
A Novel Real-Time RT-PCR-Based Methodology for the Preliminary Typing of SARS-CoV-2 Variants, Employing Non-Extendable LNA Oligonucleotides and Three Signature Mutations at the Spike Protein Receptor-Binding Domain

 ,
,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Primers, TaqMan Probes and Non-Extendable Blocking Oligonucleotides
2.2. Real-Time RT-PCR
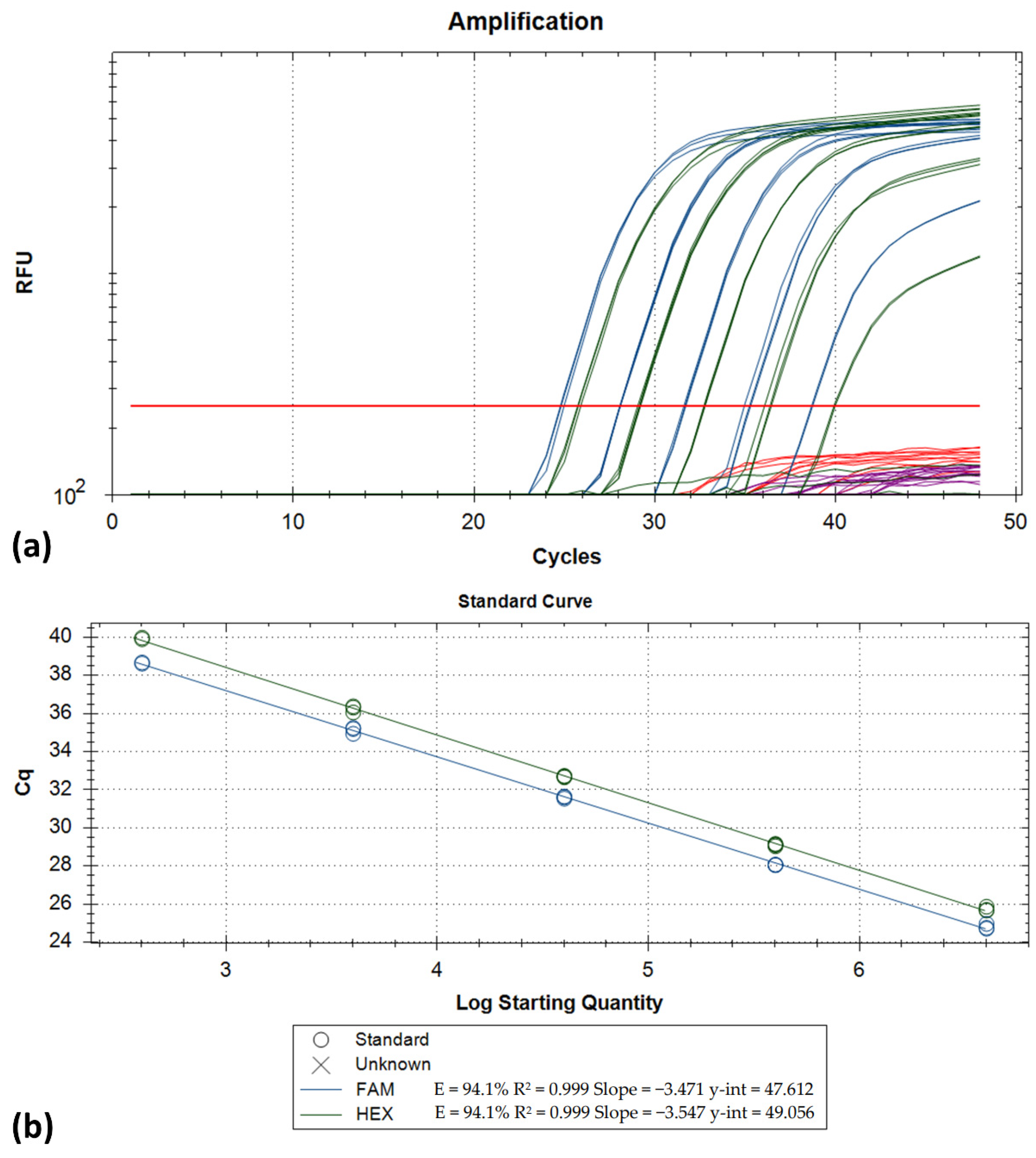
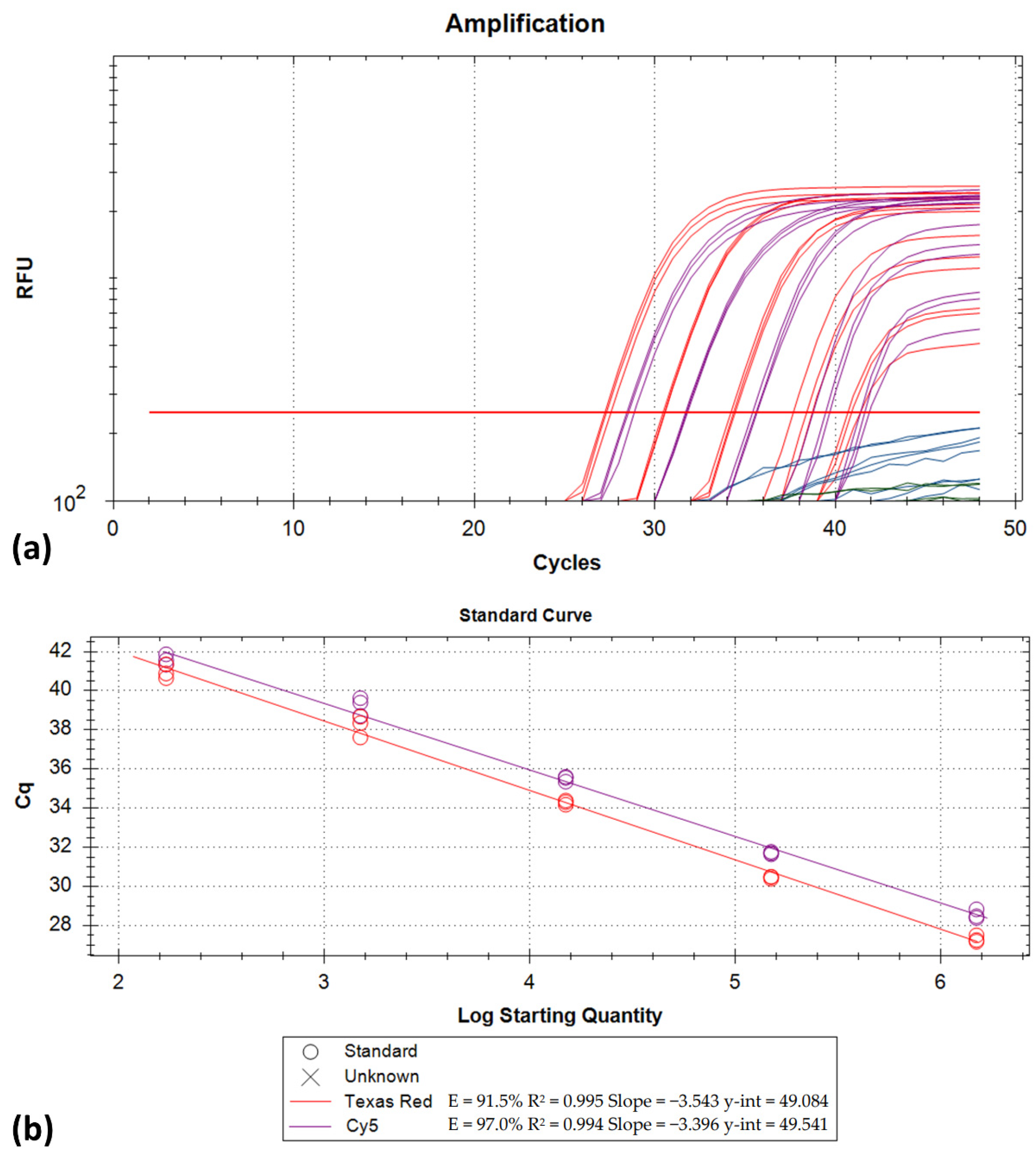
2.3. Analytical Characteristics
2.4. Specificity and Diagnostic Performance
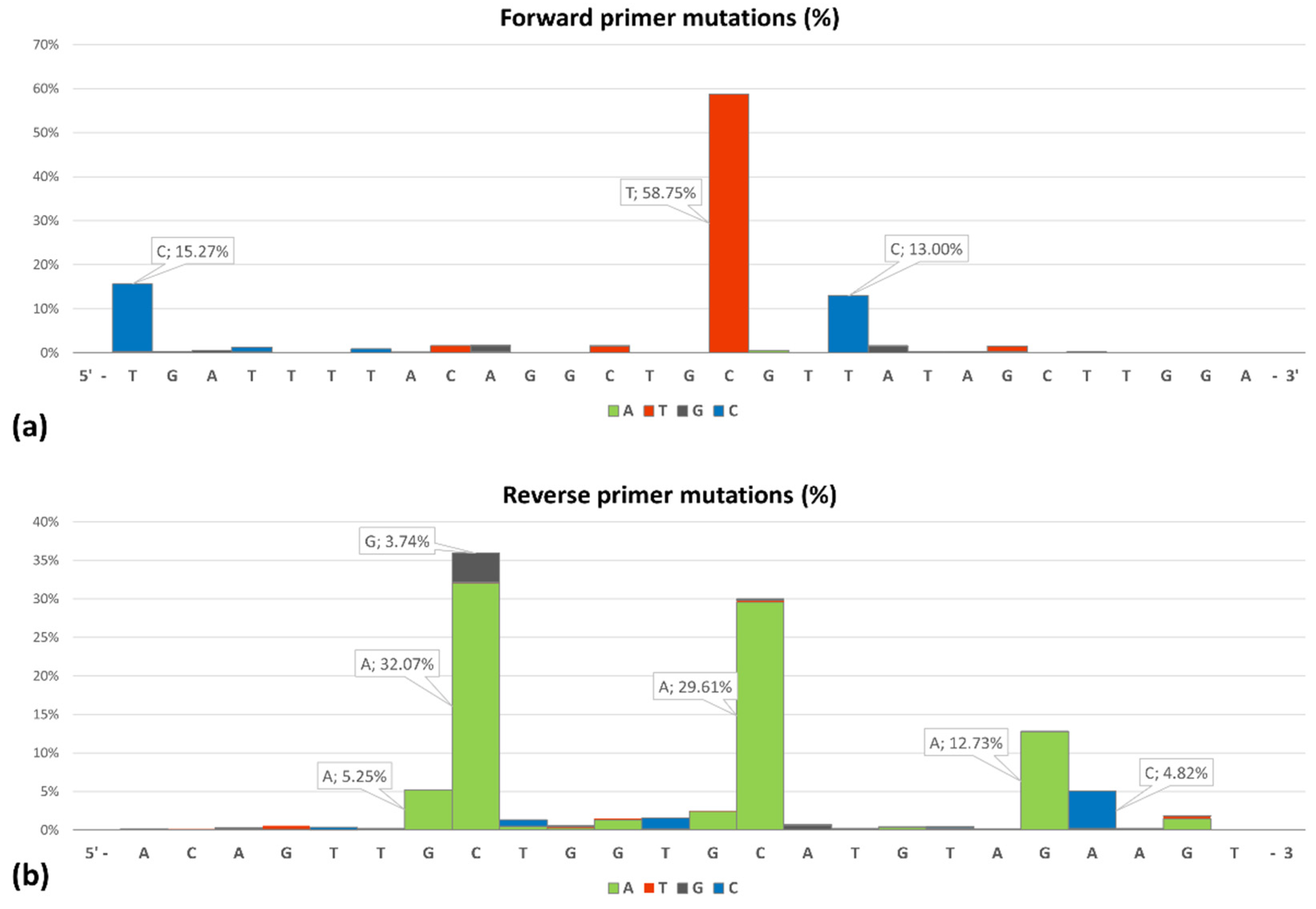
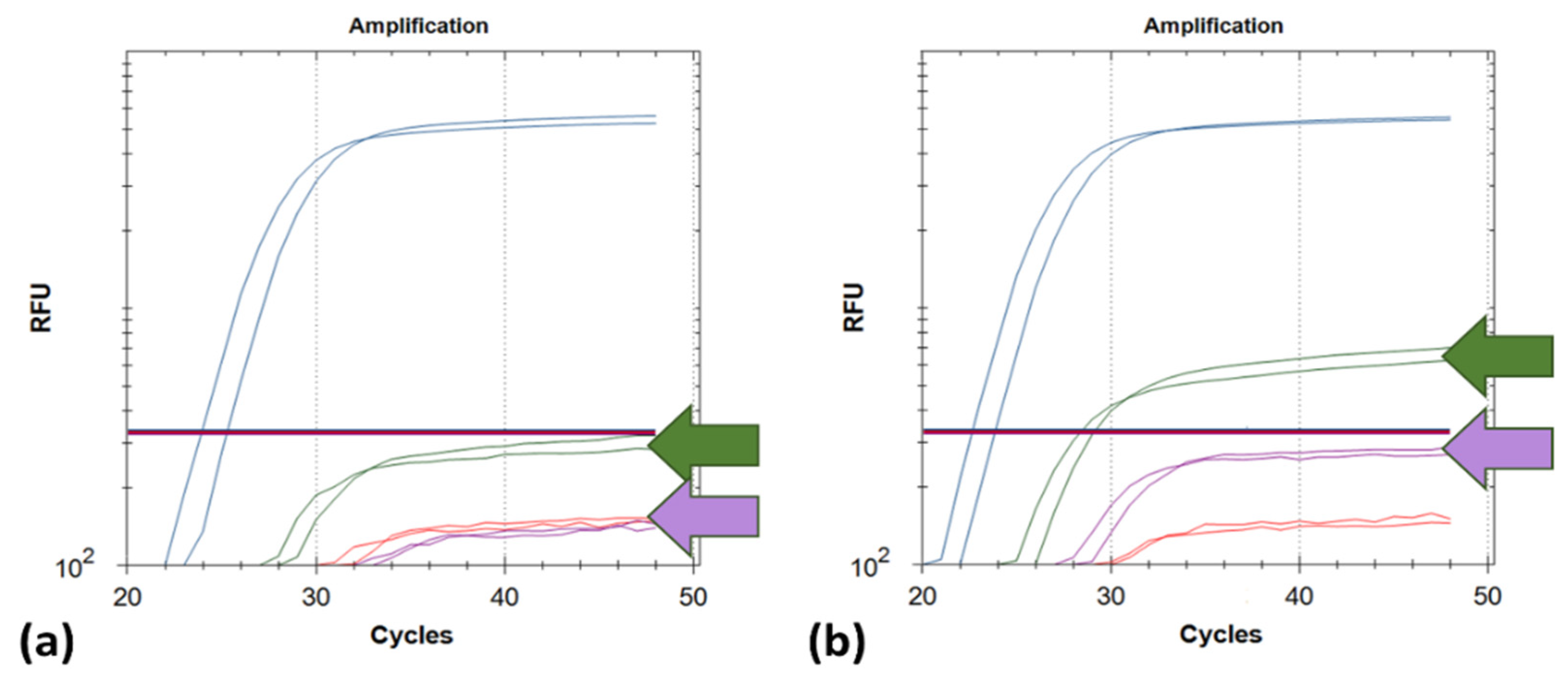
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Caro, A.; Cunha, F.; Petrosillo, N.; Beeching, N.; Ergonul, O.; Petersen, E.; Koopmans, M. SARS-CoV-2 