018, 14, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Zou, W.; Xu, H.; Liang, Y.; Huang, B. Efficient labeling and imaging of protein-coding genes in living cells using CRISPR-Tag. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Ali, Z.; Abulfaraj, A.; Li, L.; Ghosh, N.; Piatek, M.; Mahjoub, A.; Aouida, M.; Piatek, A.; Baltes, N.J.; Voytas, D.; et al. Efficient Virus-Mediated Genome Editing in Plants Using the CRISPR/Cas9 System. Mol. Plant 2015, 8, 1288–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltes, N.J.; Gil Humanes, J.; Cermak, T.; Atkins, P.A.; Voytas, D.F. DNA Replicons for Plant Genome Engineering. Plant Cell 2014, 26, 151–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil Humanes, J.; Wang, Y.; Liang, Z.; Shan, Q.; Ozuna, C.V.; Sánchez-León, S.; Baltes, N.J.; Starker, C.; Barro, F.; Gao, C.; et al. High-efficiency gene targeting in hexaploid wheat using DNA replicons and CRISPR /Cas9. Plant J. 2017, 89, 1251–1262. [Google Scholar] [CrossRef] [Green Version]
- Fauser, F.; Schiml, S.; Puchta, H. Both CRISPR/Cas-based nucleases and nickases can be used efficiently for genome engineering inArabidopsis thaliana. Plant J. 2014, 79, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Wang, Y.; Li, C.; Zhang, R.; Chen, K.; Ran, Y.; Qiu, J.-L.; Wang, D.; Gao, C. Precise base editing in rice, wheat and maize with a Cas9-cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 438–440. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Wan, T.; Ping, Y. Off-targeting of base editors: BE3 but not ABE induces substantial off-target single nucleotide vari-ants. Signal Transduct. Target. Ther. 2019, 4, 9. [Google Scholar]
- Kang, B.-C.; Yun, J.-Y.; Kim, S.-T.; Shin, Y.; Ryu, J.; Choi, M.; Woo, J.W.; Kim, J.-S. Precision genome engineering through adenine base editing in plants. Nat. Plants 2018, 4, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Sretenovic, S.; Ren, Q.; Jia, X.; Li, M.; Fan, T.; Yin, D.; Xiang, S.; Guo, Y.; Liu, L.; et al. Plant Prime Editors Enable Precise Gene Editing in Rice Cells. Mol. Plant 2020, 13, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Zong, Y.; Xue, C.; Wang, S.; Jin, S.; Zhu, Z.; Wang, Y.; Anzalone, A.V.; Raguram, A.; Doman, J.L.; et al. Prime genome editing in rice and wheat. Nat. Biotechnol. 2020, 38, 582–585. [Google Scholar] [CrossRef]
- Gerard, G.F. The role of template-primer in protection of reverse transcriptase from thermal inactivation. Nucleic Acids Res. 2002, 30, 3118–3129. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, X.; Li, K.; Wang, D.; Ding, Y.; Liu, X.; Luo, J.; Fang, C. A simple and efficient cloning system for CRISPR/Cas9-mediated genome editing in rice. PeerJ 2020, 8, e8491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Z.; Zhang, B.; Ding, W.; Liu, X.; Yang, D.