Carotenoid Raman Signatures Are Better Preserved in Dried Cells of the Desert Cyanobacterium Chroococcidiopsis than in Hydrated Counterparts after High-Dose Gamma Irradiation

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cyanobacterial Strain and Culture Conditions
2.2. Sample Preparation and Irradiation
2.3. Raman Spectroscopy: Set-up and Spectra Parameters
2.4. Data Processing
2.5. Bioinformatics Analysis
3. Results
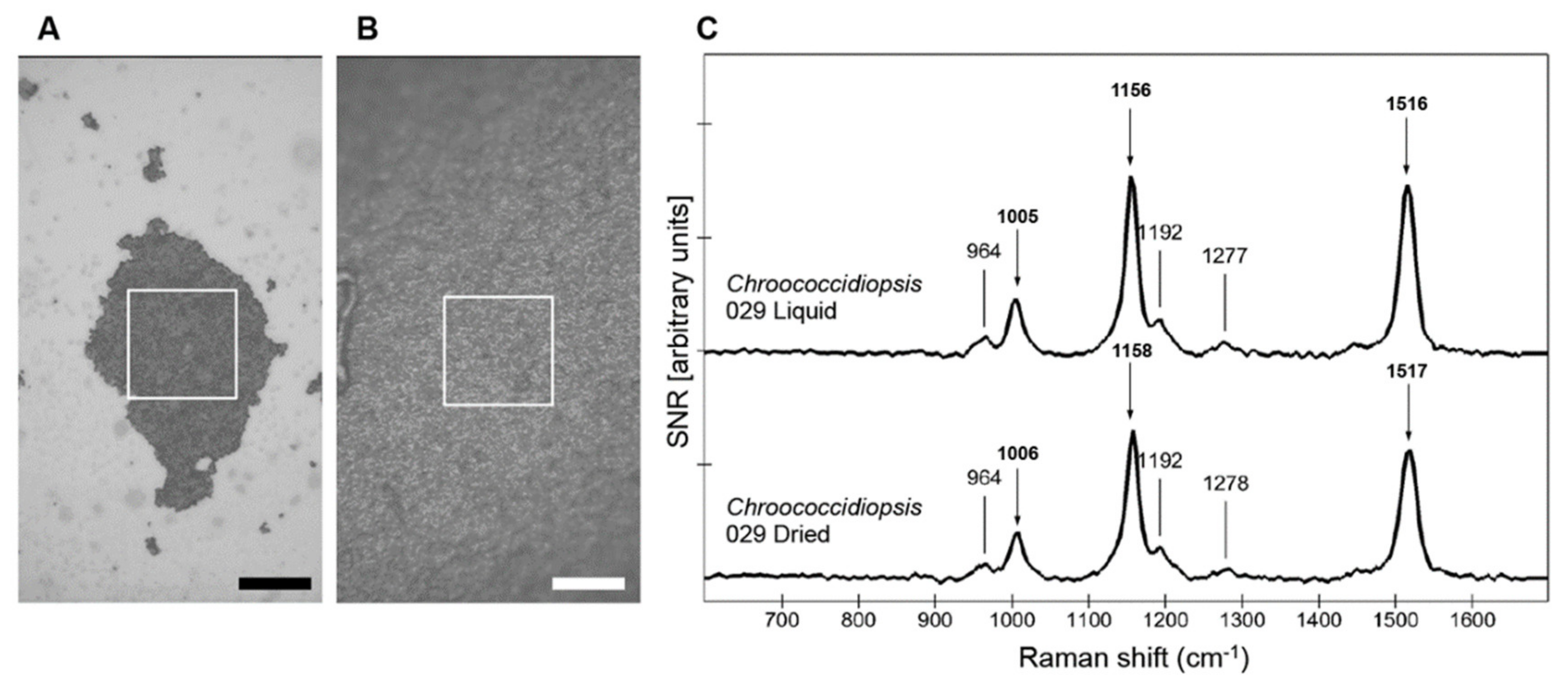
3.1. Comparison of Carotenoid Raman Signal in Dried and Hydrated Cells
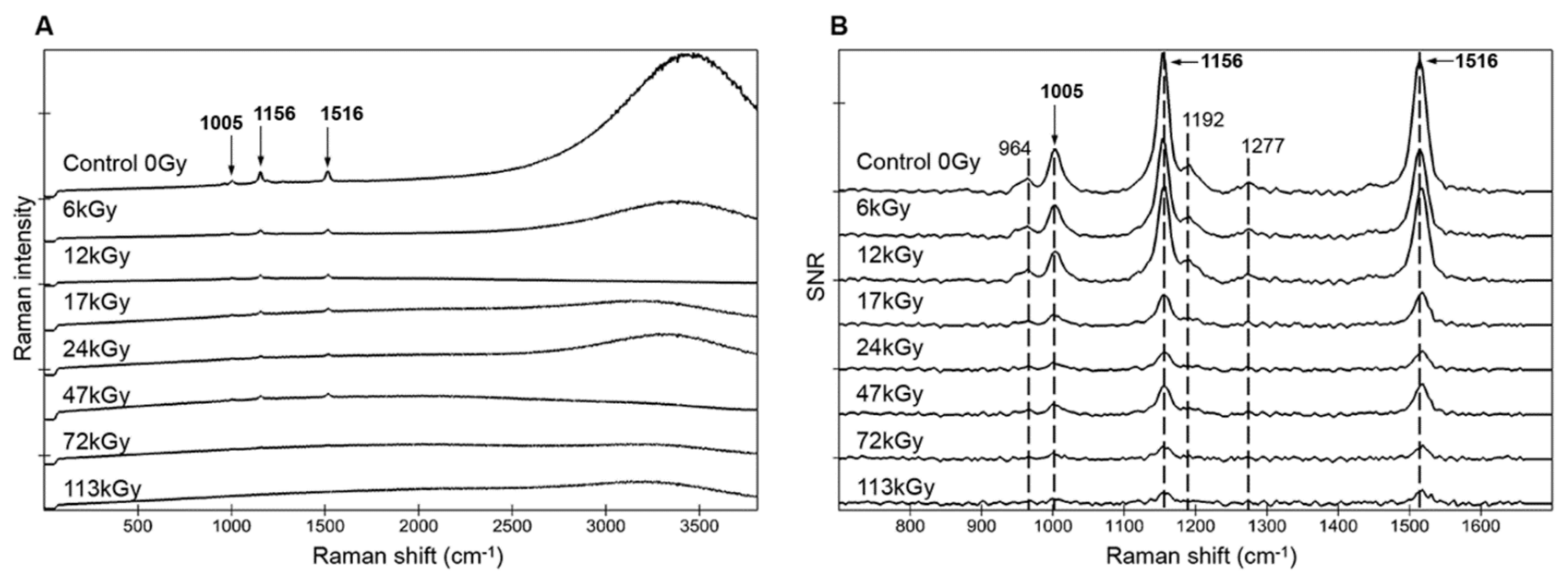
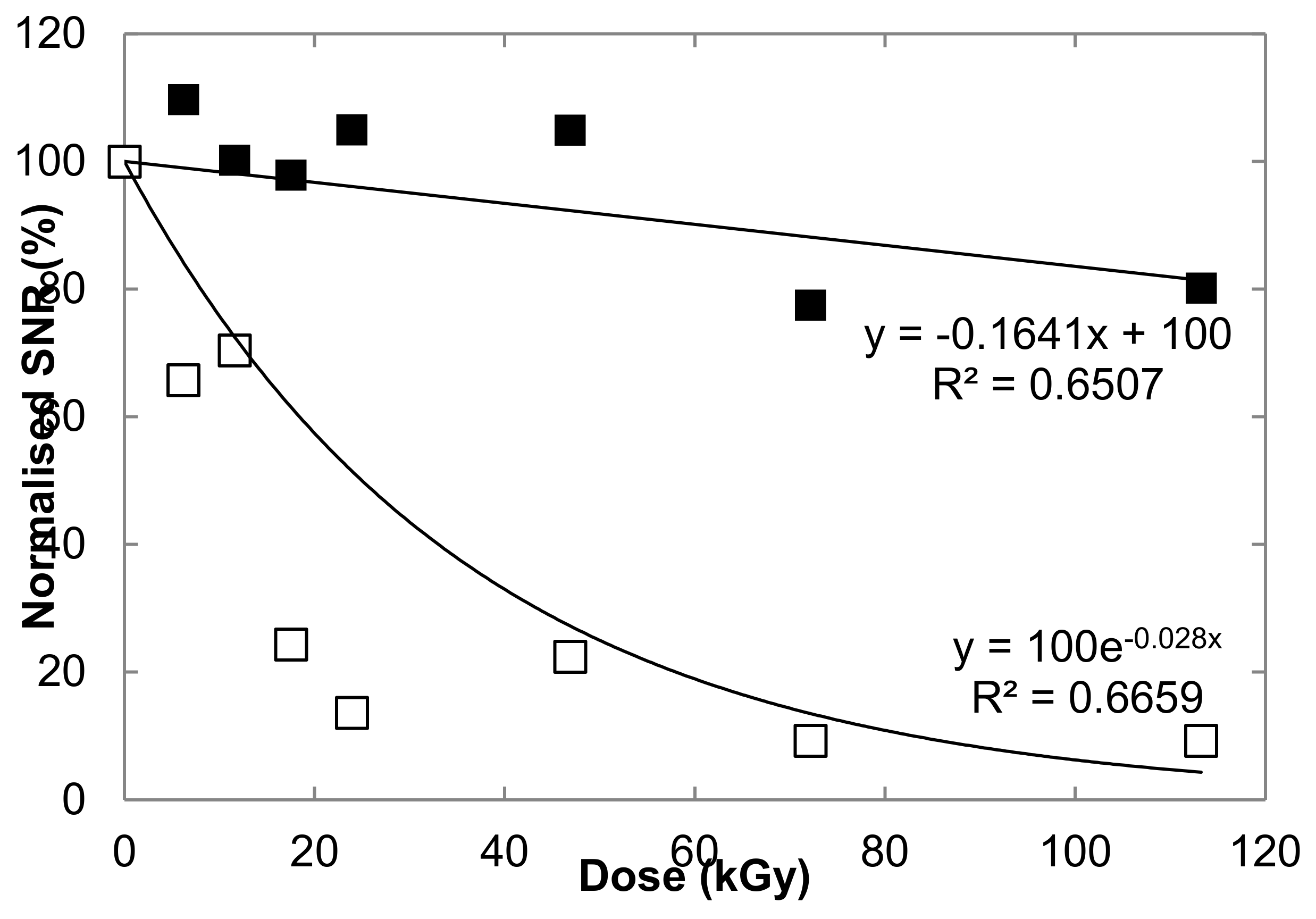
3.2. Degradation of Carotenoid Raman Signatures in Hydrated Cells
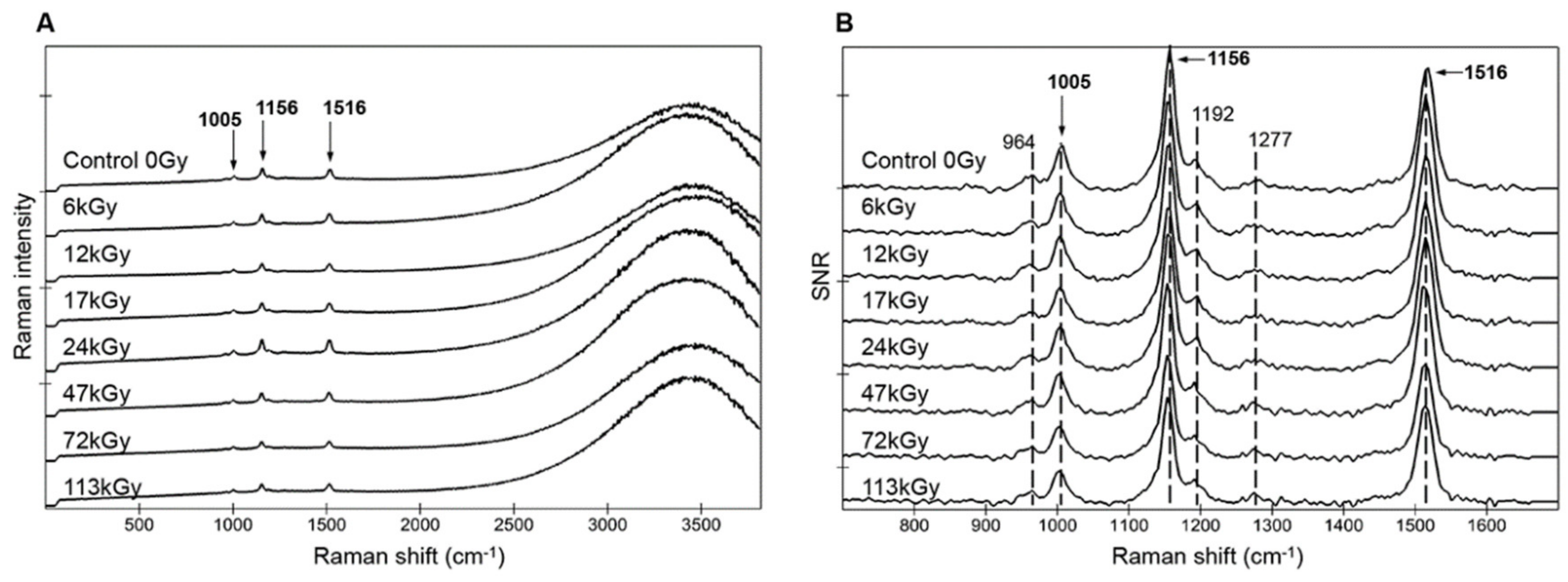
3.3. Preservation of Carotenoid Raman Signal in Dried Cells
3.4. Identification of Carotenogenesis Genes
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lasne, J.; Noblet, A.; Szopa, C.; Navarro-González, R.; Cabane, M.; Poch, O.; Stalport, F.; François, P.; Atreya, S.K.; Coll, P. Oxidants at the Surface of Mars: A Review in Light of Recent Exploration Results. Astrobiology 2016, 16, 977–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassler, D.M.; Zeitlin, C.; Wimmer-Schweingruber, R.F.; Ehresmann, B.; Rafkin, S.; Eigenbrode, J.L.; Brinza, D.E.; Weigle, G.; Böttcher, S.; Böhm, E.; et al. Mars’ Surface Radiation Environment Measured with the Mars Science Laboratory’s Curiosity Rover. Science 2014, 343, 1244797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlov, A.A.; Vasilyev, G.; Ostryakov, V.M.; Pavlov, A.K.; Mahaffy, P. Degradation of the organic molecules in the shallow subsurface of Mars due to irradiation by cosmic rays. Geophys. Res. Lett. 2012, 39, L13202. [Google Scholar] [CrossRef] [Green Version]
- Neveu, M.; Hays, L.E.; Voytek, M.A.; New, M.H.; Schulte, M.D. The Ladder of Life Detection. Astrobiology 2018. [Google Scholar] [CrossRef] [Green Version]