escape mutants and protective immunity from natural infections or immunizations. Clin. Microbiol. Infect. 2021. [Google Scholar] [CrossRef] [PubMed]
- Benton, D.J.; Wrobel, A.G.; Roustan, C.; Borg, A.; Xu, P.; Martin, S.R.; Rosenthal, P.B.; Skehel, J.J.; Gamblin, S.J. The effect of the D614G substitution on the structure of the spike glycoprotein of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2021, 118, e2022586118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cai, Y.; Xiao, T.; Lu, J.; Peng, H.; Sterling, S.M.; Walsh, R.M., Jr.; Rits-Volloch, S.; Zhu, H.; Woosley, A.N.; et al. Structural impact on SARS-CoV-2 spike protein by D614G substitution. Science 2021, 372, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Starr, T.N.; Greaney, A.J.; Hilton, S.K.; Ellis, D.; Crawford, K.H.D.; Dingens, A.S.; Navarro, M.J.; Bowen, J.E.; Tortorici, M.A.; Walls, A.C.; et al. Deep Mutational Scanning of SARS-CoV-2 Receptor Binding Domain Reveals Constraints on Folding and ACE2 Binding. Cell 2020, 182, 1295–1310. [Google Scholar] [CrossRef] [PubMed]
- Wibmer, C.K.; Ayres, F.; Hermanus, T.; Madzivhandila, M.; Kgagudi, P.; Oosthuysen, B.; Lambson, B.E.; de Oliveira, T.; Vermeulen, M.; van der Berg, K.; et al. SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma. Nat. Med. 2021, 1–4. [Google Scholar] [CrossRef]
- CoVariants. Available online: https://covariants.org/shared-mutations (accessed on 18 August 2021).
