Anti-Inflammatory Effects of Adenine Enhance Osteogenesis in the Osteoblast-Like MG-63 Cells

 , ,
, ,
Abstract
:1. Background
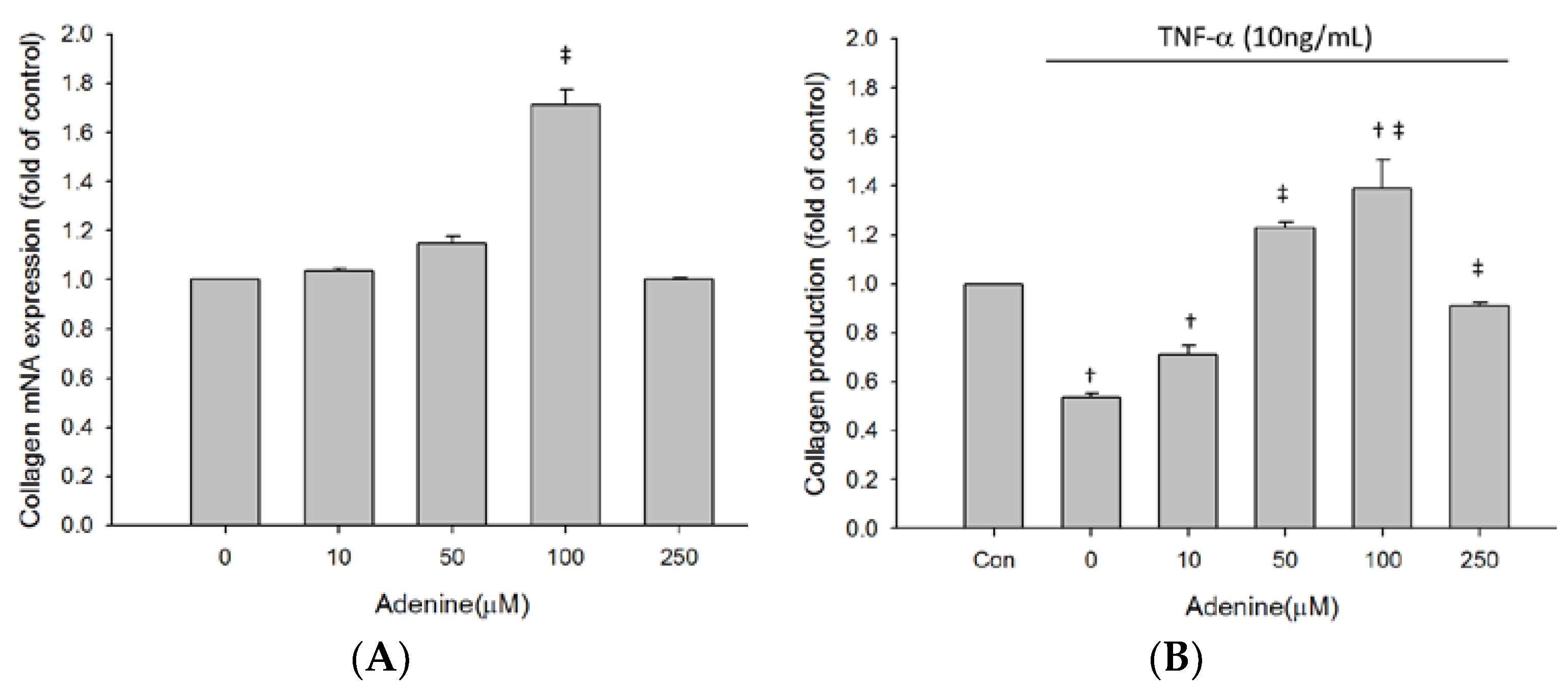
2. Results
2.1. Effects of Adenine on Viability of Osteoblast-Like MG-63 Cells
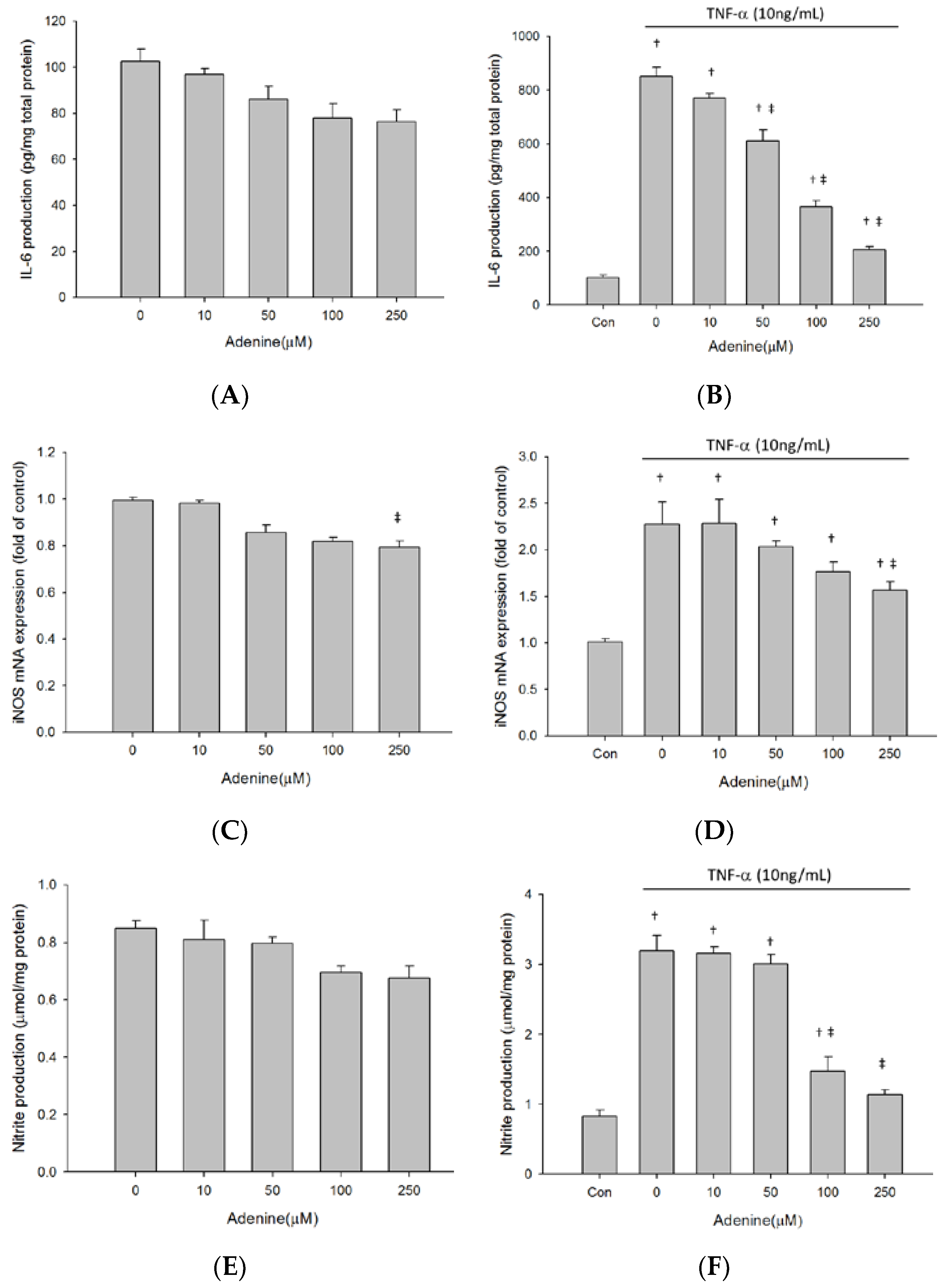
2.2. Adenine Ameliorated TNF-α-Induced IL-6 Production in Osteoblast-Like MG-63 Cells
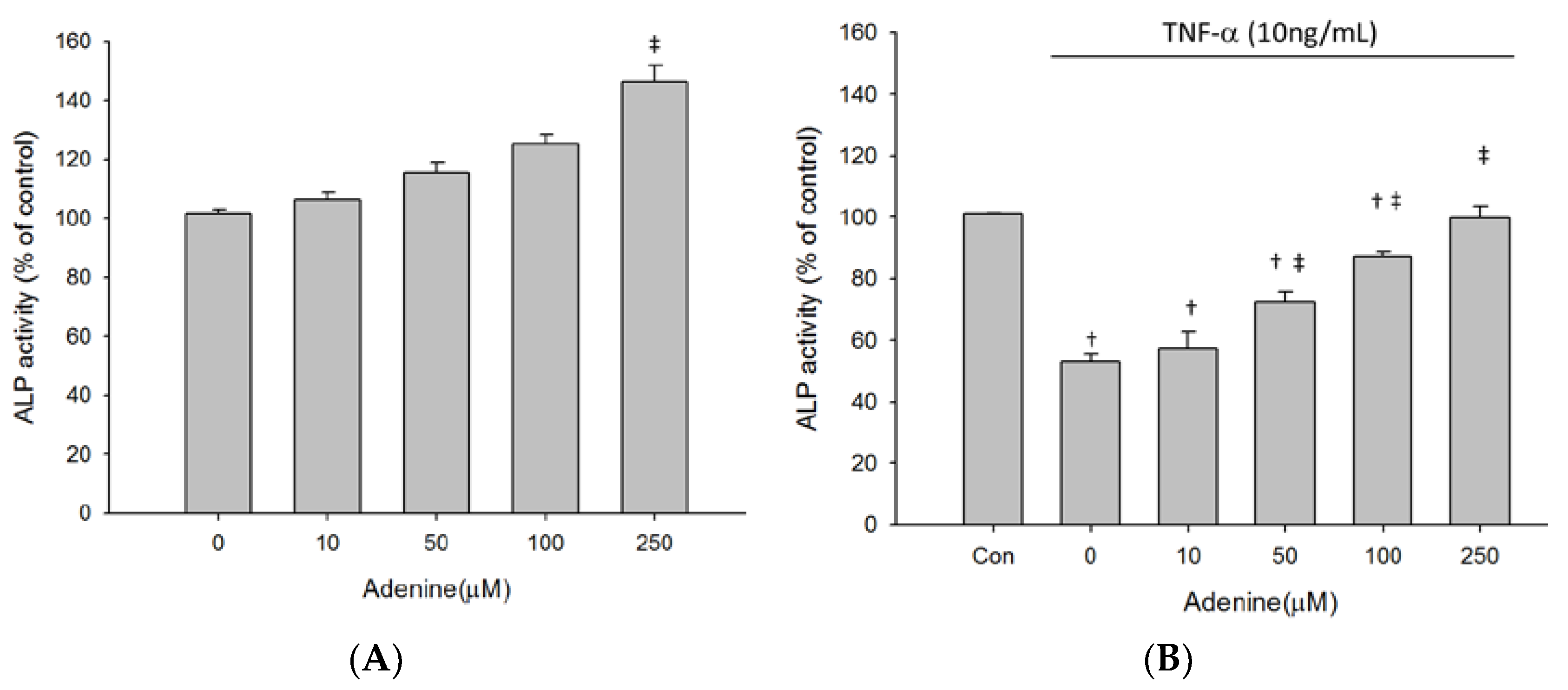
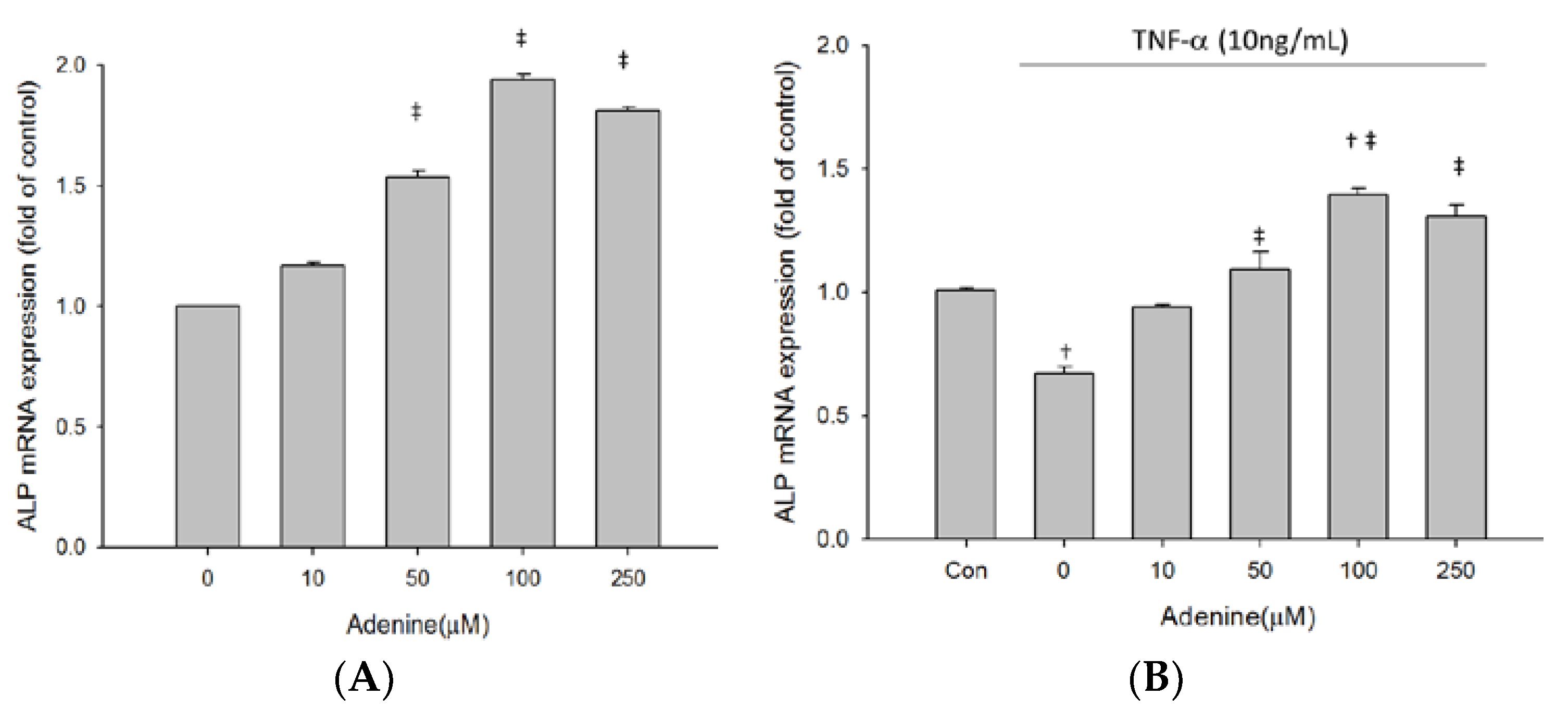
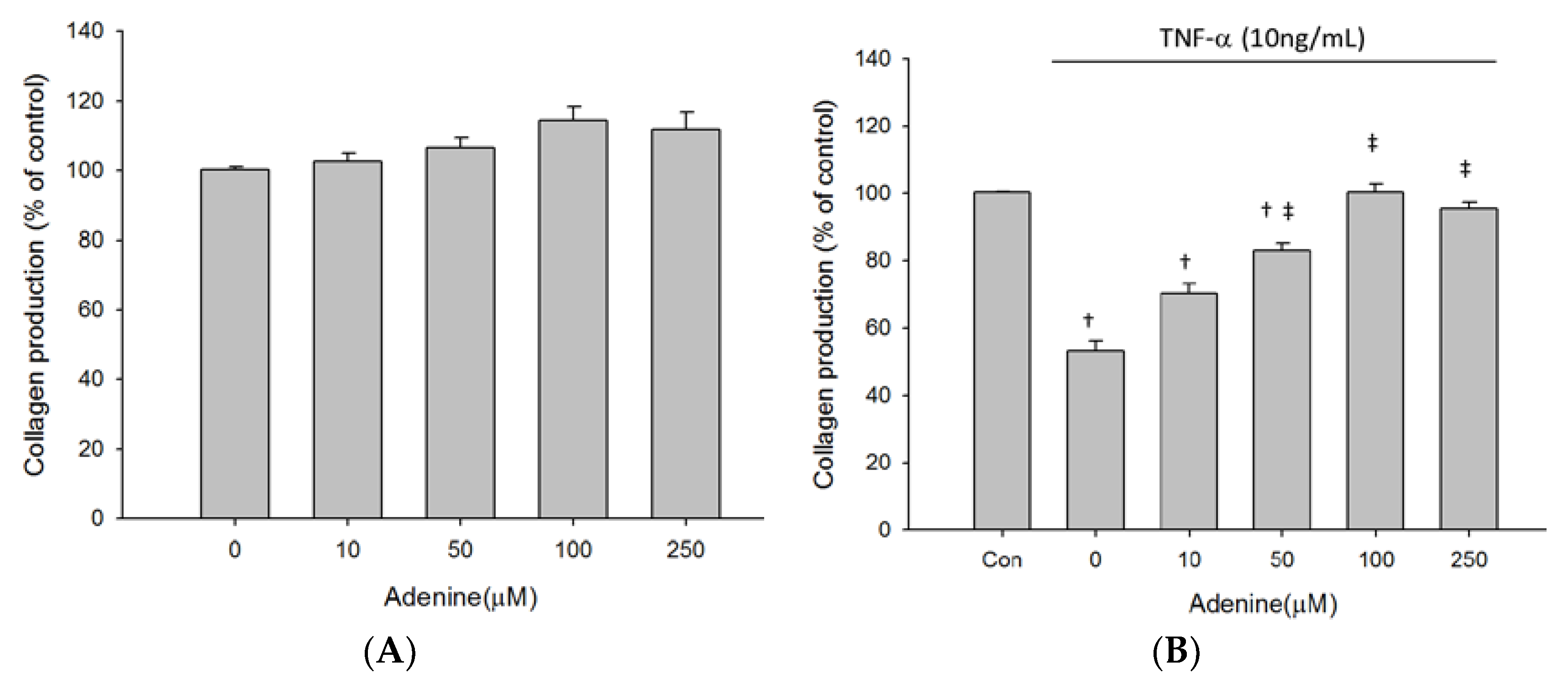
2.3. Effects of Adenine on MG-63 Differentiation
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Viability Assay
4.3. Alkaline Phosphatase Activity Assay
4.4. RNA Extraction, RT-PCR and Quantitative PCR (qPCR)
4.5. ELISA
4.6. Nitric Oxide Assay
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TNF-α | tumor necrosis factor-α |
| ALP | Alkaline phosphatase |
| ATP | adenosine triphosphate |
| DNA | deoxyribonucleic acid |
| NAD | nicotinamide adenine dinucleotide |
| FAD | flavin adenine dinucleotide |
| DMEM | Dulbecco’s modified Eagle’s medium |
| FBS | fetal bovine serum |
| PBS | phosphate-buffered saline |
| ANOVA | one-way analysis of variance |
| iNOS | inducible nitric oxide synthase |
| SLE | systemic lupus erythematosus |
| IBD | inflammatory bowel disease |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide |
| ELISA | enzyme-linked immunosorbent assay |
References
- Johnell, O.; Kanis, J.A. An estimate of the worldwide prevalence and disability associated with osteoporotic fractures. Osteoporos. Int. 2006, 17, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Who are candidates for prevention and treatment for osteoporosis? Osteoporos. Int. 1997, 7, 1–6. [CrossRef] [PubMed]