- Pace, N.R. The universal nature of biochemistry. Proc. Natl. Acad. Sci. USA 2001, 98, 805–808. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, G.A. Eubacteria show their true colors: Genetics of carotenoid pigment biosynthesis from microbes to plants. J. Bacteriol. 1994, 176, 4795–4802. [Google Scholar] [CrossRef] [Green Version]
- Alcaíno, J.; Baeza, M.; Cifuentes, V. Carotenoid Distribution in Nature. In Carotenoids in Nature; Stange, C., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 79, pp. 3–33. ISBN 978-3-319-39124-3. [Google Scholar]
- Parnell, J.; Cullen, D.; Sims, M.R.; Bowden, S.; Cockell, C.S.; Court, R.; Ehrenfreund, P.; Gaubert, F.; Grant, W.; Parro, V.; et al. Searching for Life on Mars: Selection of Molecular Targets for ESA’s Aurora ExoMars Mission. Astrobiology 2007, 7, 578–604. [Google Scholar] [CrossRef]
- Rull, F.; Maurice, S.; Hutchinson, I.; Moral, A.; Perez, C.; Diaz, C.; Colombo, M.; Belenguer, T.; Lopez-Reyes, G.; Sansano, A.; et al. The Raman Laser Spectrometer for the ExoMars Rover Mission to Mars. Astrobiology 2017, 17, 627–654. [Google Scholar] [CrossRef] [Green Version]
- Wiens, R.C.; Maurice, S.; Mccabe, K.; Cais, P.; Anderson, R.B.; Beyssac, O.; Bonal, L.; Clegg, S.; Deflores, L.; Dromart, G.; et al. The SuperCam Remote Sensing Instrument Suite for Mars 2020. In Proceedings of the 47th Lunar and Planetary Science Conference, The Woodlands, TX, USA, 21–25 March 2016. [Google Scholar]
- Beegle, L.; Bhartia, R.; White, M.; DeFlores, L.; Abbey, W.; Wu, Y.-H.; Cameron, B.; Moore, J.; Fries, M.; Burton, A.; et al. SHERLOC: Scanning habitable environments with Raman amp; luminescence for organics amp; chemicals. In Proceedings of the 2015 IEEE Aerospace Conference, Big Sky, MT, USA, 7–14 March 2015. [Google Scholar]