- Di Giacomo, S.; Mercatelli, D.; Rakhimov, A.; Giorgi, F.M. Preliminary report on severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) Spike mutation T478K. J. Med. Virol. 2021, 93, 5638–5643. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J.; et al. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Public Health England SARS-CoV-2 Variants of Concern and Variants under Investigation in England. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/972247/Variants_of_Concern_VOC_Technical_Briefing_7_England.pdf (accessed on 18 August 2021).
- European Centre for Disease Prevention and Control (ECDC) SARS-CoV-2 Variants of Concern as of 5 August 2021. Available online: https://www.ecdc.europa.eu/en/COVID-19/variants-concern (accessed on 18 August 2021).
- European Centre for Disease Prevention and Control (ECDC) Methods for the Detection and Identification of SARS-CoV-2 Variants. Available online: https://www.ecdc.europa.eu/en/publications-data/methods-detection-and-identification-SARS-CoV-2-variants (accessed on 18 August 2021).
- Chaintoutis, S.C.; Chassalevris, T.; Tsiolas, G.; Balaska, S.; Vlatakis, I.; Mouchtaropoulou, E.; Siarkou, V.I.; Tychala, A.; Koutsioulis, D.; Skoura, L.; et al. A one-step real-time RT-PCR assay for simultaneous typing of SARS-CoV-2 mutations associated with the E484K and N501Y spike protein amino-acid substitutions. J. Virol. Methods 2021, 296, 114242. [Google Scholar] [CrossRef] [PubMed]
- GISAID PrimerChecker. Available online: https://www.epicov.org/epi3/ (accessed on 18 August 2021).