- Chandran, M.; Lau, T.C.; Gagnon-Arpin, I.; Dobrescu, A.; Li, W.; Leung, M.Y.M.; Patil, N.; Zhao, Z. The health and economic burden of osteoporotic fractures in Singapore and the potential impact of increasing treatment rates through more pharmacological options. Arch. Osteoporos. 2019, 14, 114. [Google Scholar] [CrossRef] [PubMed]
- Borgstrom, F.; Sobocki, P.; Strom, O.; Jonsson, B. The societal burden of osteoporosis in Sweden. Bone 2007, 40, 1602–1609. [Google Scholar] [CrossRef]
- Hernlund, E.; Svedbom, A.; Ivergard, M.; Compston, J.; Cooper, C.; Stenmark, J.; McCloskey, E.V.; Jonsson, B.; Kanis, J.A. Osteoporosis in the European Union: Medical management, epidemiology and economic burden. A report prepared in collaboration with the International Osteoporosis Foundation (IOF) and the European Federation of Pharmaceutical Industry Associations (EFPIA). Arch. Osteoporos. 2013, 8, 136. [Google Scholar] [CrossRef] [Green Version]
- Wimalawansa, S.J. Prevention and treatment of osteoporosis: Efficacy of combination of hormone replacement therapy with other antiresorptive agents. J. Clin. Densitom. 2000, 3, 187–201. [Google Scholar] [CrossRef]
- Schonberg, M.A.; Davis, R.B.; Wee, C.C. After the Women’s Health Initiative: Decision making and trust of women taking hormone therapy. Women’s Health Issues 2005, 15, 187–195. [Google Scholar] [CrossRef]
- Green, J.; Czanner, G.; Reeves, G.; Watson, J.; Wise, L.; Beral, V. Oral bisphosphonates and risk of cancer of oesophagus, stomach, and colorectum: Case-control analysis within a UK primary care cohort. BMJ 2010, 341, c4444. [Google Scholar] [CrossRef] [Green Version]
- Ginaldi, L.; Di Benedetto, M.C.; De Martinis, M. Osteoporosis, inflammation and ageing. Immun. Ageing 2005, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Boyce, B.F.; Li, J.; Xing, L.; Yao, Z. Bone Remodeling and the Role of TRAF3 in Osteoclastic Bone Resorption. Front. Immunol. 2018, 9, 2263. [Google Scholar] [CrossRef]