- Cottin, H.; Kotler, J.M.; Billi, D.; Cockell, C.; Demets, R.; Ehrenfreund, P.; Elsaesser, A.; d’Hendecourt, L.; Loon, J.J.W.A.; van Martins, Z.; et al. Space as a Tool for Astrobiology: Review and Recommendations for Experimentations in Earth Orbit and Beyond. Space Sci. Rev. 2017, 209, 83–181. [Google Scholar] [CrossRef]
- Martins, Z.; Cottin, H.; Kotler, J.M.; Carrasco, N.; Cockell, C.S.; de la Noetzel, R.T.; Demets, R.; Vera, J.-P.; de d’Hendecourt, L.; Ehrenfreund, P.; et al. Earth as a Tool for Astrobiology—A European Perspective. Space Sci. Rev. 2017, 1–39. [Google Scholar] [CrossRef]
- Verseux, C.; Baqué, M.; Cifariello, R.; Fagliarone, C.; Raguse, M.; Moeller, R.; Billi, D. Evaluation of the Resistance of Chroococcidiopsis spp. to Sparsely and Densely Ionizing Irradiation. Astrobiology 2017, 17, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Billi, D.; Verseux, C.; Fagliarone, C.; Napoli, A.; Baqué, M.; de Vera, J.-P. A Desert Cyanobacterium under Simulated Mars-like Conditions in Low Earth Orbit: Implications for the Habitability of Mars. Astrobiology 2019, 19, 158–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horneck, G.; Klaus, D.M.; Mancinelli, R.L. Space Microbiology. Microbiol. Mol. Biol. Rev. MMBR 2010, 74, 121–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertem, G.; Ertem, M.C.; McKay, C.P.; Hazen, R.M. Shielding biomolecules from effects of radiation by Mars analogue minerals and soils. Int. J. Astrobiol. 2017, 16, 280–285. [Google Scholar] [CrossRef]
- Baqué, M.; Verseux, C.; Böttger, U.; Rabbow, E.; de Vera, J.-P.; Billi, D. Preservation of Biomarkers from Cyanobacteria Mixed with Mars Like Regolith Under Simulated Martian Atmosphere and UV Flux. Orig. Life Evol. Biosph. 2015, 46, 289–310. [Google Scholar] [CrossRef]
- Cockell, C.S.; Rettberg, P.; Rabbow, E.; Olsson-Francis, K. Exposure of phototrophs to 548 days in low Earth orbit: Microbial selection pressures in outer space and on early earth. ISME J. 2011, 5, 1671–1682. [Google Scholar] [CrossRef] [Green Version]