- Markham, N.R.; Zuker, M. DINAMelt web server for nucleic acid melting prediction. Nucleic Acids Res. 2005, 33, W577–W581. [Google Scholar] [CrossRef] [PubMed]
- Chassalevris, T.; Chaintoutis, S.C.; Apostolidi, E.D.; Giadinis, N.D.; Vlemmas, I.; Brellou, G.D.; Dovas, C.I. A highly sensitive semi-nested real-time PCR utilizing oligospermine-conjugated degenerate primers for the detection of diverse strains of small ruminant lentiviruses. Mol. Cell. Probes 2020, 51, 101528. [Google Scholar] [CrossRef] [PubMed]
- IDT PrimeTime qPCR Probes. Available online: https://www.idtdna.com/site/order/qpcr/primetimeprobes/multiplex (accessed on 18 August 2021).
- Lee, S.-T.; Kim, J.-Y.; Kown, M.-J.; Kim, S.W.; Chung, J.H.; Ahn, M.-J.; Oh, Y.L.; Kim, J.-W.; Ki, C.-S. Mutant Enrichment with 3′-Modified Oligonucleotides: A Practical PCR Method for Detecting Trace Mutant DNAs. J. Mol. Diagn. 2011, 13, 657. [Google Scholar] [CrossRef] [PubMed]
- Chaintoutis, S.C.; Siarkou, V.I.; Mylonakis, M.E.; Kazakos, G.M.; Skeva, P.; Bampali, M.; Dimitriou, M.; Dovrolis, N.; Polizopoulou, Z.S.; Karakasiliotis, I.; et al. Limited cross-species transmission and absence of mutations associated with SARS-CoV-2 adaptation in cats: A case study of infection in a small household setting. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, S.; Frost, K.; Colson, B.; Simon, K.; Mäde, D.; Reiting, R.; Gowik, P.; Grohmann, L. Validation of qualitative PCR methods on the basis of mathematical–statistical modelling of the probability of detection. Accredit. Qual. Assur. 2015, 20, 75–83. [Google Scholar] [CrossRef]
- National Public Health Organization-Greece (EODY) Information on the Results of the Genomic Surveillance Network for SARS-CoV-2 Mutations. Available online: https://eody.gov.gr/enimerosi-schetika-me-ta-apotelesmata-diktyoy-gonidiomatikis-epitirisis-gia-tis-metallaxeis-toy-SARS-CoV-2/#main-container (accessed on 18 August 2021). (In Greek)
- Chaintoutis, S.C.; Thomou, Z.; Mouchtaropoulou, E.; Tsiolas, G.; Chassalevris, T.; Stylianaki, I.; Lagou, M.; Michailidou, S.; Moutou, E.; Koenen, J.J.H.; et al. Outbreaks of SARS-CoV-2 in naturally infected mink farms: Impact, transmission dynamics, genetic patterns, and environmental contamination. PLoS Pathog. 2021. [Google Scholar] [CrossRef] [PubMed]
- Rochman, N.D.; Wolf, Y.I.; Faure, G.; Mutz, P.; Zhang, F.; Koonin, E.V. Ongoing global and regional adaptive evolution of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2021, 118, e2104241118. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greaney, A.J.; Loes, A.N.; Crawford, K.H.D.; Starr, T.N.; Malone, K.D.; Chu, H.Y.; Bloom, J.D. Comprehensive mapping of mutations in the SARS-CoV-2 receptor-binding domain that affect recognition by polyclonal human plasma antibodies. Cell Host Microbe 2021, 29, 463–476.e6. [Google Scholar] [CrossRef] [PubMed]