-L.; Wei, P.; Cao, F.; Zhu, S.; Zhang, F.; Mao, Y.; et al. Efficient genome editing in plants using a CRISPR/Cas system. Cell Res. 2013, 23, 1229–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.A.; El-Sayed, A.; Akbar, A.; Mangravita-Novo, A.; Bibi, S.; Afzal, Z.; Norman, D.J.; Ali, G.S. A highly effi-cient ligation-independent cloning system for CRISPR/Cas9 based genome editing in plants. Plant Methods 2017, 13, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, S.K. Genome Engineering for Crop Improvement; Upadhyay, S.K., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2021. [Google Scholar] [CrossRef]
- Sushmita; Kaur, G.; Upadhyay, S.K.; Verma, P.C. An Overview of Genome-Engineering Methods. Genome Eng. Crop Improv. 2021, 1–21. [Google Scholar] [CrossRef]
- Altae-Tran, H.; Kannan, S.; Demircioglu, F.E.; Oshiro, R.; Nety, S.P.; McKay, L.J.; Dlakić, M.; Inskeep, W.P.; Makarova, K.S.; Macrae, R.K.; et al. The widespread IS200/605 transposon family encodes diverse programmable RNA-guided endonucleases. Science 2021. [Google Scholar] [CrossRef]
- Alok, A.; Sandhya, D.; Jogam, P.; Rodrigues, V.; Bhati, K.K.; Sharma, H.; Kumar, J. The Rise of the CRISPR/Cpf1 System for Efficient Genome Editing in Plants. Front. Plant Sci. 2020, 11, 264. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cas Endonuclease | Size | Bacterial Source | PAM Recognition Site | References |
|---|---|---|---|---|
| SpCas9 | 4104 bp | Streptococcus pyogenes | 3’ NGG | [19] |
| xCas9 3.7 | 4140 bp | Streptococcus pyogenes | NG, NNG, GAT, and CAA | [17,20] |
| SpCas9-NG | Streptococcus pyogenes | NG | [17] | |
| SpRY | Streptococcus pyogenes | PAM-less | [18] | |
| SpG | Streptococcus pyogenes | NGD | [18] | |
| StCas9 | 3.4 kb | Streptococcus thermophiles | NNAGAAW | [19] |
| SaCas9 | 3156 bp | Staphylococcus aureus | 3’ NNGRRT or NNGRR(N) | [19] |
| SpCas9 D1135E variant | 4104 bp | Streptococcus pyogenes | 3’ NGG | [19] |
| SpCas9 VRER variant | 4104 bp | Streptococcus pyogenes | 3’ NGCG | [19] |
| SpCas9 EQR variant | 1716 bp | Streptococcus pyogenes | 3’ NGAG | [19] |
| SpCas9 VQR variant | 4104 bp | Streptococcus pyogenes | 3’ NGAN or NGNG | [19] |
| SpCas9 nickases | 4.1 kb | Streptococcus pyogenes | Enhanced specificity,3’ NGG | [10] |
| Dimeric FokI-dCas9 fusions | [21] | |||
| NmCas9 | 3243bp | Neisseria meningitidis | NNNNGMTT | [15] |
| FnCas12a | 3.9 kb | Francisella novicida | TTN, CTA | [22] |
| AsCas12a | 3921bp | Acidaminococcus sp. | 5’ TTTV | [23] |
| AsCas12aRR variant | 3918 bp | Acidaminococcus sp | 5’ TYCV | [23] |
| AsCas12a RVR variant | 3918 bp | Acidaminococcus sp | 5’ TATV | [23,24] |
| LbCas12a | 3684 bp | Lachnospiraceae bacterium | 5’ TTTV | [23] |
| LbCas12a RR variant | 3681 bp | Lachnospiraceae bacterium | 5’ TYCV | [23,24] |
| AacCas12b | 3386 bp | Alicyclobacillus acidoterrestris | 5’ NTTV, VTTTV | [25] |
| AaCas12b | 3486 bp | Alicyclobacillus acidiphilus | 5’ NTTV, VTTTV | [25] |
| BthCas12b | 3423 bp | Bacillus thermoamylovorans | 5’ NTTV | [25] |