- Bryce, C.C.; Horneck, G.; Rabbow, E.; Edwards, H.G.M.; Cockell, C.S. Impact shocked rocks as protective habitats on an anoxic early Earth. Int. J. Astrobiol. 2015, 14, 115–122. [Google Scholar] [CrossRef] [Green Version]
- de Vera, J.-P.; Alawi, M.; Backhaus, T.; Baqué, M.; Billi, D.; Böttger, U.; Berger, T.; Bohmeier, M.; Cockell, C.; Demets, R.; et al. Limits of Life and the Habitability of Mars: The ESA Space Experiment BIOMEX on the ISS. Astrobiology 2019, 19, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Dachev, T.P.; Bankov, N.G.; Tomov, B.T.; Matviichuk, Y.N.; Dimitrov, P.G.; Häder, D.-P.; Horneck, G. Overview of the ISS Radiation Environment Observed during the ESA EXPOSE-R2 Mission in 2014–2016. Space Weather 2017, 15, 2016SW001580. [Google Scholar] [CrossRef]
- Dartnell, L.R.; Desorgher, L.; Ward, J.M.; Coates, A.J. Martian sub-surface ionising radiation: Biosignatures and geology. Biogeosciences 2007, 4, 545–558. [Google Scholar] [CrossRef] [Green Version]
- Moeller, R.; Raguse, M.; Leuko, S.; Berger, T.; Hellweg, C.E.; Fujimori, A.; Okayasu, R.; Horneck, G. STARLIFE—An International Campaign to Study the Role of Galactic Cosmic Radiation in Astrobiological Model Systems. Astrobiology 2017, 17, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Meeßen, J.; Backhaus, T.; Brandt, A.; Raguse, M.; Böttger, U.; de Vera, J.-P.; de la Torre, R. The Effect of High-Dose Ionizing Radiation on the Isolated Photobiont of the Astrobiological Model Lichen Circinaria gyrosa. Astrobiology 2017, 17, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Baqué, M.; Hanke, F.; Böttger, U.; Leya, T.; Moeller, R.; Vera, J.-P. de Protection of cyanobacterial carotenoids’ Raman signatures by Martian mineral analogues after high-dose gamma irradiation. J. Raman Spectrosc. 2018, 49, 1617–1627. [Google Scholar] [CrossRef]