- Boyce, B.F.; Li, P.; Yao, Z.; Zhang, Q.; Badell, I.R.; Schwarz, E.M.; O’Keefe, R.J.; Xing, L. TNF-alpha and pathologic bone resorption. Keio J. Med. 2005, 54, 127–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaschke, M.; Koepp, R.; Cortis, J.; Komrakova, M.; Schieker, M.; Hempel, U.; Siggelkow, H. IL-6, IL-1beta, and TNF-alpha only in combination influence the osteoporotic phenotype in Crohn’s patients via bone formation and bone resorption. Adv. Clin. Exp. Med. 2018, 27, 45–56. [Google Scholar] [CrossRef]
- Grassi, F.; Tell, G.; Robbie-Ryan, M.; Gao, Y.; Terauchi, M.; Yang, X.; Romanello, M.; Jones, D.P.; Weitzmann, M.N.; Pacifici, R. Oxidative stress causes bone loss in estrogen-deficient mice through enhanced bone marrow dendritic cell activation. Proc. Natl. Acad. Sci. USA 2007, 104, 15087–15092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; He, H.; Gao, C.; Luo, G.; Xie, Y.; Wang, H.; Tian, L.; Chen, X.; Yu, X.; He, C. Ovariectomy-induced bone loss in TNFalpha and IL6 gene knockout mice is regulated by different mechanisms. J. Mol. Endocrinol. 2018, 60, 185–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidney, L.E.; Kirkham, G.R.; Buttery, L.D. Comparison of osteogenic differentiation of embryonic stem cells and primary osteoblasts revealed by responses to IL-1beta, TNF-alpha, and IFN-gamma. Stem Cells Dev. 2014, 23, 605–617. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.Z.; Yang, X.; Bi, Z.G. The effects of 6-gingerol on proliferation, differentiation, and maturation of osteoblast-like MG-63 cells. Braz. J. Med. Biol. Res. 2015, 48, 637–643. [Google Scholar] [CrossRef] [Green Version]