| BhCas12b | 3423 bp | Bacillus hisashii | 5’ NTTV | [25] |
| Cas12e (or DpbCasX) | 2958bp | Deltaproteobacteria | 5’TTCN | [3] |
| Cas12j (also known as Casɸ) | 2142bp | Phage | 5’TTA | [4] |
| CRISPR/Cas Vectors | Size (Kb) | Bacterial Selection | Replication Origin Agro, E. coli | Plant Selection Gene | Remark | References |
|---|---|---|---|---|---|---|
| Knock out Vectors | ||||||
| pRGEB31 | 15.0 | Kan | pVS1, ColE1 | Hyg | Cas9 regulated under CaMV35; gRNA under OsU6 | [32] |
| pHSN401 | 12.7 | Kan | pSA, pUC | Hyg | Cas9 regulated under CaMV35; gRNA under AtU6 | [37] |
| pRGEB32 | 15.8 | Kan | pVS1, ColE1 | Hyg | Cas9 regulated with maize Ubi promoter | [36] |
| pBSN401 | 12.5 | Kan | pSA, pUC | Bar | Cas9 regulated under CaMV35; gRNA under AtU6 | [37] |
| pKSE401 | 12.5 | Kan | pSA, pUC | Kan | Cas9 regulated under CaMV35; gRNA under AtU6 | [37] |
| pHSE401 | 16.6 | Kan | pVS1, ColE1 | Hyg | Cas9 regulated under CaMV35; gRNA under AtU6 | [37] |
| pFGC-pcoCas9 | 13.3 | Kan | pVS1, ColE1 | Bas | Plant codon optimised Cas9 under a hybrid 35SPPDK promoter | [46] |
| Cas9 MDC32 | 14.1 | Kan | pVS1, RepA, sta1 | Hyg | Soybean codon optimized Cas9 regulated by 35S promoter | [25] |
| Cas9 MDC123 | 13.6 | Kan | pVS1, RepA, sta1 | Bas | Soybean codon optimized Cas9 | [25] |
| G10 Cas9 MDC123 | 13.2 | Kan | pVS1, RepA, sta1 | Bas | Soybean codon optimized Cas9 regulated by G10 promoter | [25] |
| pDIRECT_22C | 16.0 | Kan | pVS1, ColE1 | Kan | Csy4-P2A fused with Arabidopsis codon optimised Cas9 | [38] |
| pGEL031 | 15.7 | Kan | pVS1, ColE1 | Cas9 regulated with maize Ubi promoter | [64] | |
| pGEL029 | 15.6 | Kan | pVS1, ColE1 | Cas9 regulated with maize Ubi; gRNA without promoter | [64] | |
| pG3H-U6EC1 | 13.6 | Kan | ColE1 | Hyg | pGreen3 derived vector, required pSoup vector for Agrobacterium | [65] |
| pAGM51547 | Kan | pVS1, ColE1 | Bar | Cas9 with multiple introns; incresed editng efficiency | [66] | |
| pAGM55273 | 17.0 | Kan | pVS1, ColE1 | Kan | Cas9 with multiple introns; incresed editng efficiency | [66] |
| Knock-in Vectors | ||||||
| pTC217 | 17.9 | Kan | pVS1, RepA | Kan | Bean Yellow Dwarf Virus replicon based; donor teplated is Pnos:NptII-35S:ANT1 flanked with 5’ and 3’ homology arm | [67] |
| pDe-Cas9-Hpt-GT-DFR#3-DFR#4-DFRtemp | Greater than 16 | Chlor and Spec | pVS1, sta1 | Hyg | Donor DNA left and right homologous arms, each corresponding to the 400 bp and 392 bp sequences flanking both sides of the 1013 bp DFR deletion | [68] |
| TmicT2donorRepUbi10 | NA | Kan | NA | Kan | contains the CRISPR/Cas9, Rep protein on the T-DNA outside the geminiviral replicon and the donor repair template is within the replicon. | [69] |
| TmicT1donorRepUbi10 | NA | Kan | NA | Kan | Same as above | [69] |
| pRGEB-VirD2-Cas9 | 17.2 | Kan | pVS1, ColE1 | Hyg | Chimeric Cas9-VirD under ubiquitin promoter; VirD2 relaxase facilitates homology-directed repair | [70] |