- Vago, J.L.; Westall, F.; Pasteur Instrument Teams, L.S.S.W.G.; Other Contributors; Coates, A.J.; Jaumann, R.; Korablev, O.; Ciarletti, V.; Mitrofanov, I.; Josset, J.-L.; et al. Habitability on Early Mars and the Search for Biosignatures with the ExoMars Rover. Astrobiology 2017, 17, 471–510. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinforma. Oxf. Engl. 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Cuccuru, G.; Orsini, M.; Pinna, A.; Sbardellati, A.; Soranzo, N.; Travaglione, A.; Uva, P.; Zanetti, G.; Fotia, G. Orione, a web-based framework for NGS analysis in microbiology. Bioinformatics 2014, 30, 1928–1929. [Google Scholar] [CrossRef] [Green Version]
- Vítek, P.; Jehlička, J.; Edwards, H.G.M.; Osterrothová, K. Identification of β-carotene in an evaporitic matrix—Evaluation of Raman spectroscopic analysis for astrobiological research on Mars. Anal. Bioanal. Chem. 2009, 393, 1967–1975. [Google Scholar] [CrossRef]
- Hooijschuur, J.-H.; Verkaaik, M.F.C.; Davies, G.R.; Ariese, F. Raman spectroscopy for future planetary exploration: Photodegradation, self-absorption and quantification of carotenoids in microorganisms and mineral matrices. J. Raman Spectrosc. 2015, 46, 856–862. [Google Scholar] [CrossRef] [Green Version]
- Sebben, J.A.; da Silveira Espindola, J.; Ranzan, L.; Fernandes de Moura, N.; Trierweiler, L.F.; Trierweiler, J.O. Development of a quantitative approach using Raman spectroscopy for carotenoids determination in processed sweet potato. Food Chem. 2018, 245, 1224–1231. [Google Scholar] [CrossRef]
- Hershkovitz, N.; Oren, A.; Cohen, Y. Accumulation of Trehalose and Sucrose in Cyanobacteria Exposed to Matric Water Stress. Appl. Environ. Microbiol. 1991, 57, 645–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagliarone, C.; Mosca, C.; Ubaldi, I.; Verseux, C.; Baqué, M.; Wilmotte, A.; Billi, D. Avoidance of protein oxidation correlates with the desiccation and radiation resistance of hot and cold desert strains of the cyanobacterium Chroococcidiopsis. Extremophiles 2017, 21, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Dartnell, L.R.; Page, K.; Jorge-Villar, S.E.; Wright, G.; Munshi, T.; Scowen, I.J.; Ward, J.M.; Edwards, H.G.M. Destruction of Raman biosignatures by ionising radiation and the implications for life detection on Mars. Anal. Bioanal. Chem. 2012, 403, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Edwards, H.G.M.; Moody, C.D.; Jorge Villar, S.E.; Wynn-Williams, D.D. Raman spectroscopic detection of key biomarkers of cyanobacteria and lichen symbiosis in extreme Antarctic habitats: Evaluation for Mars Lander missions. Icarus 2005, 174, 560–571. [Google Scholar] [CrossRef]
- Domonkos, I.; Kis, M.; Gombos, Z.; Ughy, B. Carotenoids, versatile components of oxygenic photosynthesis. Prog. Lipid Res. 2013, 52, 539–561. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S.; Mochimaru, M. Carotenoids and carotenogenesis in cyanobacteria: Unique ketocarotenoids and carotenoid glycosides. Cell. Mol. Life Sci. CMLS 2007, 64, 2607–2619. [Google Scholar] [CrossRef]
- Kłodawska, K.; Bujas, A.; Turos-Cabal, M.; Żbik, P.; Fu, P.; Malec, P. Effect of growth temperature on biosynthesis and accumulation of carotenoids in cyanobacterium Anabaena sp. PCC 7120 under diazotrophic conditions. Microbiol. Res. 2019, 226, 34–40. [Google Scholar] [CrossRef]
- Vítek, P.; Ascaso, C.; Artieda, O.; Casero, M.C.; Wierzchos, J. Discovery of carotenoid red-shift in endolithic cyanobacteria from the Atacama Desert. Sci. Rep. 2017, 7, 11116. [Google Scholar] [CrossRef]
- Puente-Sánchez, F.; Arce-Rodríguez, A.; Oggerin, M.; García-Villadangos, M.; Moreno-Paz, M.; Blanco, Y.; Rodríguez, N.; Bird, L.; Lincoln, S.A.; Tornos, F.; et al. Viable cyanobacteria in the deep continental subsurface. Proc. Natl. Acad. Sci. USA 2018, 115, 10702–10707. [Google Scholar] [CrossRef] [Green Version]
- Crowe, J.H.; Hoekstra, F.A.; Crowe, L.M. Anhydrobiosis. Annu. Rev. Physiol. 1992, 54, 579–599. [Google Scholar] [CrossRef]