- Centrella, M.; McCarthy, T.L.; Canalis, E. Tumor necrosis factor-alpha inhibits collagen synthesis and alkaline phosphatase activity independently of its effect on deoxyribonucleic acid synthesis in osteoblast-enriched bone cell cultures. Endocrinology 1988, 123, 1442–1448. [Google Scholar] [CrossRef]
- Nazrun, A.S.; Norazlina, M.; Norliza, M.; Nirwana, S.I. The anti-inflammatory role of vitamin e in prevention of osteoporosis. Adv. Pharmacol. Sci. 2012, 2012, 142702. [Google Scholar] [CrossRef]
- O’Connor, J.P.; Lysz, T. Celecoxib, NSAIDs and the skeleton. Drugs Today 2008, 44, 693–709. [Google Scholar] [CrossRef]
- Rao, S.S.; Hu, Y.; Xie, P.L.; Cao, J.; Wang, Z.X.; Liu, J.H.; Yin, H.; Huang, J.; Tan, Y.J.; Luo, J.; et al. Omentin-1 prevents inflammation-induced osteoporosis by downregulating the pro-inflammatory cytokines. Bone Res. 2018, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Yao, Z.; Lei, W.; Duan, R.; Li, Y.; Luo, L.; Boyce, B.F. RANKL cytokine enhances TNF-induced osteoclastogenesis independently of TNF receptor associated factor (TRAF) 6 by degrading TRAF3 in osteoclast precursors. J. Biol. Chem. 2017, 292, 10169–10179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosoi, T.; Ino, S.; Ohnishi, F.; Todoroki, K.; Yoshii, M.; Kakimoto, M.; Muller, C.E.; Ozawa, K. Mechanisms of the action of adenine on anti-allergic effects in mast cells. Immun. Inflamm. Dis. 2018, 6, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Silwal, P.; Shin, K.; Choi, S.; Kang, S.W.; Park, J.B.; Lee, H.J.; Koo, S.J.; Chung, K.H.; Namgung, U.; Lim, K.; et al. Adenine suppresses IgE-mediated mast cell activation. Mol. Immunol. 2015, 65, 242–249. [Google Scholar] [CrossRef]