| pTC217 and pTC223 | 17.9 | Kan | pVS1, ColE1 | Kan | gRNA targeting ANT1 locus, donor DNA having 5’ homology arm-Pnos:NptII-35S:ANT13’ homology arm, bean yellow dwarf virus replicon | [67] |
| CRISPRi Vectors | ||||||
| pYPQ153, pYPQ series | 7.7 | Spec | ColE1 | Absent | Plant codon-optimized dCas9 fused with mutated SRDX (X3) repressor | [71] |
| pHSN6I01 | 13.0 | Kan | pSA, ColE1 | Hyg | Maize codon optimized dCas9-KRAB (Krüppel-associated box) | [37] |
| pdCas9 (GB1079) | 7.1 | Amp | f1 ori, ColE1 | Absent | Cas9 coding region with mutated (D10A, H840A) and inactivated catalytic domains (human codon optimised) | [72] |
| pDIRECT_21B | 15.2 | Kan | pVS1, ColE1 | 2x35S:hpt II | 35S:AtCas9_dead + AtU6:gRNA | [38] |
| pDIRECT_21D | 16.2 | Kan | pVS1, ColE1 | 35S:Csy4-P2A-AtCas9_dead + CmYLCV:gRNAs with Csy4 spacers | [38] | |
| pDIRECT_23D | 15.7 | Kan | pVS1, ColE1 | 2x35S:bar | 35S:Csy4-P2A-AtCas9_dead + CmYLCV:gRNAs with Csy4 spacers | [38] |
| CRISPRa Vectors | ||||||
| pEGB 35s:dCas:EDLL:tNos (GB1190) | 12.1 | Kan | pVS1, sta1 | Absent | Human codon optimizedinactivated Cas9 fused to the EDLL transcriptional activator | [72] |
| pYPQ152 | 7.7 | Spec | Absent, ColE1 | Absent | Plant codon-optimized dCas9 fused with VP64 activator | [71] |
| pHSN6A01 | 13.0 | Kan | pSA, ColE1 | Hyg | Maize codon-optimized dCas9 fused with VP64 (4× minimal VP16 activation domain) | [37] |
| pBUN6A11 | 13.8 | Kan | pSA, ColE1 | Bar | Maize codon optimized dCas9 fused with VP64 (4× minimal VP16 activation domain) | [37] |
| Vectors for Visualization | ||||||
| pK7WGF2::hCas9 | 14.4 | Spec | pVS1, ColE1 | Kan | human codon usage Cas9 nuclease with an N-terminal GFP tag | [26] |
| pHAGE-TO-dCas9-3XGFP | 12.9 | Amp | ColE1 | Absent | SpdCas9 fused with 3XsfGFP | [73] |
| pHAGE-EFS-dCas9-GFP | 11.0 | Amp | ColE1 | Absent | SpdCas9 fused with 3XsfGFP | [73] |
| NmdCas9-3xGFP | Amp | ColE1 | Absent | Chimeric NmdCas9 fused with GFP | [74] | |
| SpdCas9-3xCherry | 12.9 | Amp | ColE1 | Absent | Chimeric SpdCas9 fused with 3xCherry P | [74] |
| p221z-CAS9p-TagRFP-t35s | 7.2 | Ble | ColE1 | Absent | Chimeric Cas9 fused withTagRFP | [75] |
| Nickase Vectors | ||||||
| pHSN501 | 12.7 | Kan and Spec | pSA, ColE1 | Hyg | CaMV regulated zCas9D10A, gRNA under AtU6-26 promoter | [37] |
| pBUN501 | 13.4 | Kan and Spec | pSA, ColE1 | Bar | Ubi promoter regulated zCas9D10A, gRNA under AtU6-26 promoter | [37] |
| Transient Expression Vectors | ||||||
| pYPQ166-iSpyMac | 7.7 | Spec | Absent, ColE1 | Absent | Cas9 compatible for NAAR PAM | [76] |
| pYPQ239-RVR | 7.1 | Spec | Absent | Rice codon optimized Francisella tularensis Cas12a (Cpf1) | [77] | |
| pRGE32 | 10.0 | Amp | pVS1, ColE1 | Absent | gRNA under rice snoRNA U3 promoter; Cas9 under with rice ubiquitin promoter | [36] |
| pRGE31 | 9.1 | Amp | pVS1, ColE1 | Absent | gRNA under rice snoRNA U3 promoter | [32] |
| HBT-pcoCas9 and p35SPPDK-pcoCas9 | 7.9 | Amp | Absent, ColE1 | Absent | Hybrid constitutive promoter 35SPPDK regulates Cas9 | [46] |