- Winters, Y.D.; Lowenstein, T.K.; Timofeeff, M.N. Identification of Carotenoids in Ancient Salt from Death Valley, Saline Valley, and Searles Lake, California, Using Laser Raman Spectroscopy. Astrobiology 2013, 13, 1065–1080. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Brocks, J.J. Identification of carotane breakdown products in the 1.64 billion year old Barney Creek Formation, McArthur Basin, northern Australia. Org. Geochem. 2011, 42, 425–430. [Google Scholar] [CrossRef]
- Dartnell, L.R.; Hunter, S.J.; Lovell, K.V.; Coates, A.J.; Ward, J.M. Low-Temperature Ionizing Radiation Resistance of Deinococcus radiodurans and Antarctic Dry Valley Bacteria. Astrobiology 2010, 10, 717–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hays, L.E.; Graham, H.V.; Des Marais, D.J.; Hausrath, E.M.; Horgan, B.; McCollom, T.M.; Parenteau, M.N.; Potter-McIntyre, S.L.; Williams, A.J.; Lynch, K.L. Biosignature Preservation and Detection in Mars Analog Environments. Astrobiology 2017, 17, 363–400. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose (Nominal) | Dose (Applied) | Exposure Time |
|---|---|---|
| 0 | 0 | 0 |
| 6 | 6.21 | 1 h |
| 12 | 11.59 | 2 h |
| 18 | 17.51 | 3 h |
| 24 | 23.92 | 4 h |
| 47 | 46.88 | 7 h 50′ |
| 72 | 72.16 | 12 h |
| 113 | 113.25 | 18 h 50′ |
| Nominal Dose | Chroococcidiopsis sp. CCMEE 029 Dried | Chroococcidiopsis sp. CCMEE 029 Liquid |
|---|---|---|
| 0 | 100.0% | 100.0% |
| 6 | 100.0% | 100.0% |
| 12 | 100.0% | 96.4% |
| 18 | 100.0% | 100.0% |
| 24 | 100.0% | 99.5% |
| 47 | 100.0% | 99.8% |
| 72 | 100.0% | 73.7% |
| 113 | 100.0% | 100.0% |
| Gene | KEGG Enzyme | Closest BlastN Hit (GenBank ID) | Identity % | Genbank Accession n° |
|---|---|---|---|---|
| crtQ | K00514 | Nostoc sp. cyanobiont (CP026692.1) | 79.69% | MT438481 |
| crtB | K02291 | Scytonema sp. HK-05 (AP018194.1) | 76.91% | MT438479 |
| crtO(1) | K02292 | Scytonema sp. HK-05 (AP018194.1) | 82.29% | MT438474 |