- Scioli, M.G.; Bielli, A.; Agostinelli, S.; Tarquini, C.; Arcuri, G.; Ferlosio, A.; Costanza, G.; Doldo, E.; Orlandi, A. Antioxidant treatment prevents serum deprivation- and TNF-alpha-induced endothelial dysfunction through the inhibition of NADPH oxidase 4 and the restoration of beta-oxidation. J. Vasc. Res. 2014, 51, 327–337. [Google Scholar] [CrossRef]
- Silwal, P.; Lim, K.; Heo, J.Y.; Park, J.I.; Namgung, U.; Park, S.K. Adenine attenuates lipopolysaccharide-induced inflammatory reactions. Korean J. Physiol. Pharmacol. 2018, 22, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, S.; Yoshimi, Y.; Ikekita, M. Neuroprotective effect of adenine on purkinje cell survival in rat cerebellar primary cultures. J. Neurosci. Res. 2003, 74, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Yoshimi, Y.; Watanabe, S.; Shinomiya, T.; Makino, A.; Toyoda, M.; Ikekita, M. Nucleobase adenine as a trophic factor acting on Purkinje cells. Brain Res. 2003, 991, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Luthje, J. Extracellular adenine compounds, red blood cells and haemostasis: Facts and hypotheses. Blut 1989, 59, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Saribal, D.; Hocaoglu-Emre, F.S.; Erdogan, S.; Bahtiyar, N.; Caglar Okur, S.; Mert, M. Inflammatory cytokines IL-6 and TNF-alpha in patients with hip fracture. Osteoporos. Int. 2019, 30, 1025–1031. [Google Scholar] [CrossRef]
- Kastelan, D.; Kastelan, M.; Massari, L.P.; Korsic, M. Possible association of psoriasis and reduced bone mineral density due to increased TNF-alpha and IL-6 concentrations. Med. hypotheses 2006, 67, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.Y.; Baek, K.H.; Rhee, E.J.; Tae, H.J.; Oh, K.W.; Kang, M.I.; Lee, K.W.; Kim, S.W.; Kim, C.C.; Oh, E.S. Impact of circulating bone-resorbing cytokines on the subsequent bone loss following bone marrow transplantation. Bone Marrow Transplant. 