- Owczarzy, R.; You, Y.; Groth, C.L.; Tataurov, A.V. Stability and mismatch discrimination of locked nucleic acid-DNA duplexes. Biochemistry 2011, 50, 9352–9367. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide Name (Fluorophore) | Sequence (5′–3′) | Hybridization on Acc. NC_045512 | Tm (°C) | Conc. (μΜ) |
|---|---|---|---|---|
| DSarsUp1 | TGATTTTACAGGCTGCGTTATAGCTTGGA | 22,843–22,871 | 69.0 | 0.20 |
| SARSpDo1 | ACAGTTGCTGGTGCATGTAGAAGT | 23,110–23,133 | 66.5 | 0.20 |
| Probe 478K (HEX) | ACA+AGG+T+T+TGCT+ACC | 22,910–22,924 | 63.1 | 0.40 |
| Probe 484Εw (FAM) * | AA+AACCT+T+C+AACACCA | 23,005–23,019 | 62.9 | 0.12 |
| Probe 484Κ (Tex.Red) * | AA+AACCT+T+T+AA+CACCA | 62.2 | 0.07 | |
| Probe 501Y (Cy5) * | AACACCA+T+A+AGTGGGT | 23,056–23,071 | 62.7 | 0.30 |
| Inh478T | ACA+AGGTGTGCTA+CC/3SpC3/ | 22,910–22,924 | 61.5 | 0.50 |
| Inh501N | ACACCA+T+T+AGTGGGT/3SpC3/ | 23,057–23,071 | 62.7 | 0.30 |
| Panel | Mutation Identification Method | No. | Variant | Fluorescence | |||
|---|---|---|---|---|---|---|---|
| F | H | T | C | ||||
| Negative | N/A | 20 | N/A | - | - | - | - |
| Positive (A) | Obtained prior to the emergence of the target mutations | 20 | WT | + | - | - | - |
| ViroBOAR Spike 1.0 RT-PCR Kit | 4 | WT | + | - | - | - | |
| 21 | Alpha | + | - | - | + | ||
| 1 | Beta | - | - | + | + | ||
| SNPsig SARS-CoV-2 (EscapePLEX) | 14 | Delta | + | + | - | - | |
| Positive (B) | NGS (Illumina MiSeq) | 2 | Alpha | + | - | - | + |
| 2 | Beta | - | - | + | + | ||
| 36 | Delta | + | + | - | - | ||
| 1 | B.1.1.318 | - | - | + | - | ||
| Negative (Vet) | N/A | 3 cats, 3 minks | N/A | - | - | - | - |
| Positive (Vet) | NGS (Ion Torrent GeneStudio S5) | 2 cats | WT | + | - | - | - |
| NGS (Illumina MiSeq) | 3 minks | WT | + | - | - | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaintoutis, S.C.; Chassalevris, T.; Balaska, S.; Mouchtaropoulou, E.; Tsiolas, G.; Vlatakis, I.; Tychala, A.; Koutsioulis, D.; Argiriou, A.; Skoura, L.; et al. A Novel Real-Time RT-PCR-Based Methodology for the Preliminary Typing of SARS-CoV-2 Variants, Employing Non-Extendable LNA Oligonucleotides and Three Signature Mutations at the Spike Protein Receptor-Binding Domain. Life 2021, 11, 1015. https://doi.org/10.3390/life11101015
Chaintoutis SC, Chassalevris T, Balaska S, Mouchtaropoulou E, Tsiolas G, Vlatakis I, Tychala A, Koutsioulis D, Argiriou A, Skoura L, et al. A Novel Real-Time RT-PCR-Based Methodology for the Preliminary Typing of SARS-CoV-2 Variants, Employing Non-Extendable LNA Oligonucleotides and Three Signature Mutations at the Spike Protein Receptor-Binding Domain. Life. 2021; 11(10):1015. https://doi.org/10.3390/life11101015
Chicago/Turabian StyleChaintoutis, Serafeim C., Taxiarchis Chassalevris, Sofia Balaska, Evangelia Mouchtaropoulou, George Tsiolas, Ioannis Vlatakis, Areti Tychala, Dimitris Koutsioulis, Anagnostis Argiriou, Lemonia Skoura, and et al. 2021. "A Novel Real-Time RT-PCR-Based Methodology for the Preliminary Typing of SARS-CoV-2 Variants, Employing Non-Extendable LNA Oligonucleotides and Three Signature Mutations at the Spike Protein Receptor-Binding Domain" Life 11, no. 10: 1015. https://doi.org/10.3390/life11101015