| crtO(2) | K02292 | Scytonema sp. HK-05 (AP018194.1) | 80.32% | MT438476 |
| crtP | K02293 | Tolypothrix sp. PCC 7910 (CP050440.1) | 79.44% | MT438478 |
| crtR | K02294 | Scytonema sp. HK-05 (AP018194.1) | 78.79% | MT438480 |
| crtH | K09835 | Crinalium epipsammum PCC 9333 (CP003620.1) | 77.51 | MT438471 |
| crtW | K09836 | Nostoc sp. ATCC 53,789 (CP046703.1) | 73.80% | MT438477 |
| crtU | K09879 | Calothrix sp. NIES-2100 (AP018178.1) | 72.65% | MT438469 |
| crtI(1) | K10027 | Gloeobacter kilaueensis JS1 (CP003587.1) | 72.02% | MT438468 |
| crtI(2) | K10027 | Nostoc linckia NIES-25 (AP018223.1) | 71.80% | MT438473 |
| cruA | K14605 | Calothrix sp. PCC 7507 (CP003943.1) | 75.25% | MT438470 |
| cruP | K14606 | Nostoc sp. NIES-4103 (AP018289.1) | 77.59% | MT438467 |
| crtX | K14596 | Scytonema sp. NIES-4073 (AP018268.1) | 78.09% | MT438475 |
| crtY | K22502 | Calothrix sp. NIES-4105 (AP018290.1) | 72.97% | MT438472 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baqué, M.; Napoli, A.; Fagliarone, C.; Moeller, R.; de Vera, J.-P.; Billi, D. Carotenoid Raman Signatures Are Better Preserved in Dried Cells of the Desert Cyanobacterium Chroococcidiopsis than in Hydrated Counterparts after High-Dose Gamma Irradiation. Life 2020, 10, 83. https://doi.org/10.3390/life10060083
Baqué M, Napoli A, Fagliarone C, Moeller R, de Vera J-P, Billi D. Carotenoid Raman Signatures Are Better Preserved in Dried Cells of the Desert Cyanobacterium Chroococcidiopsis than in Hydrated Counterparts after High-Dose Gamma Irradiation. Life. 2020; 10(6):83. https://doi.org/10.3390/life10060083
Chicago/Turabian StyleBaqué, Mickael, Alessandro Napoli, Claudia Fagliarone, Ralf Moeller, Jean-Pierre de Vera, and Daniela Billi. 2020. "Carotenoid Raman Signatures Are Better Preserved in Dried Cells of the Desert Cyanobacterium Chroococcidiopsis than in Hydrated Counterparts after High-Dose Gamma Irradiation" Life 10, no. 6: 83. https://doi.org/10.3390/life10060083