2004, 34, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, S.J.; McPherson, J.R.; Pahan, K.; Koka, S. Regulation of TNF-alpha-induced IL-6 production in MG-63 human osteoblast-like cells. J. Dent. Res. 2002, 81, 17–22. [Google Scholar] [PubMed]
- Confalone, E.; D’Alessio, G.; Furia, A. IL-6 Induction by TNFalpha and IL-1beta in an Osteoblast-Like Cell Line. Int. J. Biomed. Sci. 2010, 6, 135–140. [Google Scholar] [PubMed]
- Abdel Meguid, M.H.; Hamad, Y.H.; Swilam, R.S.; Barakat, M.S. Relation of interleukin-6 in rheumatoid arthritis patients to systemic bone loss and structural bone damage. Rheumatol. Int. 2013, 33, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Kriel, M.; Sayers, A.; Fraser, W.D.; Williams, A.M.; Koch, A.; Zacharowski, K.; Probert, C.S.; Tobias, J.H. IL-6 may modulate the skeletal response to glucocorticoids during exacerbations of inflammatory bowel disease. Calcif. Tissue Int. 2010, 86, 375–381. [Google Scholar] [CrossRef] [PubMed]
- de Vries, T.J.; Yousovich, J.; Schoenmaker, T.; Scheres, N.; Everts, V. Tumor necrosis factor-alpha antagonist infliximab inhibits osteoclast formation of peripheral blood mononuclear cells but does not affect periodontal ligament fibroblast-mediated osteoclast formation. J. Periodontal Res. 2016, 51, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Timmen, M.; Hidding, H.; Wieskotter, B.; Baum, W.; Pap, T.; Raschke, M.J.; Schett, G.; Zwerina, J.; Stange, R. Influence of antiTNF-alpha antibody treatment on fracture healing under chronic inflammation. BMC Musculoskelet. Disord. 2014, 15, 184. [Google Scholar] [CrossRef] [Green Version]