| Cloning/Assembly/Entry Vectors | ||||||
| p221z-CAS9p-t35s | 6.5 | Ble | Absent, ColE1 | Absent | Based on pDONR-221z | [75] |
| pYPQ131-STU-Fn | 3.1 | Tet | Absent, ColE1 | Absent | For crRNA cloning flanked by HH and HDV ribozymes; compatible with FnCas12a | [78] |
| pFH16 | 6.1 | Spec | ColE1 | Absent | Rice codon optimized FnCas12a | [79] |
| pYPQ166-SpRY | 7.7 | Spec | Absent, ColE1 | Absent | Gateway compatible zSpRYCas9 entry clone; PAM less Cas9 | [80] |
| pYPQ239-RR | 7.1 | Spec | Absent, ColE1 | Absent | Entry for FnCas12a | [77] |
| pMOD_B2101 | 3.5 | Amp | Absent, ColE1 | SapI ccdb cassette for cloning multiple gRNA protospacers with Csy4 spacers | [38] | |
| pMOD_B2301 | 3.6 | Amp | Absent, ColE1 | SapI ccdb cassette for cloning multiple gRNA protospacers with tRNA spacers | [38] | |
| pMOD_B2403 | 3.5 | Amp | Absent, ColE1 | SapI ccdb cassette for cloning multiple gRNA protospacers with ribozyme spacers | [38] | |
| pYPQ230 (LbCpf1) | 7.3 | Spec | Absent, ColE1/pBR322/pUC | Absent | [81] | |
| pBlu/gRNA | 3.5 | Amp | Absent | Direct cloning of target oligo duplex | [25] | |
| pYPQ131-138 | 3.8 | Tet | Absent, ColE1 | Absent | Golden Gate entry vector; one to eighth gRNA under AtU6 promoter | [71] |
| pICSL01009 | 2.3 | Spec | Absent, ColE1 | Absent | [26] | |
| pYPQ141-148 | 3.6 | Spec | Absent, ColE1 | Absent | Golden Gate entry vector; one to eighth gRNA under different U6 promoter | [71] |
| pYPQ141-ZmUbi-RZ-Lb | 4.9 | Spec | Absent, ColE1 | Absent | Lb Cpf1 gRNA cloning site for ribozyme cleavage | [81] |
| pCBC-DT1T2 | 3.5 | Chlor | Absent, ColE1 | Absent | Cloning of two target under AtU6 promoter | [37] |
| pEF005-sgRNA-shuffle-in | 2.9 | Amp | Absent, ColE1 | Absent | Provide and shuffle a cassette of AtU6:sgRNA-transRNA into SM-destination vectors (pRW006 and pRW004) with golden gate cloning strategy. | [82] |
| Viral Replicon Vectors | ||||||
| pTRANS_101 | 5.8 | Spec | Absent, ColE1 | Absent | Non-T-DNA None BeYDV | [38] |
| pTRANS_102 | 6.2 | Spec | Absent, ColE1 | Absent | Non-T-DNA None ToLCV | [38] |
| pTRANS_211 | 12.4 | Spec | pVS1, ColE1 | Hyg | Non-T-DNA None WDV | [38] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alok, A.; Chauhan, H.; Upadhyay, S.K.; Pandey, A.; Kumar, J.; Singh, K. Compendium of Plant-Specific CRISPR Vectors and Their Technical Advantages. Life 2021, 11, 1021. https://doi.org/10.3390/life11101021
Alok A, Chauhan H, Upadhyay SK, Pandey A, Kumar J, Singh K. Compendium of Plant-Specific CRISPR Vectors and Their Technical Advantages. Life. 2021; 11(10):1021. https://doi.org/10.3390/life11101021
Chicago/Turabian StyleAlok, Anshu, Hanny Chauhan, Santosh Kumar Upadhyay, Ashutosh Pandey, Jitendra Kumar, and Kashmir Singh. 2021. "Compendium of Plant-Specific CRISPR Vectors and Their Technical Advantages" Life 11, no. 10: 1021. https://doi.org/10.3390/life11101021