- Musacchio, E.; Valvason, C.; Botsios, C.; Ostuni, F.; Furlan, A.; Ramonda, R.; Modesti, V.; Sartori, L.; Punzi, L. The tumor necrosis factor-{alpha}-blocking agent infliximab inhibits interleukin 1beta (IL-1beta) and IL-6 gene expression in human osteoblastic cells. J. Rheumatol. 2009, 36, 1575–1579. [Google Scholar] [CrossRef]
- Lin, S.K.; Kok, S.H.; Lin, L.D.; Wang, C.C.; Kuo, M.Y.; Lin, C.T.; Hsiao, M.; Hong, C.Y. Nitric oxide promotes the progression of periapical lesion via inducing macrophage and osteoblast apoptosis. Oral Microbiol. Immunol. 2007, 22, 24–29. [Google Scholar] [CrossRef]
- Hukkanen, M.; Hughes, F.J.; Buttery, L.D.; Gross, S.S.; Evans, T.J.; Seddon, S.; Riveros-Moreno, V.; Macintyre, I.; Polak, J.M. Cytokine-stimulated expression of inducible nitric oxide synthase by mouse, rat, and human osteoblast-like cells and its functional role in osteoblast metabolic activity. Endocrinology 1995, 136, 5445–5453. [Google Scholar] [CrossRef]
- Damoulis, P.D.; Hauschka, P.V. Nitric oxide acts in conjunction with proinflammatory cytokines to promote cell death in osteoblasts. J. Bone Miner. Res. 1997, 12, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Kalajzic, I.; Staal, A.; Yang, W.P.; Wu, Y.; Johnson, S.E.; Feyen, J.H.; Krueger, W.; Maye, P.; Yu, F.; Zhao, Y.; et al. Expression profile of osteoblast lineage at defined stages of differentiation. J. Biol. Chem. 2005, 280, 24618–24626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaturvedi, R.; Singha, P.K.; Dey, S. Water soluble bioactives of nacre mediate antioxidant activity and osteoblast differentiation. PloS ONE 2013, 8, e84584. [Google Scholar] [CrossRef]
- Choi, J.Y.; Lee, B.H.; Song, K.B.; Park, R.W.; Kim, I.S.; Sohn, K.Y.; Jo, J.S.; Ryoo, H.M. Expression patterns of bone-related proteins during osteoblastic differentiation in MC3T3-E1 cells. J. Cell. Biochem. 1996, 61, 609–618. [Google Scholar] [CrossRef]
- Manduca, P.; Sanguineti, C.; Pistone, M.; Boccignone, E.; Sanguineti, F.; Santolini, F.; Federici, A. Differential expression of alkaline phosphatase in clones of human osteoblast-like cells. J. Bone Miner. Res. 1993, 8, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Gerstenfeld, L.C.; Chipman, S.D.; Kelly, C.M.; Hodgens, K.J.; Lee, D.D.; Landis, W.J. Collagen expression, ultrastructural assembly, and mineralization in cultures of chicken embryo osteoblasts. J. Cell Biol. 1988, 106, 979–989. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IL-6 | 5′-TAGCCCTGAGAAAGGAGACATG-3′ | 5′-AGGCAAGTCTCCTCATTGAATC-3′ |
| iNOS | 5′-GCGGAGCGATGGGAAGCATG-3′ | 5′-CCCGAGCTCCTGGAACCAC-3′ |
| Alkaline phosphatase | 5’-CATATCGTGTCCAAACTCAGT-3’ | 5’-ATAAACCCCCTGTGAAGTTGCA-3’ |
| Collagen Type I | 5’-AACAGGAAGGGCCACGACAA-3’ | 5’-GCGGCACAAGGGATTGAC-3’ |
| β-actin | 5’-TGACGTGGACATCCGCAAAG-3’ | 5’-CTGGAAGGTGGACAGCGAGG-3’ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-P.; Chu, Y.-L.; Tsuang, Y.-H.; Wu, Y.; Kuo, C.-Y.; Kuo, Y.-J. Anti-Inflammatory Effects of Adenine Enhance Osteogenesis in the Osteoblast-Like MG-63 Cells. Life 2020, 10, 116. https://doi.org/10.3390/life10070116
Chen Y-P, Chu Y-L, Tsuang Y-H, Wu Y, Kuo C-Y, Kuo Y-J. Anti-Inflammatory Effects of Adenine Enhance Osteogenesis in the Osteoblast-Like MG-63 Cells. Life. 2020; 10(7):116. https://doi.org/10.3390/life10070116
Chicago/Turabian StyleChen, Yu-Pin, Yo-Lun Chu, Yang-Hwei Tsuang, Yueh Wu, Cheng-Yi Kuo, and Yi-Jie Kuo. 2020. "Anti-Inflammatory Effects of Adenine Enhance Osteogenesis in the Osteoblast-Like MG-63 Cells" Life 10, no. 7: 116. https://doi.org/10.3390/life10070116